Toll样受体引发宿主抵抗弓形虫
2011-05-15潘磊陈会周玉刚迟海
潘磊,陈会,周玉刚,迟海
(1.安徽省蚌埠市畜牧兽医技术推广站,安徽蚌埠 233000;2.安徽科技学院,安徽凤阳233100;3.安徽省蚌埠市淮上区农业综合服务站,安徽蚌埠233000)
弓形虫是一种重要的胞内寄生原虫,它可以在广泛的动物宿主和大部人群中建立广泛稳定的宿主-寄生关系。依赖MyD88的Toll样受体信号传导是激发宿主对弓形虫这种机会性致病寄生虫防御能力的一种重要途径,其也可能介导免疫功能障碍过程中的致病作用。其他MyD88的独立的信号途径也参与了宿主-寄生虫的相互作用。这些反应可由寄生虫自身引发,但与肠道菌群的相互作用增加了额外的肠道感染复杂性。
一、前言
宿主对感染的防御依赖于有效的病原检测机制和区分感染和非感染性物质的能力。对于全球分布的原虫——弓形虫来说,感染的识别诱发一个快速和强大的、对弓形虫在宿主内生存和长期持续存在所必须的Th1细胞极化免疫反应。在以中枢神经系统和骨骼组织肌肉中出现包囊为特征的慢性感染的情况下,产生干扰素-γ的CD4 +和CD8 +T淋巴细胞必须保持静态感染。阐明这种认识的最显著的证据来自于艾滋病,在该病的发展过程中,由于T细胞数量的减少,包囊可能能重新被激活,将产生灾难性后果。
过去20年间的研究,已经证实了 IL -12作为一个早期细胞因子的重要性,其刺激免疫系统发展极化Th1反应以便于控制弓形虫。在IL-12缺乏的情况下,小鼠由于过量的寄生虫感染和大量组织的破坏而快速发病。IL-12的产生允许宿主呈慢性感染而存活。在弓形虫感染期间,树突状细胞产生的IL-12显著增加,其次巨噬细胞和中性粒细胞也产生这种细胞因子。这些类型的细胞在体内感染过程中是入侵靶向细胞,但是否感染细胞本身被触发而产生IL-12或是否细胞因子是由未感染细胞对寄生虫产物做出反应而产生的,目前尚不清楚。不仅如此,显而易见的是,IL-12的产生必须被严密的控制以防止产生炎性病理反应。弓形虫感染中,IL-10是一个关键的表达量下调的细胞因子。这样,IL-10基因敲除小鼠的感染的动态过程非常类似于IL-12和IFN-γ基因缺失的动物。然而,IL-10缺失动物的死亡是与较低数量的寄生虫感染和IL-12、TNF-α和IFN-γ显著高表达相关的。然而,过去认为IL-10主要由细胞巨噬细胞系产生,最近的研究证实弓形虫感染过程中,Th1细胞本身是IL-10的重要来源。
在弓形虫和其他的微生物感染过程中,这些复杂的但必要的反应是如何被激活的,一直是研究者长期以来想弄清楚的兴趣所在。上世纪八十年代,Janeway提出了先天免疫的细胞用模式识别受体(PRRs)对病原体相关(PAMPs)分子模式做出反应的观念。10年后,Janeway和Medzhitov发现一个主要的PRR家族蛋白,即Toll样受体(TLR)。尽管TLRs在结构上类似,但它们在识别多样性的微生物分子,包括蛋白、脂类,DNA和RNA分子上是显著不同的。微生物配体触发的Toll样受体在激发对包括弓形虫在内的许多种微生物的免疫起着重要的作用。
二、依赖MyD88的Toll样受体信号传导在防御弓形虫上的重要性
关于Toll样受体及其配体的早期研究主要聚焦于细菌和病毒分子,如脂多糖和核酸的识别,但很快就发现TLRs在原虫的识别中也起着关键作用,可以激发宿主对这类病原体的免疫作用。MyD88基因敲除小鼠的研究首先证实了TLR在抵抗弓形虫中的重要性。MyD88分子是一种常见的TLR信号适配器,这是因为几乎所有的TLRs使用此分子在宿主细胞传递信号。因此,MyD88基因的灭活导致了几乎所有的TLR信号的损失。TLR3和TLR4是一个例外,它使用另一种接头分子称为包含适配器-诱导干扰素(TRIF)的Toll-IL-1受体域。在腹腔注射的小鼠模型中,用正常低毒力的ME49寄生虫虫株感染的情况下,MyD88的缺失导致小鼠的快速死亡。死亡率与高寄生虫数量和低水平的IL-12和干扰素-γ相关。此外,在MyD88的缺失的情况下,中性粒细胞被发现也缺失,其影响因素在其他使用这种小鼠品系的传染性疾病模型中已有报道。
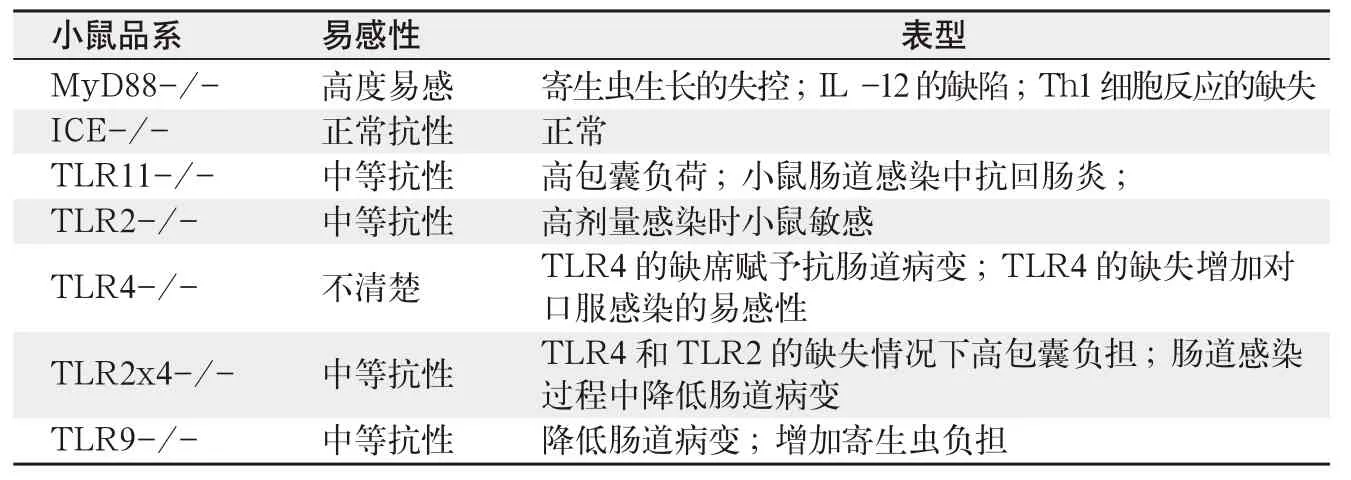
表1 TLR/MyD88基因敲除小鼠研究进展概况
最近发现,通过口服途径感染弓形虫的动物,在MyD88缺失情况下,对弓形虫的易感性增加。MyD88最常见的功能是与先天免疫细胞信号传导相关,但有证据表明,获得性免疫也使用这种常见的接头分子。因此,骨髓嵌合体小鼠的研究表明,在慢性感染过程中,MyD88的T细胞表达对于防止出现弓形虫脑炎是必须的。虽然MyD88基因敲除小鼠对弓形虫的易感性表明TLR不仅识别寄生虫,该接头分子通过受体如IL-1和IL-18参与了信号传导。然而,缺乏能产生有生物活性的IL-1和 IL-18的 IL-1β转化酶(ICE)的小鼠,表现出正常的抗感染。这也是一重要的成果,因为它提供了有力的证据表明在抗弓形虫感染中,需要TLR信号本身的参与。
虽然来自MyD88-/-小鼠的的研究结果证实了它们的重要性,但目前还没有证据证实控制弓形虫的免疫反应的主要Toll样受体。随着进一步研究,已经鉴别出一种和小鼠TLR11受体作用的寄生虫配体。相对于MyD88基因敲除的小鼠,缺乏这种基因的小鼠在易感性上仅略有增加。类似地,也有报道说TLR2-/-小鼠也显示了对弓形虫的易感性增高,但这种影响只出现在用高接种量的情况下。在对TLR2-/-小鼠的研究中,动物感染后能够存活,但阻止了小鼠大脑中包囊数量的增加。
另一项研究表明,虽然不同的研究结果有区别,但TLR4的缺乏增加了肠道感染的易感性。鉴于这种真核细胞病原体的分子复杂性,认为弓形虫拥有多种Toll样受体配体是合理的,以至于任何单一配体的敲除对寄主抗性仅有轻微的影响(表1)。在宿主抗锥虫和结核分枝杆菌的研究中也得到类似的结论,其中TLR2和TLR9的结合提供了对感染的最佳免疫力。
三、寄生虫的TLR配体
目前,已确定了两种弓形虫TLR 配体的分子结构。速殖子表面均匀覆盖着糖基磷脂酰肌醇锚定蛋白。弓形虫磷脂锚定蛋白以及核心聚糖和脂基的纯化,以及随后的编码TLR和NFκB报告基因的质粒的细胞共转染刺激,揭示磷脂基激发TLR2和TLR4。在巨噬细胞,这会导致TNF-α的产生和主要组织相容性Ⅱ类分子的上调。有趣的是,分离自弓形虫的不饱和脂肪酸阻断了肿瘤坏死因子-α诱导磷脂基团特性。这可能与活虫体不诱导产生肿瘤坏死因子-α,而且在脂多糖-TLR4的激发过程中有效抑制其合成的观察相关。
迄今为止,磷脂基最接近作为真实的原生动物PAMP分子。这样,除了那些来自弓形虫的磷脂基,来自布氏锥虫、克氏锥虫、利什曼原虫、属原虫和疟原虫的磷脂基都具有TLR2或TLR4的激活特性。在一些虫种(原虫和锥虫属)中,这有助于提升保护性反应,但在其他的一些虫种中,这可能是致病的潜在原因(疟原虫)。为什么寄生虫磷脂激活TLR而哺乳动物磷脂不会触发自身免疫反应,可能与在这些寄生虫的表面GPI蛋白大量的表达和这些分子结构的特异性因虫种的不同而变化等相关。
众所周知,通过超声裂解并高速离心后获得的寄生虫可溶性裂解液(STAg)能够以一种依赖MyD88的方式刺激树突状细胞 IL-12的产生。裂解液中直接的生化分析导致了profilin的发现,其作为弓形虫的MyD88的依赖IL-12的诱导分子起作用。有趣的是,于此同时,也发现了在艾美耳球虫中具有IL-12诱导性能的profilin分子。然而,并非所有的顶复器原虫都具备诱导产生IL-12的能力,恶性疟原虫profilin仅具有很微弱的诱导产生IL-12的能力。弓形虫的TLR11被认为是profilin的宿主细胞受体。在小鼠体内表达,而人类体内不表达的TLR11 分子,起初被认为与肾盂肾炎细菌反应,但迄今为止弓形虫profilin是唯一被认定作为TLR11受体配体的这种分子。除了诱导宿主细胞的IL-12合成外,在弓形虫的入侵过程中也需要profilin,揭示了profilin在寄生虫-宿主相互作用中的双重作用。profilin不是一种分泌分子,而是在速殖子细胞质内表达。这表明,IL-12的引发可能是由那些对虫体降解物产生反应而不是对感染本事作用的细胞介导的。
TLR/MyD88途径被认为在控制T细胞免疫优势方面发挥了重要作用。这是因为专职抗原提呈细胞如树突状细胞可以同时处理和提呈抗原,同时以依赖于MyD88的方式上调T细胞共刺激分子的表达。在这种模型的支持下,表明在同一个树突状细胞群中,STAg的腹腔注射诱导的针对profilin的CD4+ T细胞反应是独立于TLR11表达和主要组织相容II类识别的。
四、TLR/MyD88在肠道中的作用,那一个是前驱者?
目前已经公认,在高剂量弓形虫感染时,某些近交系小鼠如C57BL/6株小鼠,出现严重的由炎性细胞因子介导的小肠病变。在低剂量感染情况下,IL-10预防这种病变的关键作用在IL-10-/-小鼠已得到证实。类似的肠道病变在其他感染弓形虫的动物物种也有报道,但这种情况在人身上能否发生,尚不清楚。C57BL/6小鼠上的病变在几方面类似于人的克罗恩病,这种病目前认为涉及肠道黏膜Th1细胞和Th17的错调反应。正如人类的炎性肠炎,其病变与肠道中微生物的变化相关。是否这些变化是病变的结果或诱导因素,目前是一个研究的热点。
在高剂量弓形虫感染过程中,革兰氏阴性细菌积累在肠道损伤的位。有趣的是,不仅TLR4和TLR9基因敲除小鼠对寄生虫引起的肠道黏膜损伤有抗性,而且这与前炎症细胞因子水平下降有关。此外,肠道菌群被广谱抗生素破坏的小鼠具有耐弓形虫引起的回肠损害的作用。以上结果表明以TLR为基础的细菌识别,而不是寄生虫感应本身,引起了肠道黏膜炎症病变。
最近的证据还表明,在低剂量弓形虫感染条件下,内源性肠道微生物对寄生虫有免疫佐剂的作用。这样,TLR11基因敲除小鼠,虽然无法对IL-12诱导TGPRF分子做出反应,但可以在肠道感染过程中发展了健全的Th1反应。然而,在TLR2x4-/ -和TLR9-/-小鼠,与保护相关的Th1细胞是缺失的。无菌小鼠弓形虫感染导致在肠道黏膜IL-12反应的缺陷,但如果用TLR4的配体脂多糖饲喂小鼠,这种反应能够恢复,进一步支持了这些结果。因此,在口服引起弓形虫感染时,虫体表达的TLR配体可能没有肠道粘膜表达的TLR配体作用大。根据这种观点,弓形虫感染会引起肠道的局部损伤,可能是寄生虫的入侵和细胞裂解特性的结果。在低剂量感染条件下,这允许将细菌易位,促进Th1细胞保护性分化,但高剂量感染条件下,将导致暴发性病变。然而,有趣的是,有报道说TLR11-/-小鼠可以抵抗寄生虫诱导的肠道病变,这个结果暗示了弓形虫TGPRF的作用,以及TLR4和TLR9对肠道炎症的作用(见表1)。
五、无MyD88参与的免疫识别和免疫保护作用
虽然Toll样受体和MyD88依赖的信号在宿主对弓形虫的反应是很重要的,但它并不是唯一途径。在骨髓来源的巨噬细胞的体外感染过程中,细胞产生独立MyD88信号的IL-12。有趣的是,这种作用具有寄生虫株特异性,因为诱导产生低水平的IL-12的生产的高毒力I型虫体病不依赖于MyD88,而诱导产生高水平的IL-12的低毒力II型虫体需部分依赖于MyD88。IL-12的产量控制与ROP16有关系,ROP16是一种通过介导影响信号转导和转录激活因子的棒状体激酶。
在肠道感染中,发现MyD88-/-的动物由于寄生虫复制的失控和传播而出现一致的死亡率。不过,虽然Th1细胞出现有一定的延误,但产生干扰素-γ的Th1细胞在感染后的一周作用达到正常水平。推测在这种情形下,虽然对于控制寄生虫来说需要MyD88,但MyD88对于诱导产生获得性免疫反应并不是必不可少的。有可能在无MyD88的情况下,免疫诱导是由细菌识别系统诸如并没参与TLR/MyD88信号途径的NOD2或非基于Toll样受体的寄生虫自身识别已经可以充分诱使延迟反应。
为了搞清楚这些问题,检验MyD88的基因敲除小鼠是否能产生保护性功能反应,我们用无毒尿嘧啶营养缺陷型菌株即cps1寄生虫- 1免疫小鼠,用RH株的毒力株攻毒。在这种情况下,寄生虫特异的Th1细胞的出现诱导了免疫反应,并证实了以前的预测,即动物对致命的攻毒具有一定的抵抗能力。因此,这似乎是一个弓形虫不依赖于MyD88的识别导致了具完全功能的适应性免疫。
不依赖于TLR/MyD88弓形虫识别的分子基础目前尚不清楚。但是,这种寄生虫引发的Gi-蛋白和GiPCR,导致一些包括CCL17和CCL22在内的趋化因子的释放。一个特别的趋化因子受体,CCR5,与IL-12的产生相关。如上所述,弓形虫在入侵过程中激活STAT3和STAT6。这个过程的精确机制目前尚不清楚,但这些转录因子的磷酸化并不需要MyD88(Butcher和Denkers,未出版)。
六、人TLR在抗弓形虫中的重要性
在小鼠体内广泛的研究已表明了TLR/MyD88通路在抗病毒、细菌,真菌和原生动物感染中的重要性。然而,这一途径在人类宿主防御的作用尚不明确。一个MyD88遗传性连续缺陷9代的患儿被发现对化脓性细菌易感,但出乎意料的对于包括弓形虫在内的其他感染具有抗性。这样,相对于这一信号通路在小鼠抗感染中的重大影响,TLR/MyD88可能在人类宿主防御可能发挥较小的作用。但是,另一种解释是,MyD88的缺失导致在感染早期致命的敏感性,以至于,患者根本无法被用于研究。对于弓形虫感染的特定案例,人类TLR11的缺乏表达被认为寄生虫profilin不是IL-12的主要诱导因素。不过,用人的TLR2,CHO细胞转染后会对速殖子磷脂分子衍生物产生反应,表明TLR2可能在人的感染过程中发挥作用。因此,TLR/MyD88在人类的抗弓形虫和其他感染的过程中的作用仍然是一个悬而未决的问题。
原作:Eric Y.Denkers康奈尔大学兽医学院微生物学与免疫学教研室
(略)
