干旱胁迫对乌头部分生理指标的影响
2011-05-12李洁周春娥梁志英苗青
李洁,周春娥,梁志英,苗青
(1.山西农业大学林学院,030801,山西太谷;2.河南师范大学生命科学学院,453007,河南新乡;3.晋中学院生命科学院,030600,山西晋中)
我国乌头(Aconitum)种质资源占世界总种数的57.14%。乌头属(Aconitum)植物主要作为药材栽培利用,也是宝贵的蓝色花卉基因资源和良好的观花园林植物[1];因此,对乌头的综合开发利用越来越受到人们的关注。但近年来,我国干旱缺水趋势日趋严重,尤其北方地区,给乌头的正常生长和推广应用带来了严峻考验。一直以来,有关乌头属植物的研究都集中在资源调查[2-3]、化学成分和药用价值方面[4-7],作为重要的药用植物,乌头属植物栽培技术[8]和组织培养[9]也有研究;但关于乌头属植物抗旱性的研究所见不多,对其在干旱条件下的综合开发利用有一定的盲目性。笔者选择山西省北乌头(Aconitum jeholense Nakai et Kitag.)、高乌头(Aconitum sinomontanum Nakai)、山西乌头(Aconitum smithii Ulbr.ex Hand.-Mazz.var.smithii)、牛扁(Aconitum barbatum Pers.var.puberulum Ledeb.)、华北乌头(Aconitum soongaricum Stapf var.angustius)5种乌头属植物为试材进行抗旱研究,并运用模糊隶属法,对其抗旱性做出综合评价,从而为选取抗逆性强的植物资源,并为乌头属植物在我国干旱、半干旱地区进一步综合开发利用提供方法和依据。
1 研究地概况
试验是在山西农业大学林学院苗圃进行,其位于 E 112°53'、N 37°30'的晋中盆地南部,海拔 870 m,属于温带大陆性季风气候,年降水量540 mm,无霜期为140~180 d,试验地土壤质地为壤土,基本理化性状为:pH值7.85,有机质质量分数9.22 g/kg,土壤氮、磷、钾的质量分数分别为1.1 g/kg、11.6 mg/kg、115.7 mg/kg。试验材料为乌头属高乌头、牛扁、山西乌头、北乌头、华北乌头5个种移栽苗,均于2009年6月采自于山西省关帝山,该地有乌头5种6变种,分布于郝家沟、崔家沟,赫赫崖等处。
2 材料与方法
2.1 试验设计和样本采集
采用完全随机区组设计[10]。植株采集后,分区组按20 cm×20 cm栽植,水分处理分3个水平:CK为正常供水,土壤相对含水量维持在70% ~75%;T1为中度干旱胁迫,土壤相对含水量维持在50%~55%之间;T2为严重干旱胁迫,土壤相对含水量维持在30% ~35%之间;设遮雨棚。
每个处理中每个种种植30株,设置3个重复。至2009年8月植株生长稳定后,开始水分胁迫试验,分别在处理后10、20、30 d取样,用于生理指标测定。试验材料为植株新鲜成熟叶片,测定时各乌头平均株高为:高乌头121 cm,牛扁52 cm,山西乌头92 cm,北乌头50 cm,华北乌头60 cm。在作柱形图的统计分析时,取各次所测结果的总平均值。
土样采集:每隔5 d,在每个样地分3处用土钻取试验材料根部10和20 cm处的土样混合,装入铝盒中,带回实验室称鲜质量,在105℃烘箱中烘干至恒质量,取平均值计算土壤相对含水量,依此监测并调整水分供应。
2.2 指标测定
细胞膜相对透性采用(DDS-11A)电导法测定[11],叶绿素(Chl)质量分数测定采用分光光度法,丙二醛(MDA)质量分数测定采用硫代巴比妥酸[12],光合速率(Pn)、气孔导度(Cs)、胞间 CO2摩尔分数采用便携式光合作用测定系统(Li—6400,Li-COR)进行同步测定[13],气孔限制值采用 Berry等[14-15]提出的计算公式

式中:Ls为气孔限制值;Ci为胞间 CO2摩尔分数,μmol/mol;C0为空气中 CO2摩尔分数,μmol/mol。
土壤相对含水量(%)采用称质量法测定。
2.3 数据处理及抗旱评价方法
对5种乌头的抗旱性评价采用模糊评价法,以平均抗旱隶属度为依据对各供试品种抗旱性强弱进行排序[10],其隶属函数度计算公式为
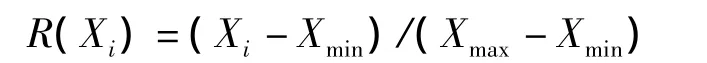
式中:Xi为指标i测定值,Xmin、Xmax分别为所有参试材料某一指标的最小值和最大值。隶属度越大,抗旱性越强。如果所测指标与抗旱性呈负相关,则用反隶属函数计算其抗旱隶属度
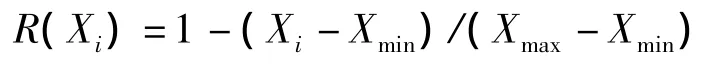
根据隶属函数法,用上述公式计算各指标在T1、T2处理下的隶属度,其中叶绿素质量分数和净光合速率的隶属度根据隶属函数公式计算,细胞膜透性和丙二醛的隶属度用反隶属函数公式计算,然后再分别将各乌头在T1和T2处理下的4个指标的隶属度相加,求其平均值,即为该乌头在对应干旱胁迫下的抗旱隶属度,隶属度越高,其抗旱性越强。
用Excel和SPSS10.0统计分析软件进行数据处理。
实验班数学成绩的平均分呈上升趋势,这反映了在辩证动态双主教学过程中,师生的角色定位逐渐趋向明晰,教与学的品质逐渐得到提升。教师由困惑与彷徨的“不会教”到目标明确、措施得力的“会教”,从不善于调控教学到收放自如,体现了“教”的品质的提升。学生从茫然不知所措的“不会学”,到善于沟通与交流的“会学”,真正成为学习的主人。他们学习方法的转变与学习过程的优化印证了“学”的品质的提升。辩证动态双主教学不仅实现了教与学的高品质,而且营造了良好的学习氛围,融洽了师生关系,促使师生地位趋向平等。就在教与学的契合点上,师生有了较大的”公约数“,思想的火花在动态中迸发,心灵的智慧在碰撞中萌生。
3 结果与分析
3.1 干旱胁迫对乌头细胞膜相对透性的影响
不同水分处理对乌头细胞膜相对透性的影响见图1。
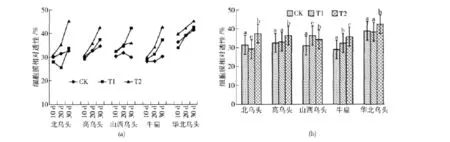
图1 不同水分处理对乌头细胞膜相对透性的影响Fig.1 Impacts of different water stress treatments on membrane permeability of different species
图1(a)为干旱处理后10、20、30 d所测乌头叶片的细胞膜透性。可以看出,北乌头的细胞膜透性在中度干旱胁迫后20 d时最低,表现出北乌头此时的短期修复作用,其余乌头的细胞膜透性在3种水分处理下基本表现出随时间推移而逐渐增大的趋势,除山西乌头外,其余乌头在对照条件下的细胞膜透性变化均较干旱胁迫下变化缓慢,山西乌头在对照条件下的细胞膜透性在30 d时比20 d时有所下降,表现出山西乌头后期的修复作用,同时也反映出此时的严重干旱胁迫在其可修复范围内。
图1(b)为不同水分处理下乌头细胞膜相对透性变化的柱形图:北乌头在严重干旱胁迫下细胞膜透性比对照增大了19.02%,增幅较大,差异达显著水平(P<0.05),其细胞膜透性在中度干旱胁迫下显著小于对照,反映出北乌头短期修复干旱损伤的能力较强,但不耐忍受长期和严重干旱胁迫;高乌头和华北乌头的细胞膜透性在中度干旱胁迫下没有显著变化,说明中度干旱胁迫对其细胞膜透性没有显著伤害,这2种乌头在严重干旱胁迫下细胞膜透性显著增大,分别比对照增大了12.31%和8.90%,反映出严重干旱胁迫下其细胞膜受害严重;山西乌头的细胞膜透性在中度干旱胁迫下升高幅度较大,达17.45%,表明中度干旱胁迫即可对其造成损伤,在严重干旱胁迫下上升幅度为11.00%,均与对照有显著性差异,山西乌头的细胞膜透性在严重干旱胁迫下反而小于中度干旱胁迫,说明该乌头对中度干旱胁迫反应较迅速,随着胁迫加剧很快能通过其他途径(如系统保护酶)来降低干旱的伤害,以适应或抵抗干旱。
逆境胁迫下,植物体内活性氧的积累与系统保护酶活性和抗氧化剂的含量直接相关,植物可以通过体内超氧化物歧化酶、过氧化氢酶、过氧化物酶来减少过氧化产物,这在喻晓丽等[16]的研究结果中也得以证明。牛扁的细胞膜透性随干旱胁迫加重依次增大,中度干旱和严重干旱胁迫下分别比对照增大了13.28%和23.38%,差异均达显著水平,反映出随着干旱胁迫的加剧,牛扁的细胞膜受害加重。只有高乌头和牛扁的细胞膜透性表现出随干旱胁迫加剧而逐渐增大的趋势,其余3种乌头在干旱胁迫下细胞膜透性变化不一。供试乌头的细胞膜透性在中度干旱胁迫下的反应较复杂,表现出显著减小、差异不显著、显著增大3种变化,说明供试乌头在中度干旱胁迫下做出的修复反应和修复过程比较复杂,不同种之间也存在差异,5种乌头的细胞膜透性在严重干旱胁迫下均显著增大,反映出严重干旱胁迫对乌头细胞膜的伤害严重。
3.2 干旱胁迫对乌头丙二醛质量分数的影响
不同水分处理对乌头丙二醛(MDA)质量分数的影响见图2。
从图2(a)可以看出:北乌头的MDA质量分数在对照条件下随时间推移变化最小,其余各乌头在对照条件下的MDA质量分数变化也较平缓,基本上均缓慢升高,华北乌头在对照条件下20 d和30 d时的MDA质量分数几乎相等;除北乌头外,其余乌头在干旱胁迫下的MDA质量分数均表现出随时间推移而逐渐升高的趋势,反映出随着干旱的持续,乌头细胞膜脂过氧化作用增大,北乌头在严重干旱处理后20 d时的MDA质量分数最低,30 d时又升高,表现出其对干旱胁迫的反应速度较快,但可持续修复的时间较短。
从图2(b)可知:北乌头的MDA质量分数在中度干旱胁迫下显著上升,比对照升高32.42%,增幅最大,在严重干旱胁迫下无显著变化,反映出中度干旱胁迫即对北乌头细胞造成伤害,但其随即做出修复反应,本试验设计的严重干旱胁迫在其可修复范围;山西乌头的MDA质量分数在中度干旱胁迫下比对照有所升高,但差异不显著,说明中度干旱胁迫下山西乌头细胞膜脂过氧化作用不明显,在严重干旱胁迫下比对照升高43.94%,与对照差异达显著水平,说明严重干旱胁迫显著增大了山西乌头的细胞膜脂过氧化作用;高乌头、牛扁、华北乌头的MDA质量分数在中度干旱胁迫下分别比对照升高28.38%、28.79%、10.40%,在严重干旱胁迫下分别比对照升高68.47%、68.08%、46.77%。这3种乌头的MDA质量分数均表现出随干旱胁迫加剧而逐渐升高的趋势,且在3种水分处理条件下的差异均达显著水平,反映出随着干旱胁迫加剧,这3种乌头的细胞膜脂过氧化作用显著增大,细胞膜受害加重。
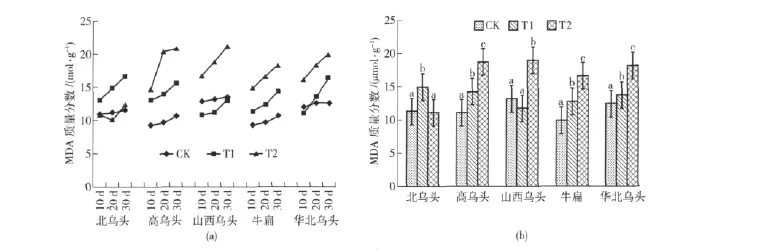
图2 不同水分处理对乌头MDA质量分数的影响Fig.2 Impacts of different water stress treatments on contents of MDA in different species
3.3 干旱胁迫对乌头叶绿素质量分数的影响
干旱条件下叶绿体片层膜系统受损,严重干旱时可观察到叶绿体变形和片层结构受到破坏[17],从而影响叶绿素质量分数。从表1可知:北乌头的叶绿素a(Chla)质量分数随干旱胁迫依次极显著递减,叶绿素b(Chlb)质量分数在3种水分处理间差异均达极显著水平(P<0.01),且变化幅度最大,中度干旱和严重干旱胁迫下分别比对照下降了70.16%和53.82%,Chla与Chlb的比值在中度干旱胁迫下为最大值4.57,在严重干旱胁迫下其比值也高于对照;高乌头的Chla质量分数在干旱胁迫下极显著低于对照,但中度干旱和严重干旱胁迫间无显著差异,Chlb质量分数在干旱胁迫下极显著高于对照,且在中度干旱胁迫下最高,中度干旱和严重干旱胁迫之间差异显著,Chla与Chlb的比值在对照条件下最高,比值为2.67;山西乌头的Chla和Chlb质量分数均随干旱胁迫依次极显著递减,Chla与Chlb的比值依次递增,严重干旱胁迫下最高,比值为3.19;牛扁的Chla和Chlb质量分数均随干旱胁迫依次递增,差异达显著或极显著水平,Chla与Chlb的比值在中度干旱胁迫下最高,比值为3.4,这种现象可能与干旱胁迫影响植物的氮代谢及体内物质的运输和转化有关,致使叶绿素的合成物质(尤其是L-谷氨酸)[18]增加而使其叶绿素质量分数上升,随之光合速率上升,但叶绿素的生物合成过程十分复杂,其中某些步骤迄今尚未明确[19];华北乌头的Chla质量分数在干旱胁迫下极显著低于对照,但在严重干旱胁迫下极显著高于中度干旱胁迫,Chlb质量分数在中度干旱胁迫下最高,极显著高于对照和严重干旱胁迫,对照和严重干旱胁迫下的Chlb质量分数则无显著差异,Chla与Chlb的比值变化幅度不大,在对照条件下最高,比值为1.63。

表1 不同水分处理对乌头Chla、Chlb和Chla与Chlb比值的影响Tab.1 Impacts of three water stress treatments on contents of Chla,Chlb and ratio of Chla to Chlb in different species
3.4 干旱胁迫对乌头光合速率的影响
干旱胁迫条件下,光合作用减弱的原因主要有2方面:气孔限制和非气孔限制[20]。气孔是CO2和水汽逸出的通道,植物缺水时气孔开度减小,明显限制CO2的供应,因而光合作用减弱,阴生植物更敏感。另有研究者发现,CO2供应的增加并不能降低对光合作用的抑制,因而,推测水分胁迫能引起叶绿体光合活性的改变和叶绿素含量的减少,以及其他非气孔因素也影响着光合作用,如叶绿体的希尔反应减弱,离体叶绿体放氧率降低,光系统Ⅱ的活力下降,光合电子传递和光合磷酸化受抑制,RuBP羧化酶和PEP羧化酶活力下降,水解加强,糖类积累等[18]。
受旱时植物光合作用的降低是由于气孔因素还是非气孔因素的限制,通常的判断依据是胞间CO2摩尔分数和气孔限制值的变化方向,胞间CO2摩尔分数降低和气孔限制值升高表明光合速率的降低是由于气孔因素的限制;相反,光合速率的降低伴随有胞间CO2摩尔分数的升高,则主要限制因素是非气孔因素[21]。
根据这一观点判定,北乌头和山西乌头在干旱胁迫下叶绿素质量分数逐渐降低,胞间CO2摩尔分数降低,气孔限制值升高,北乌头的净光合速率降低是受到气孔和非气孔2方面因素的限制,但以前者为主;高乌头随着干旱的加剧,Chla质量分数降低而Chlb质量分数升高,胞间CO2摩尔分数降低,气孔限制值升高,故气孔因素对其净光合速率降低的影响更大;牛扁在干旱胁迫下净光合速率的变化和其叶绿素质量分数的变化一致,均在干旱条件下上升,胞间CO2摩尔分数和气孔限制值却变化不大,说明牛扁净光合速率的变化主要由非气孔因素决定;华北乌头在干旱胁迫下叶绿素质量分数基本呈下降趋势,而胞间CO2摩尔分数、气孔限制值和净光合速率变化均不大,这可能是因为随着干旱胁迫加剧,植物体内产生一些渗透调节物质来维持植物水势平衡,从而保持一定气孔开度,说明气孔因素是影响华北乌头净光合速率的主要因素。

表2 不同水分处理对乌头不同光合指标的影响Tab.2 Impacts of three water stress treatments on different photosynthetic indices of different species
3.5 抗旱性综合评价
由表3比较结果可以看出,山西乌头、牛扁的抗旱隶属度随着干旱胁迫程度的加重逐步增大,表现出其对干旱胁迫较强的适应性和抵抗性,而高乌头、北乌头、华北乌头的抗旱隶属度却随胁迫程度而减小,尤其是华北乌头降幅超过50%,表明其抗旱能力逐步减弱。在本文设计的干旱胁迫条件下,5种乌头的抗旱性由大到小顺序为北乌头、高乌头、山西乌头、华北乌头、牛扁。

表3 乌头的隶属函数值Tab.3 Subordinate function values of five species
4 结论
1)北乌头在中度干旱胁迫下短期修复干旱损伤的能力较强,高乌头和华北乌头的细胞膜在中度干旱胁迫下没有明显伤害,山西乌头和牛扁的细胞膜在中度干旱胁迫下即受到伤害;5种乌头的细胞膜在严重干旱胁迫下均受到不同程度的伤害。5种乌头细胞膜透性由大到小的顺序为牛扁、山西乌头、高乌头、华北乌头、北乌头。
2)北乌头只在中度干旱胁迫下细胞膜脂过氧作用显著增大,细胞膜受到伤害,山西乌头在中度干旱胁迫下膜脂过氧化作用不明显,在严重干旱胁迫下细胞膜脂过氧化作用才显著增大,干旱胁迫下,高乌头、牛扁、华北乌头的细胞膜脂过氧化作用均依次显著增大,细胞受害依次加重。干旱胁迫下,5种乌头MDA质量分数的变化幅度大于细胞膜透性的变化幅度。干旱胁迫下5种乌头细胞膜脂过氧化作用由大到小的顺序为牛扁、高乌头、华北乌头、山西乌头、北乌头。
3)除牛扁的叶绿素质量分数在干旱胁迫下上升外,其余4个种的Chla质量分数在2种干旱胁迫条件下均低于对照,且除高乌头外,不同胁迫程度对乌头Chla质量分数的影响差异也较大;干旱胁迫对5种乌头Chlb质量分数的影响结果不一,有增有减,北乌头、牛扁的Chla与Chlb的比值在中度干旱时达最大值,高乌头和华北乌头则在中度干旱时最小,山西乌头的Chla与Chlb的比值随着干旱胁迫的加重依次增大。
4)干旱胁迫显著降低了北乌头、高乌头的净光合速率,但胁迫程度间差异不显著,除山西乌头和牛扁的净光合速率在中度干旱胁迫下升高外,其余乌头的净光合速率都随干旱胁迫程度呈下降趋势,干旱胁迫对华北乌头的净光合速率没有显著影响。北乌头、高乌头、山西乌头、华北乌头的净光合速率变化主要是受气孔因素限制,牛扁的净光合速率变化则受非气孔因素限制。
5)5种乌头的抗旱性由大到小的顺序依次为北乌头、高乌头、山西乌头、华北乌头、牛扁。
5 讨论
1)膜系统通常被认为是干旱伤害的最初和关键部位。干旱引起的膜伤害主要是由于机体积累自由基,并由此引发或加剧了细胞的膜脂过氧化,这一点在许多研究[22]中得到证实。膜脂发生过氧化反应的氧化终产物为MDA,它会引起蛋白质、核酸等生命大分子的交联聚合,且具有细胞毒性。刘裕强等[23]研究也表明当MDA质量分数大量增加时,体内细胞受到较严重的破坏。但在本试验中,山西乌头的细胞膜透性在严重干旱胁迫下小于中度干旱胁迫,其在中度干旱胁迫下的MDA质量分数也不显著的低于对照,类似结果在相关研究[10,16]中也有报道。
结合各乌头干旱胁迫下细胞膜透性和MDA质量分数的变化,反映出细胞在干旱胁迫下的代谢(尤其是修复)过程十分复杂,MDA质量分数并不能完全反映细胞膜透性变化的程度。
2)一般阳生植物叶片的Chla与Chlb的比值约为3∶1,Chlb质量分数的相对提高有可能更有效的利用漫射光中较多的蓝紫光,Chla与Chlb的比值也能反映出叶片光合活性的强弱[24]。许多研究也证实干旱可以影响Chla,Chlb和Chla与Chlb的比值的变化,从而进一步影响光合作用[25-26]。但干旱胁迫对光合作用的影响颇为复杂,还涉及到植物本身的基因型差异、生长发育状况、土肥条件、生长环境等[27]。
3)该抗旱性综合评价方法在应用时要选择合适的研究方法和一定数量的指标体系。在本文设计的2种干旱胁迫条件下,通过对4个指标体系的测定分析表明,北乌头的抗旱性最大,但其与高乌头、华北乌头的抗旱性却随着胁迫程度的加重而减弱,山西乌头、牛扁的抗旱隶属度随着干旱胁迫程度加重而逐步增大,其对干旱胁迫有较强的适应性和抵抗性,抗旱潜力较大,故在乌头的育种、引种和其在干旱、半干旱地区推广过程中要综合选择。另外,要得到较为全面的评价结果,还可以同时对其系统保护酶活性[28]、羧化酶活性[29]、水势[30]等指标体系进行测定分析。
论文得到了韩有志、杨云龙、吕晋慧老师的热情帮助和建议,在此表示感谢!
[1]高文韬,王壮,王新波,等.乌头的研究进展[J].华北大学学报,2009,10(2):144-148
[2]张继,杨永利,姚健,等.多根乌头的药用资源研究[J].西北植物学报,2002,22(4):975-978
[3]胡兆勇,赵汝能.甘肃乌头属药用植物资源[J].兰州医学院学报,1990,16(2):90-92
[4]李英和,陈迪华.短距乌头根的两个新二萜生物碱[J].植物学报,1994,36(2):148-152
[5]陈瑛,王明奎,吴凤锷,等.工布乌头根中的二萜生物碱[J].植物学报,1994,36(12):970-972
[6]丁立生,陈瑛,王明奎,等.瓜叶乌头的二萜生物碱[J].植物学报,1994,36(11):901-904
[7]贾淑珍.麝香乌龙丸中乌头碱的提取[J].现代中西医结合杂志,2011(4):472-473
[8]王会斌.黄花乌头的种子育苗技术[J].特种经济动植物,2004(1):23
[9]于荣敏,徐秀泉,赵昱,等.不同理化因子对黄花乌头花药愈伤组织诱导的影响[J].暨南大学学报:自然科学版,2003,24(3):109-114
[10]王宇超,王得祥,彭少兵,等.干旱胁迫对木本滨藜生理特性的影响[J].林业科学,2010,46(1):61-65
[11]高俊凤.植物生理学实验技术[M].西安:世界图书出版公司,2000:31-210
[12]邹琦.植物生理学实验指导[M].北京:中国农业出版社,2001:1-198
[13]李玉灵,朱帆,王俊刚,等.水分胁迫下臭柏光合特性和色素组成的季节变化[J].生态学报,2009,20(8):4347-4352
[14]Berry J A,Downton W J S.Environmental regulation of photosynthesis∥ Govindjee. Photosynthesis,Development,Carbon Metabolism and Plant Productivity[M].New York:Academic Press,1982:263-343
[15]江天然,张立新,毕玉蓉,等.水分胁迫对梭梭叶片气体交换特征的影响[J].兰州大学学报:自然科学版,2001,37(6):57-62
[16]喻晓丽,邸雪颖,宋丽萍.水分胁迫对火炬树幼苗生长和生理特性的影响[J].林业科学,2007,43(11):57-61
[17]Bailly C,Benamar A,Corbineau F,et al.Changes in malondialdehyde content and in superoxide dismutase,catalase and glutathione reductase activities in sunflower seed as related todeterioration during accelerated aging[J].Plant Physiology,1996,97:104-110
[18]王忠.植物生理学[M].北京:中国农业出版社,2000:131
[19]张继澍.植物生理学[M].北京:高等教育出版社,2006:20-414
[20]王晓琴,袁继超,熊庆娥.玉米抗旱性研究的现状及展望[J].玉米科学,2002,10(1):7-60
[21]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annu Rev PlantPhysiology,1982,33:317-345
[22]陈少瑜,郎南军,贾利强,等.干旱胁迫对坡柳等抗旱树种幼苗膜脂过氧化及保护酶活性的影响[J].植物研究,2006,26(1):88-92
[23]刘裕强,江玲,孙立宏,等.褐飞虱刺吸诱导的水稻一些防御性酶活性的变化[J].植物生理与分子生物学学报,2005,31(6):643-650
[24]曹晶,姜卫兵,翁忙玲,等.夏秋季旱涝胁迫对红叶石楠光合特性的影响[J].园艺学报,2007,34(1):163-172
[25]应小芳,刘鹏.铝胁迫对大豆叶片光合特性的影响[J].应用生态学报,2005,16(1):166-170
[26]Downton W J S,Grant W J,Robbinson S P.Stomatal closurefully accounts for the inhibition of photosynthesis by abscisic acid[J].Plant Physiology,1985,77:85-88
[27]柯世省.干旱胁迫对夏蜡梅光合特性的影响[J].西北植物学报,2007,27(6):1209-1215
[28]Alskog G,Huss-Danell K.Catalase and nitrogenase activities of symbiotic Frankin in response to different oxygen tensions[J].Plant Physio,1997,99:286-292
[29]徐俊增,彭世彰,丁家丽,等.控制灌溉的水稻气孔限制值变化规律试验研究[J].水利学报,2006,30(4):486-491
[30]蔡昆争,吴学祝,骆世明,等.不同生育期水分胁迫对水稻根系活力、叶片水势和保护酶活性的影响[J].华南农业大学学报,2008,29(2):7-10
