元谋干热河谷滇榄仁群落林下物种多样性与幼苗更新特征
2011-05-30王小庆刘方炎陈友根
王小庆,刘方炎,2,李 昆,陈友根
(1.中国林业科学研究院 资源昆虫研究所,国家林业局 云南元谋荒漠生态系统定位观测站,云南 昆明 650224;2.北京林业大学 生物科学与技术学院,北京 100083)
中国云南金沙江干热河谷是长江上游重要的水源涵养和水土保持区,由于恶劣的气候环境以及长期严重的人为干扰作用导致了该区域植被严重退化和生物多样性逐渐丧失。该区域现存森林植被已极为稀少,且破碎化严重,大多已逐渐退化成稀树灌草丛植被类型[1]。元谋坝位于金沙江流域中下游,是金沙江干热河谷最为典型的生态脆弱区。人为干扰异常频繁,植被极度退化,土壤裸露,水土流失严重,是中国甚至世界上生态恢复极为困难的区域。一直以来,植被恢复是该区退化生态系统恢复和水土流失控制的重要手段。滇榄仁Terminalia franchetii系使君子科Combretaceae榄仁树属Terminalia落叶小乔木,高4~10 m,耐干旱瘠薄,喜光,为金沙江干热河谷一级标志种植物[1]。滇榄仁群落是元谋干热河谷区仅有的几种重要天然植被之一,对其群落物种多样性及幼苗更新特征的研究有助于认识该区现有天然植被的组成、发展及变化,有助于加强对滇榄仁植物群落的保护和合理利用,也为人工促进干热河谷退化植被的恢复提供一定的理论依据。本研究旨在了解元谋干热河谷区现有滇榄仁天然群落的物种组成及其多样性特征、滇榄仁群落下幼苗更新及其动态变化,以及滇榄仁群落结构对林下幼苗更新的影响。
1 研究区概况
研究区位于云南省楚雄彝族自治州元谋县(25°25′~ 26°07′N 101°35′~ 102°15′E)境内, 属于低纬度高原季风气候,年平均气温为21.9℃,最冷月(12月)平均气温14.5℃,极端低温-0.8℃,最热月(5月)平均气温27.1℃,极端高温42.0℃,≥10℃的年积温8003.0℃,年平均地温26.2℃,最热月(4月)平均地温61.1℃,极端最高地温74.3℃,最冷月(1月)平均地温16.4℃,极端最低地温1.4℃;年均降水量为634.0mm,其中雨季(5-10月)降水量583.8mm,占全年总降水量的92%左右,年均蒸发量为3911.2mm,约为年均降水量的6.4倍。滇榄仁群落位于河谷两侧山坡上,由于受人为干扰程度不同,现有群落类型也存在较大差异。全面勘察滇榄仁分布情况,依据乔木层郁闭度、干扰程度等选取3个典型群落类型,分别为群落Ⅰ,群落Ⅱ和群落Ⅲ,均为滇榄仁纯林,林下灌木层主要有车桑子Dodonaea viscose,疏序黄荆Vitex negundo,余甘子Phyllanthus emblica等。各群落参数见表1。
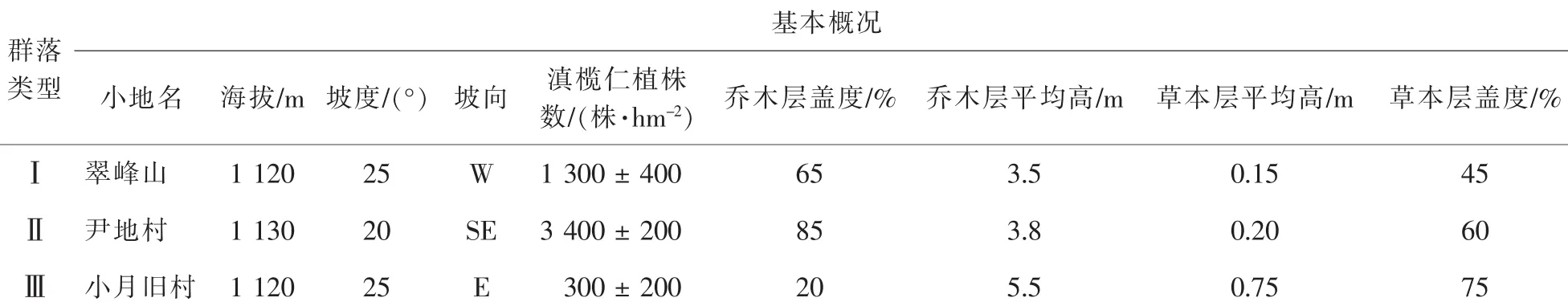
表1 不同群落类型基本概况Table1 Basic situation in three communities
2 研究方法
2.1 群落调查
在不同滇榄仁群落类型中依据典型取样原则各设置7个10 m×10 m大样方,然后用相邻格子法把每个大样方细分为4个5 m×5 m的小样方,在小样方中心设置1个1 m×1 m样方调查草本层植物。在大样方内对树高1.5 m以上植株进行每木检尺,记录物种名称、胸径、树高、枝下高、冠幅和生长状况等;在1 m×1 m的小样方中,记录1.5 m以下草本层植物种名,株(丛)数、平均高度及盖度等。
2.2 幼苗更新调查
滇榄仁林下幼苗更新调查采用样线法。具体方法为:沿山体等高线布置50 m样线,间隔1 m设置1 m×1 m小样方,分别计数样方内滇榄仁当年萌发幼苗数、当年萌发幼苗枯萎数、2年生幼苗数、3年生及以上幼苗数。每条样线计25个小样方,每一群落类型分别设置3条样线,计75个小样方。幼苗调查时,由于林下当年萌发幼苗和2年生幼苗通过叶片生长状况及茎干木质化程度较容易分清,但2年生以上幼苗不易区分,因此,将1.5 m以下2年生以上幼苗均作为3年生及其以上幼苗计数。间隔2个月进行1次调查,即2009年6月、8月、10月、12月及2010年2月、4月各调查1次林下幼苗更新。
2.3 数据分析
2.3.1 分析方法 具体如下:重要值VI=(相对多度AR+相对频度FR+相对盖度CR)/3;物种丰富度指数R=S;Shannon-Wiener多样性指数H′=-∑Pilog Pi(简称Shannon指数);Simpson优势度指数D=1-∑ Pi2;Pielou均匀度指数J=H′/log S。其中:Pi为种i在样方内的重要值,S为样方内物种总数[2-5]。2.3.2 统计分析 利用SPSS 16.0对数据进行处理分析,首先用方差齐性检验(homogeneity of variance test)进行方差齐性检验,再进行方差分析,并采用最小显著差法(LSD)进行多重比较。柱形图、折线图均使用Excel 2003软件绘制。
3 结果与分析
3.1 群落物种组成及多样性特征
3.1.1 群落物种组成特征 滇榄仁群落林下物种组成较为简单。发现种子植物32种,隶属于16科28属,其中禾本科Poaceae植物(9属9种)居多,占28.1%;蝶形花科Papilionaceae次之(4属5种),占15.6%;单科单属单种植物较多,占34.4%。从不同植物在群落中的重要程度来看(表2),扭黄茅Heteropogon contortus,矛叶荩草Arthraxon prionodes,龙须草Eulaliopsis binata及草本叶下珠Phyllanthus urinaria等耐旱草本植物在不同类型群落均存在,但优势程度不同。
3.1.2 群落物种多样性 由图1可以看出,不同滇榄仁群落林下植被中物种丰富度均相对较低,单位面积(1 m×1 m)物种数约4~7种,其中,群落Ⅰ和群落Ⅱ相对较低,且两者不存在显著差异,而群落Ⅲ的丰富度指数显著高于前两者。采用Shannon-Wiener多样性指数、Simpson优势度指数和Pielou均匀度指数进一步对其物种多样性进行分析发现,滇榄仁群落中,Shannon-Wiener多样性指数为1.35~2.82,Simpson优势度指数为0.63~0.81,Pielou均匀度指数为0.68~0.85,不同滇榄仁群落类型间存在显著差异。其中,群落Ⅲ的Shannon-Wiener多样性指数和Pielou均匀度指数均为最大,其次是群落Ⅱ,而群落Ⅰ最小。在Simpson优势度指数上,3个群落依次为群落Ⅰ>群落Ⅱ>群落Ⅲ。Shannon-Wiener多样性指数反映群落结构的复杂程度,其值越大,群落越复杂;Simpson优势度指数反映群落物种的地位和作用,其值越大,优势种越不明显;而均匀度为群落中不同物种的多度(生物量、盖度或其他指标)分布的均匀程度。由此可知:群落Ⅰ中林下植物种类最少,结构最为简单,各种植物在群落中的地位较为平衡,不存在明显的优势种(Simpson优势度指数最大),且其分布的均匀程度最低;群落Ⅱ林下物种多样性程度居中,林下有少量植物占有一定的优势,如扭黄茅等;群落Ⅲ中植物种类最多,林下草本植物得到了较大发展,分布最为均匀,少量植物在林下已占居较大优势(Simpson优势度指数最小)。
3.2 林下幼苗更新特征
从图2中可以看出,不同年龄滇榄仁幼苗数量在各群落中的变化趋势基本一致。其中,当年萌发幼苗数量在6月和8月时最多,10月开始出现锐减,而至翌年4月时数量最少;当年萌发幼苗枯萎数量与萌发数变化基本一致,即6月和8月最多,其他时间段均较少;而2年生及以上幼苗数量相对稳定,不同时间段差异不大。方差分析结果也表明,当年萌发幼苗及枯萎幼苗数在1 a内变化差异极显著(P<0.01),2年生及以上幼苗在1年中变化差异不显著(P>0.05)。同时,各群落中,6月和8月时,当年萌发幼苗均远高于2年生及以上幼苗数量,但10月、12月以及翌年2月和4月时却少于2年生及以上幼苗数量,表明在10月和12月以及翌年2月和4月时,当年萌发幼苗出现了大量死亡现象。另一方面,3种群落类型之间比较可知,6月和8月以及10月时,群落Ⅱ林下当年萌发幼苗及枯萎幼苗数均远高于群落Ⅰ和群落Ⅲ,其中,群落Ⅲ最少;10月和12月以及翌年2月和4月时,3种群落中,当年萌发幼苗数量相差不大。而1 a中,2年生及以上幼苗数量变化不大。由此可知:滇榄仁当年萌发幼苗无论是在应对不同季节的气候变化,还是在应对不同群落环境时均表现出了较大的脆弱性,容易出现枯萎和死亡,而2年生以上幼苗在不同季节时以及不同群落环境中均具有较好的稳定性。
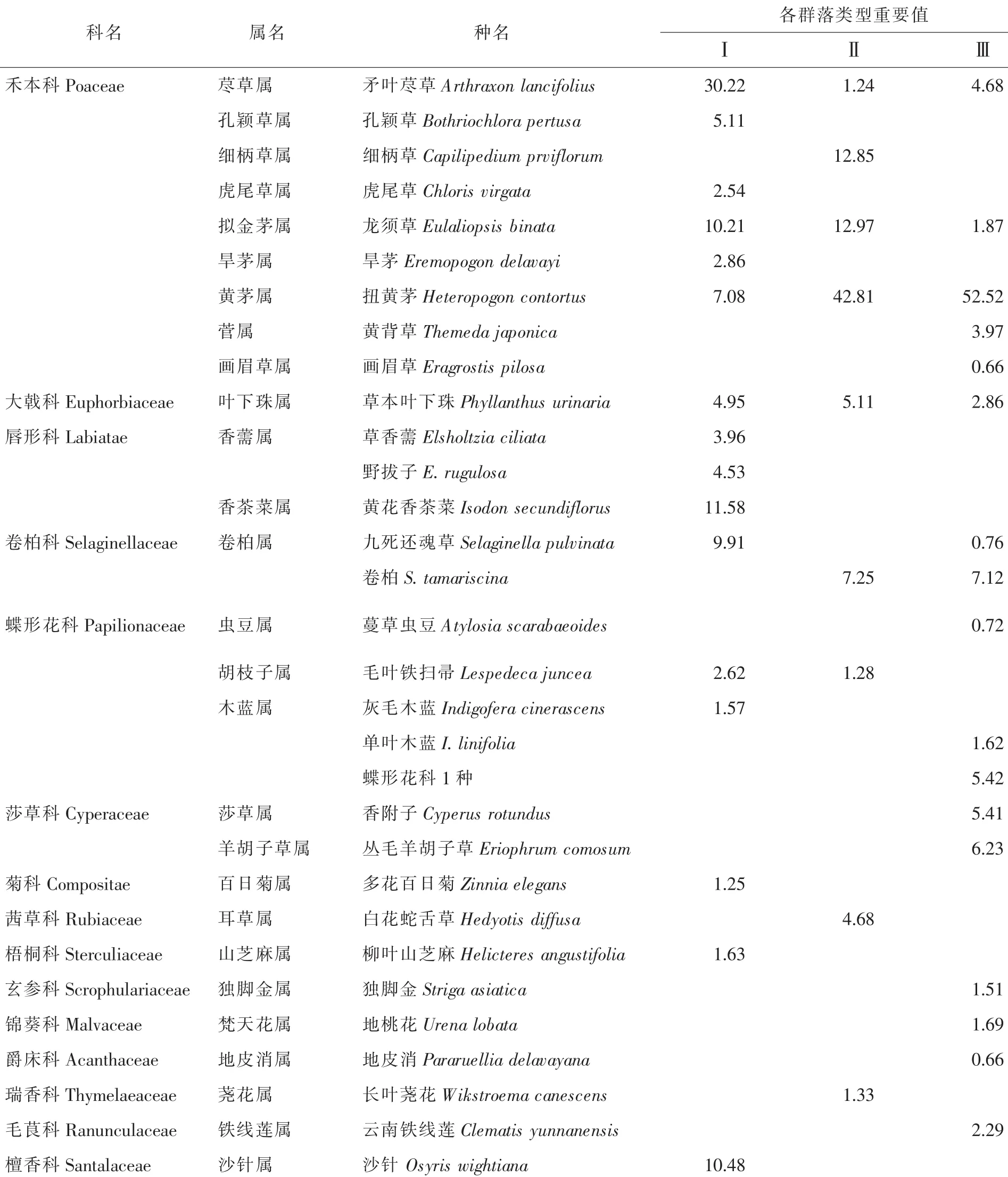
表2 滇榄仁群落林下物种组成及重要值分布Table2 Species composition and important values of Terminalia franchetii communities
4 结论与讨论
4.1 滇榄仁群落学特征
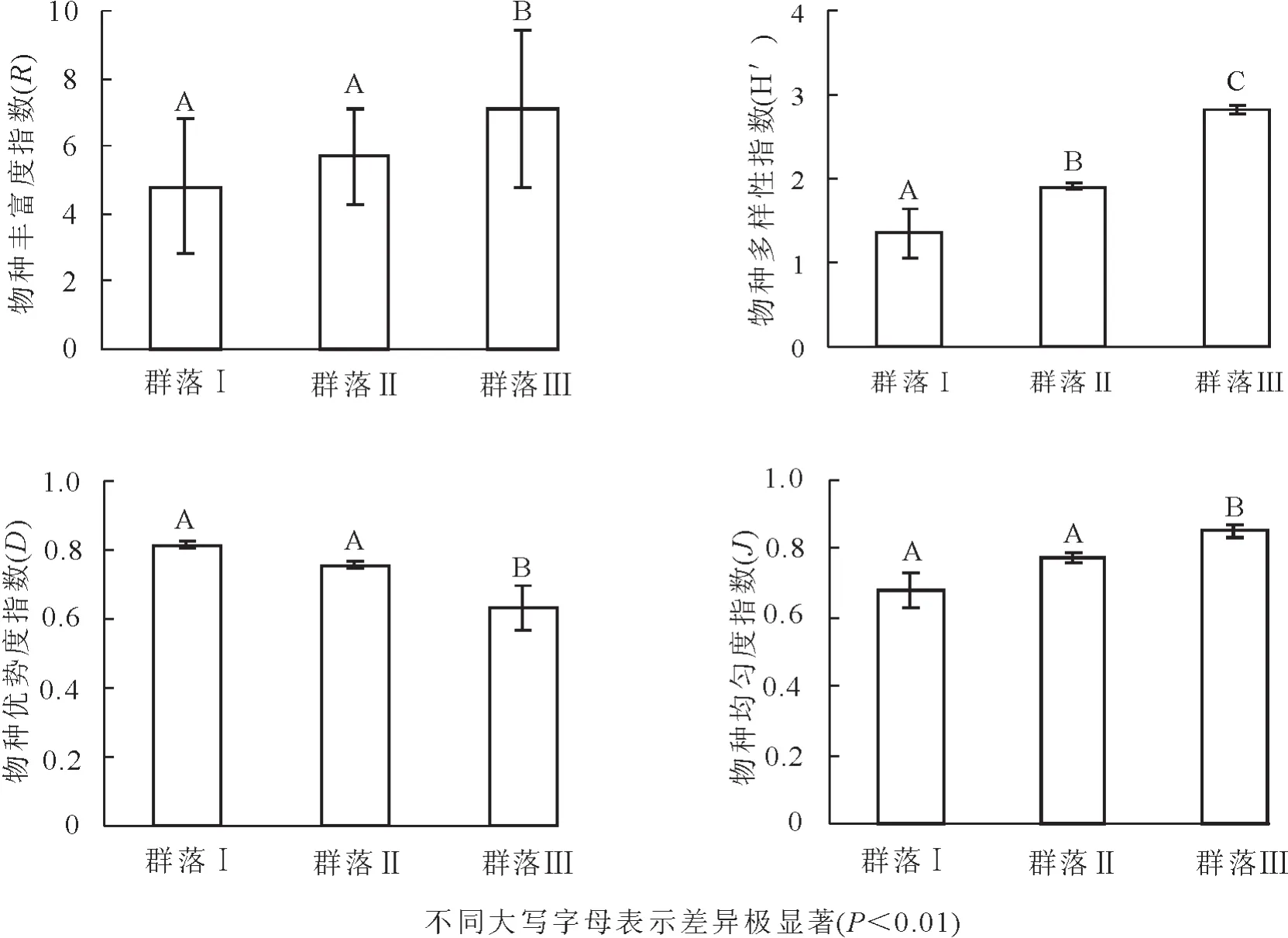
图1 不同滇榄仁群落林下物种多样性比较Figure1 Species diversity of three Terminalia franchetii communities
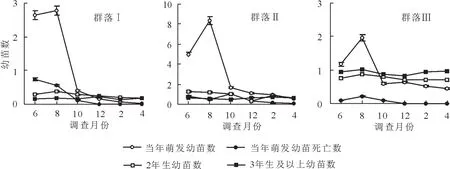
图2 滇榄仁群落林下幼苗数量动态变化Figure2 Soil moisture content and seedling regeneration in three Terminalia franchetii communities
滇榄仁是元谋干热河谷一种重要的乡土植物,由其组成的群落类型也是该区域仅有的几种天然植被。由于生存环境极为恶劣,且受人为影响程度不同,其林地破碎化程度存在较大差异。本研究所调查的3种群落类型中,人为活动的痕迹均较为明显,主要方式为砍伐、刈割、放牧、收获林产品和林下凋落物等,林下植被受影响程度较大。通过对不同类型滇榄仁群落调查发现,滇榄仁群落林下物种组成较为简单,以禾本科和蝶形花科植物居多,其中,有50%以上的样方统计到扭黄茅、矛叶荩草等植物,由此可知,虽然不同群落类型中林地生境、地貌存在着较大差异,但这2种植物始终是滇榄仁群落中分布较为广泛的植物种类。九死还魂草Selaginella pulvinata,卷柏S.tamariscina,龙须草,草本叶下珠等旱生草本植物在各群落中均存在,且在群落中的重要值较大,为各群落林下植被的优势种。
干热河谷林下植被中禾草等丛生植物较多,记录植物个体数目存在较大困难,因此以重要值作为参数来测度其多样性程度具有更好的准确性[6-7]。从滇榄仁群落的多样性程度可以看出,不同类型滇榄仁群落林下物种多样性程度均较低,且存在显著差异。其中,Shannon-Wiener多样性指数的变化最大,为1.35~2.82,其指数最小值已接近北方沙漠中的荒漠群落植物多样性[8-9],表明元谋干热河谷区滇榄仁群落环境条件极为恶劣,林下物种多样性较低。仅从植物多样性方面来看,林下植被的生存、繁衍已受到较大的环境压力,已接近沙漠中的荒漠群落类型。由于受干扰程度不同,群落又表现出了较大的差异。
4.2 林下幼苗更新特征
林下幼苗更新是群落动态的重要组成部分,是群落维持稳定的决定因素,且直接影响种群繁衍[10]。元谋干热河谷区由于气候、地质环境等条件较为恶劣,尤其是降水量的年内分配极为不均,常导致长达7个月的旱季(11月至翌年5月),极易出现严重干旱,直接影响天然植被林下幼苗的成功更新。滇榄仁作为中国西南干热河谷地区特有植物种,在长期的环境压力和各种强烈干扰下仍能维持种群的长期存在,表明其更新机制具有较强的干热河谷环境适应能力。
研究结果表明:元谋干热河谷区不同滇榄仁群落林下2年生及以上幼苗数量虽较少,但并没有随季节的变化而产生较大变动,表明2年生及以上幼苗已具有较强的抵抗恶劣环境压力的适应能力,能较好地在其林下定居。而不同群落中当年萌发幼苗产生时间较为集中而短暂,且数量较大。当年萌发幼苗由于建成初期还未形成较大的根系系统,个体生长较为脆弱,对环境变化极为敏感[11],因此,不同季节时其数量变动极大。更新幼苗输入主要发生在雨季早期(6月)和中期(8月),随着雨季的结束,林下种子萌发逐渐停止,当年萌发幼苗数也逐渐减少。而旱季时,滇榄仁林下种子库不能萌发,且随着干旱程度的加剧,雨季萌发的更新幼苗也因干旱胁迫而越来越少,尤其是干旱最严重的3月和4月,林下基本不存在当年萌发幼苗,仅荫蔽的石缝、草丛、母树基部等环境相对较好的地方有极少幼苗能够勉强存活,待到雨季的再次来临。群落环境,包括林下植被、土壤、地形等对更新幼苗数量具有较大影响[12-14]。通常,成功的天然更新取决于群落所处的大气候条件以及群落内外的小气候或环境因子。同时,天然更新的必要条件是具有充足的、较强生命力的种子以及适宜的林地条件[15]。群落Ⅲ中由于滇榄仁母树极少(表1),林地存在较大的种源限制可能是导致其雨季时当年萌发幼苗数量为3种群落中最少的直接原因,但其旱季时的幼苗保存率却为3种群落中最高,这可能是因为该群落林下植被发育较好,形成较好的林地环境,有助于幼苗躲避极度干旱等环境因子而更多地存活下来。而群落Ⅰ和Ⅱ中成年植株较多,种源相对充足,雨季时也有较大量的萌发幼苗产生,但由于群落内多样性程度低下,植被稀疏,土壤裸露,环境退化,无法保护幼苗抵御漫长旱季的干旱胁迫导致大量幼苗死亡从而更新不佳。
在生态脆弱区,群落结构、类型以及多样性程度往往反映的是林地受干扰程度[16]。对元谋干热河谷不同类型滇榄仁群落的调查发现,由于当地放牧活动异常频繁,因此距离村庄较近的群落Ⅰ林下物种多样性最差,其次是群落Ⅱ,而群落Ⅲ距离村庄较远,干扰强度相对较小,其林下植物保存较好,物种多样性程度亦相对较高。因此,只要加强森林保护的宣传力度,提高当地群众的保护意识,加大封山育林的力度,从而减少人为干扰,使之形成良好的林下环境,滇榄仁幼苗便容易成功定居,从而通过天然更新达到自然恢复天然植被的目的,加速干热河谷植被恢复和生态治理步伐。
[1]金振洲,杨永平,陶国达.华西南干热河谷种子植物区系的特征、性质和起源[J].云南植物研究,1995,17(2): 129-143.JIN Zhenzhou, YANG Yongping, TAO Guoda.The floristic characteristics, nature and origin of seed plants in the dry-hot river valley of southwest China [J].Acta Bot Yunnan, 1995, 17 (2): 129-143.
[2]马克平,刘玉明.生物群落多样性的测度方法(Ⅰ)α多样性的测度方法(下)[J].生物多样性,1994,2(4):231-239.MA Keping, LIU Yuming.The method on the measurement of species diversity in biotic community(Ⅰ)the measurement of α diversity [J].Biodiversity Sci, 1994, 2 (4): 231-239.
[3]李振基,陈小麟,郑海雷,等.生态学[M].北京:科学出版社,2000:204-207.
[4]朱彪,陈安平,刘增力,等.广西猫儿山植物群落物种组成、群落结构及树种多样性的垂直分布格局[J].生物多样性, 2004, 12 (1): 44-52.ZHU Biao, CHEN Anping, LIU Zengli, et al.Changes in floristic composition, community structure, and tree species diversity of plant communities along altitudinal gradients on Mt.Mao’er, Guangxi, China [J].Biodiversity Sci,2004, 12 (1): 44-52.
[5]李林,周小勇,黄忠良,等.鼎湖山植物群落α多样性与环境的关系[J].生态学报,2006,26(7):2301-2307.LI Lin, ZHOU Xiaoyong, HUANG Zhongliang, et al.Study on the relationship between α diversity of plant community and environment on Dinghushan [J].Acta Ecol Sin, 2006, 26 (7): 2301-2307.
[6]马克平,黄建辉,于顺利,等.北京东灵山地区植物群落多样性的研究(Ⅱ)丰富度、均匀度和物种多样性指数[J].生态学报, 1995, 15 (3): 268-277.MA Keping, HUANG Jianhui, YU Shunli, et al.Plant community diversity in Dongling Mountain, Beijing, China(Ⅱ)Species richness, evenness and species diversities [J].Acta Ecol Sin, 1995, 15 (3): 268-277.
[7]李贵祥,施海静,孟广涛,等.云南松原始林群落结构特征及物种多样性分析[J].浙江林学院学报,2007,24(4): 396-400.LI Guixiang, SHI Haijing, MENG Guangtao, et al.Community structural properties and species diversity in primary Pinus yunnanensis forest[J].J Zhejiang For Coll, 2007, 24 (4): 396-400.
[8]张林静,岳明,张远东,等.新疆阜康绿洲荒漠过渡带植物群落物种多样性特征[J].地理科学,2003,23(3):329-334.ZHANG Linjing, YUE Ming, ZHANG Yuandong, et al.Characteristics of plant community species diversity of oasis desert ecotone in Fukang, Xinjiang [J].Sci Geogr Sin, 2003, 23 (3): 329-334.
[9]孔丽娟,沈吉庆.腾格里沙漠东南边缘植物群落物种多样性分析[J].宁夏农学院学报,2003,24(4):25-28,41.KONG Lijuan,SHEN Jiqing.Analysis on species diversity of plant communities in the southeastern Tenggeli Desert[J].J Ningxia Agric Coll, 2003, 24 (4): 25-28, 41.
[10]潘春柳,赖家业,黎向东,等.单性木兰种子雨与天然更新的初步调查[J].生态学杂志,2008,27(12):2235-2239.PAN Chunliu, LAI Jiaye, LI Xiangdong, et al.Seed rain and natural regeneration of Kmeria septentrionalis in Mulun of Guangxi[J].Chin J Ecol, 2008, 27 (12): 2235-2239.
[11]陈迪马,潘存得,刘翠玲,等.影响天山云杉天然更新与幼苗存活的微生境变量分析[J].新疆农业大学学报,2005, 28 (3): 35-39.CHEN Dima, PAN Cunde, LIU Cuiling, et al.Analysis on micro-habitat variables affecting natural regeneration and survival of seedlings in Picea schrenkiana stand [J].J Xinjiang Agric Univ, 2005, 28 (3): 35-39.
[12]张昆林,钟蓉军.地形条件对高山松天然更新和生长的影响[J].林业科技,2000,25(1):22-23.ZHANG Kunlin,ZHONG Rongjun.The impact of landform condition on natural regeneration and growth of Pinus densata [J].For Sci Technol, 2000, 25 (1): 22-23.
[13]曾德慧,尤文忠,范志平,等.樟子松人工固沙林天然更新障碍因子分析[J].应用生态学报,2002,13(3):257-261.ZENG Dehui, YOU Wenzhong, FAN Zhiping, et al.Analysis of natural regeneration barriers of Pinus sylvestris var.mongolica plantation on sandy land [J].Chin J Appl Ecol, 2002, 13 (3): 257-261.
[14]朱教君,刘足根,王贺新.辽东山区长白落叶松人工林天然更新障碍分析[J].应用生态学报,2008,19(4):695-703.ZHU Jiaojun, LIU Zugen, WANG Hexin.Obstacles for natural regeneration of Larix olgensis plantations in montane regions of eastern Liaoning Province, China [J].Chin J Appl Ecol, 2008, 19 (4): 695-703.
[15]姜凤岐,朱教君,周新华,等.林带的防护成熟与更新[J].应用生态学报,1994,5(4):337-341.JIANG Fengqi, ZHU Jiaojun, ZHOU Xinhua, et al.Protective maturity (PM) and regeneration of shelterbelts [J].Chin J Appl Ecol, 1994, 5 (4): 337-341.
[16]王永健,陶建平,张炜银,等.茂县土地岭植被恢复过程中物种多样性动态特征[J].生态学报,2006,26(4):1028-1036.WANG Yongjian, TAO Jianping, ZHANG Weiyin, et al.Dynamics of species diversity in vegetation restoration on Tudiling of Mao County Southwest China [J].Acta Ecol Sin, 2006, 26 (4): 1028-1036.
