自然降温下4个狗牙根品种(系)的生理指标比较
2011-04-25李秋丽包满珠王文恩
李秋丽,包满珠,王文恩
(华中农业大学园艺林学学院,湖北 武汉 430070)
狗牙根(Cynodondactylon)是典型的暖季型草坪草之一,常在冬季不耐寒而枯黄。因此,低温是限制其推广应用的主要因素[1-2]。近年来,已有不少有关狗牙根耐寒性的研究报道,主要对人工低温胁迫下狗牙根的生理生化指标进行比较研究,并取得了一定的进展[3-5]。本研究以华中农业大学通过辐射诱变选育的狗牙根新品系‘矮败狗牙根’和田间筛选的优良狗牙根无性系‘HN019’为对象,与目前草坪工程中广泛应用的‘天堂419’和‘公主’作比较,在露地栽培条件下的自然降温过程中对其抗寒性进行分析和比较,从各品种(系)的相关生理指标对露地栽培下自然降温过程中的变化来评价其耐寒性,以期筛选出耐寒性较好的品种(系),为抗寒性暖季型草坪草的选育工作提供理论依据和参考。
1 材料与方法
1.1试验材料 研究对象是华中农业大学通过60Co-γ射线辐射诱变选育出的‘矮败狗牙根’和田间筛选的优良狗牙根无性系‘HN019’,与目前草坪工程中广泛应用的狗牙根品种‘天堂419’(Tifway)和‘公主’(Princess)作比较。4种材料均种植在华中农业大学花卉基地草坪品种圃。
1.2试验方法 2009年10-12月,记录每天的气温变化(图1)。试验期间温度整体呈下降趋势,其中10月30日的温度骤然升高,当天的最低温度为19℃;11月3日的温度骤然降低,最低温度为3℃;11月3日以后,最低温度均在12℃以下,其中12月5日的最低温度为1℃。试验期间,每隔10 d从品种圃随机剪取4个狗牙根品种的嫩叶,分别对各品种叶片的电导率值、丙二醛(MDA)含量、脯氨酸(Fpro)含量、可溶性糖(WSS)含量、可溶性蛋白(SP)含量、叶绿素(Chl)含量和超氧化物歧化酶(SOD)活性进行测定,3次重复。电导率测定采用电导法[6],MDA含量采用硫代巴比妥酸法[7],Fpro含量采用酸性茚三酮法[7],WSS含量采用蒽酮比色法[7],SP含量采用考马斯亮蓝G-250染色法[7],Chl含量采用95%乙醇热浸法[8],SOD活性采用SOD试剂盒[7](南京建成生物工程研究所生产)。
1.3模糊隶属函数法 为了全面利用各指标对植物的抗寒性进行综合评价,采用模糊数学隶属函数法对各指标测定值进行定量转换[8]。
如果指标与抗寒性呈正相关:
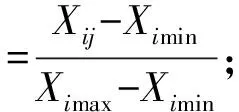
如果指标与抗寒性呈负相关:
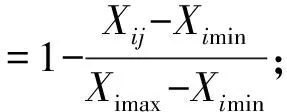
式中,Zij为i品种j指标的抗寒性隶属函数值,Xij为i品种j指标的测定值,Ximax和Ximin为各品种中指标的最大和最小测定值。
将植物各指标的隶属函数值累加起来,求其平均值,隶属函数值越大,该品种的抗寒性就越强。

图1 试验期间日均温变化
1.4数据分析 用Excel 2003和SAS统计分析软件进行方差分析,用Excel 2003制图,采用模糊隶属函数法综合评价出各品种(系)的耐寒性强弱。
2 结果与分析
2.1电导率的变化 在自然降温过程中4个狗牙根品种(系)的电导率变化比较一致,呈先升高后降低再升高的趋势(图2)。在降温前期10月至11月初,电导率上升较缓慢,11月15日的电导率值增幅变大,其中‘公主’的电导率值增幅最大,达106.82%,‘矮败狗牙根’、‘HN019’和‘天堂419’的增幅分别为84.29%、61.06%和67.36%。11月25日电导率值有所下降,在12月15日各品种(系)的电导率值达最高峰,‘公主’的峰值最大,为30.13%,‘矮败狗牙根’的峰值较小,为25.16%(图2)。通过对各品种(系)的电导率均值进行差异显著性比较发现,在整个自然降温过程中,‘公主’和‘天堂419’的电导率均值显著高于‘HN019’和‘矮败狗牙根’(P<0.05), 其中‘矮败狗牙根’的电导率值显著低于其他品种(系)(表1)。
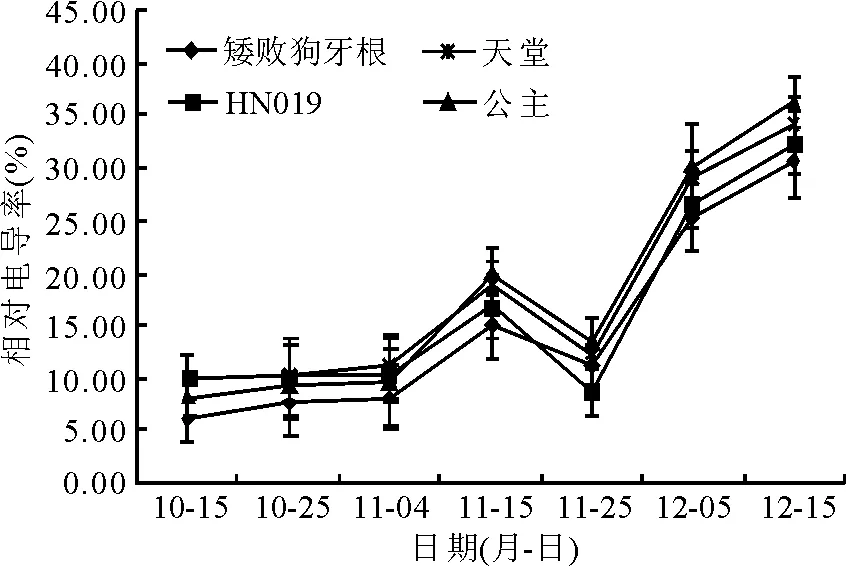
图2 露地栽培下狗牙根相对电导率的变化
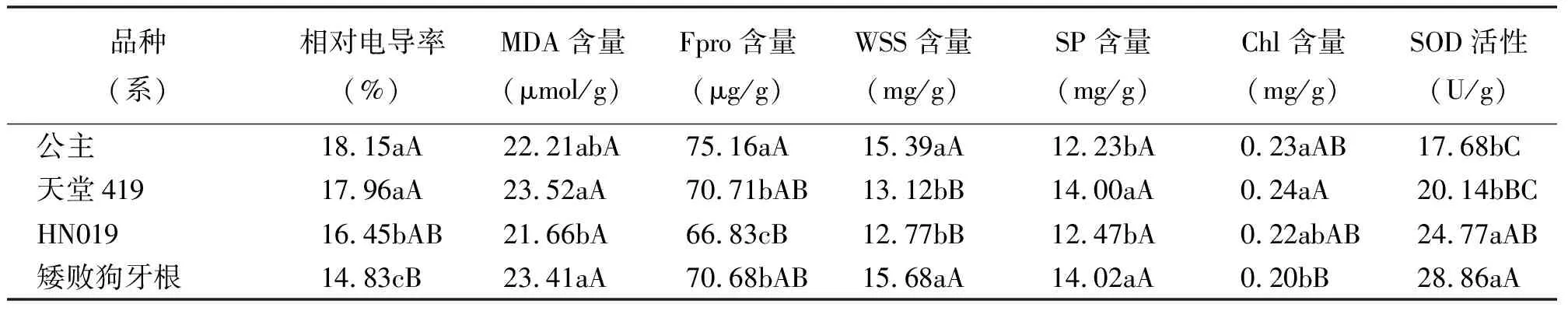
表1 狗牙根品种(系)间生理指标比较
2.2丙二醛含量的变化 在自然降温过程中,4个狗牙根品种(系)的MDA含量总体呈缓慢上升的趋势(图3)。在降温初期10月15日-11月4日,MDA含量上升缓慢,11月15日时MDA含量增幅变大,‘天堂419’和‘矮败狗牙根’的增幅为47.03%和40.73%,大于‘公主’和‘HN019’的增幅。在11月15日之后,MDA含量出现小的下降然后平缓上升,于12月15日达到最高峰(图3)。通过对各品种(系)的MDA均值进行差异显著性比较发现,在整个自然降温过程中,‘天堂419’、‘矮败狗牙根’、 ‘公主’三者之间的MDA含量差异不显著(P>0.05),‘天堂419’和‘矮败狗牙根’的MDA含量显著高于‘HN019’(P<0.05)(表1)。

图3 露地栽培下狗牙根丙二醛含量的变化
2.3脯氨酸含量的变化 在自然降温过程中,4个狗牙根品种(系)的Fpro含量变化呈先上升又缓慢下降的趋势(图4)。在降温前期10月-11月初,各品种(系)的Fpro含量增幅较大,11月4日-11月15日Fpro含量变化平缓,11月15日各品种(系)的Fpro含量达到最大值,其中‘公主’的峰值最大,为92.56μg/g, ‘天堂419’、‘矮败狗牙根’和‘HN019’的峰值分别为90.85、90.11和86.12 μg/g(图4)。通过对各品种(系)的Fpro均值进行差异显著性比较发现,在整个自然降温过程中,‘公主’的Fpro含量显著高于其他品种(P<0.05),‘HN019’ 的Fpro含量显著低于其他品种(P<0.05),‘天堂419’和‘矮败狗牙根’的的Fpro含量差异不显著(P>0.05)(表1)。

图4 露地栽培下狗牙根脯氨酸含量的变化
2.4可溶性糖含量的变化 在自然降温过程中,4个狗牙根品种(系)的WSS含量呈现出先缓慢升高又下降再升高再下降的趋势(图5)。降温初期各品种(系)的WSS含量增长较慢,‘天堂419’和‘HN019’的WSS含量在11月4日达到第1个峰值,分别为15.48和17.16 mg/g,‘矮败狗牙根’和‘公主’在11月15日达到第1个峰值,分别为21.10和18.85 mg/g。随后WSS含量有一定程度的下降,各品种(系)的WSS含量均在12月5日达到了第2个峰值,随后逐渐下降。通过对各品种(系)的WSS均值进行差异显著性比较发现,在整个自然降温过程中,‘矮败狗牙根’和‘公主’的WSS含量差异不显著(P>0.05),‘天堂419’和‘HN019’的WSS含量差异不显著(P>0.05),‘矮败狗牙根’和‘公主’的WSS含量极显著高于‘天堂419’和‘HN019’的WSS含量(P<0.01)(表1)。
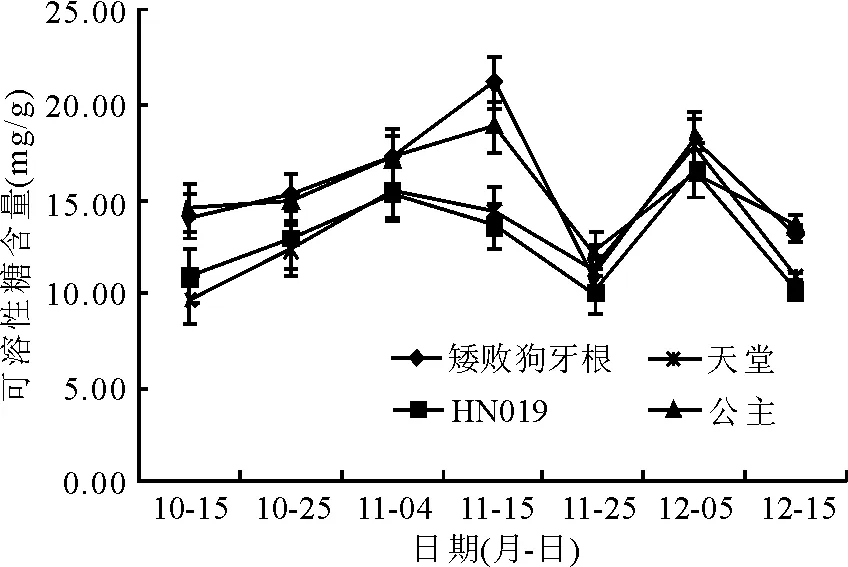
图5 露地栽培下狗牙根可溶性糖含量的变化
2.5可溶性蛋白含量的变化 在自然降温过程中,4个狗牙根品种(系)的SP含量变化呈现出先迅速升高又迅速下降再缓慢升高的趋势(图6)。4个狗牙根品种(系)的SP含量均在11月4日时迅速升高并且到达峰值,‘矮败狗牙根’和‘天堂419’的峰值较大,分别为23.50和23.28 mg/g。11月4日-11月15日各品种(系)的SP含量迅速下降,11月15日-12月15日的降温期间SP含量又逐渐升高(图6)。通过对各品种(系)的SP均值进行差异显著性比较发现,‘矮败狗牙根’和‘天堂419’ 的SP含量显著高于 ‘HN019’和‘公主’的SP含量(P<0.05)(表1)。

图6 露地栽培下狗牙根可溶性蛋白含量的变化
2.6叶绿素含量的变化 在自然降温过程中,4个狗牙根品种(系)的Chl含量变化表现一致,总体呈现出曲折下降的趋势(图7)。在降温初期10月15日-11月4日,各品种(系)的Chl含量先上升后下降,11月15日随着温度的回升Chl含量也有一定的增加,11月15日之后随着低温的加剧Chl含量呈现逐步下降的趋势。可以看出,在整个自然降温过程中,‘天堂419’的Chl含量相对处于较高水平,‘矮败狗牙根’的Chl含量一直较低,‘HN019’和‘公主’的Chl含量处于中间水平(图7)。通过对各品种(系)的Chl均值进行差异显著性比较发现,‘天堂419’和‘公主’的Chl含量高于‘HN019’和‘矮败狗牙根’,但前两者与‘HN019’差异不显著(P>0.05),与‘矮败狗牙根’差异显著(P<0.05)(表1)。

图7 露地栽培下狗牙根叶绿素含量的变化
2.7超氧化物歧化酶活性的变化 在自然降温的过程中,4个狗牙根品种(系)的SOD活性变化呈现出先上升后下降的趋势(图8)。10月15日-11月4日的降温过程中,各品种(系)的SOD活性急速上升,11月4日-11月15日随着温度的回升SOD活性又急速下降,随后SOD活性随着低温的加剧而缓慢下降。各品种(系)的SOD活性于11月4日达到最高峰值,‘矮败狗牙根’和‘天堂419’的峰值最大,分别为51.86和51.08 U/g,‘公主’和‘HN019’的峰值分别为38.11和39.34 U/g。可以看出,在整个自然降温过程中‘矮败狗牙根’的SOD活性一直处于较高水平,11月4日之后‘HN019’的SOD活性也随着低温加剧而保持较高水平,‘天堂419’(除11月4日外)和‘公主’的SOD活性在整个降温过程中相对较低(图8)。通过对各品种(系)的SOD活性均值进行差异显著性比较发现,在整个自然降温过程中,‘矮败狗牙根’和‘HN019’的SOD活性显著高于‘天堂419’和‘公主’(P<0.05)(表1)。
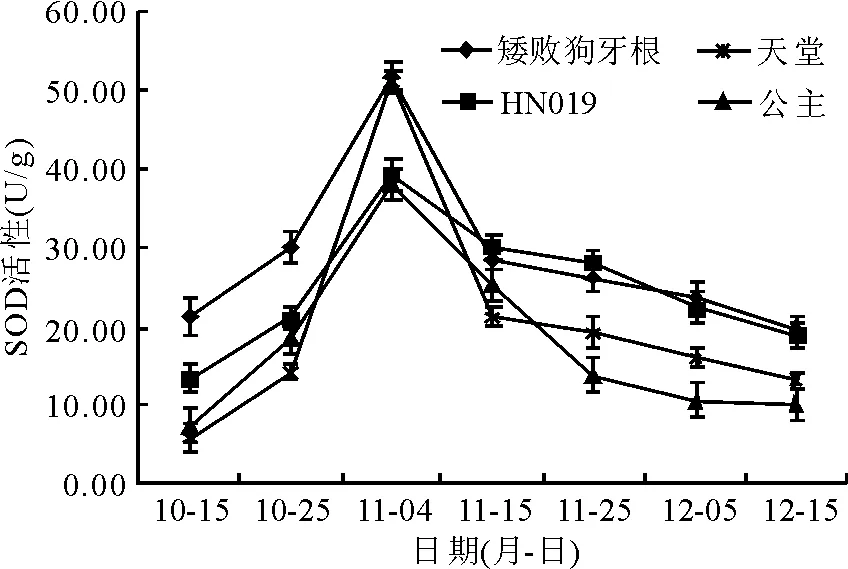
图8 露地栽培下狗牙根SOD活性的变化
2.8模糊隶属函数法综合评价 ‘矮败狗牙根’的隶属度平均值最大,为0.645 7,其次为‘公主’和‘天堂419’,隶属度分别为0.477 8和0.407 4,‘HN019’的隶属度较小,为0.379 2(表2)。由此可以得出4个狗牙根品种(系)的耐寒性强弱顺序为‘矮败狗牙根’>‘公主’>‘天堂419’>‘HN019’。
3 讨论与结论
细胞膜是细胞与外界环境发生物质交换、能量转换、信号传递等生命活动的重要场所和媒介,也是细胞感受环境胁迫最敏感的部位[9]。植物受到低温胁迫时,细胞质膜透性会有不同程度增大,电解质会不同程度外渗,导致电导率升高[10]。丙二醛是细胞膜脂过氧化产物之一,能与细胞内部各种成分发生强烈的反应而引起酶和膜的严重损伤,导致结构和生理完整性的破坏,是膜系统受伤害的重要指标[11-12]。
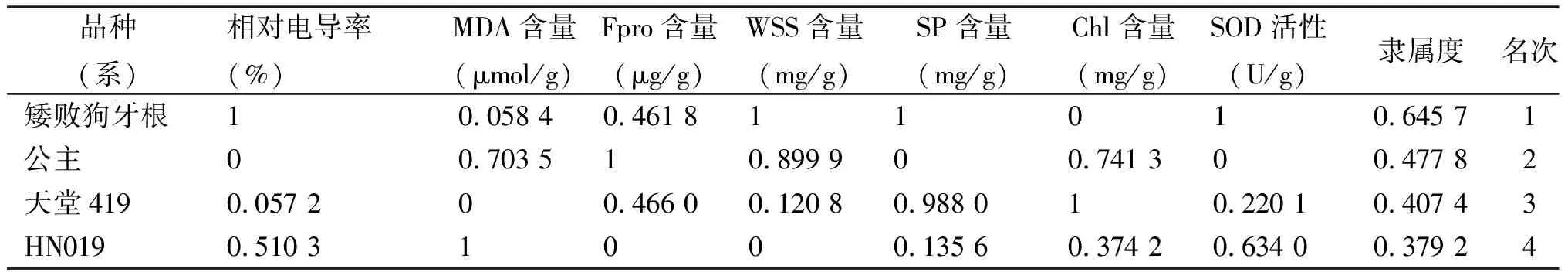
表2 露地栽培条件下各狗牙根材料耐寒性综合评价
本研究通过对电导率和丙二醛含量的测定,发现4个狗牙根品种(系)的电导率随着温度的降低整体呈现出增大趋势,丙二醛含量随着温度的降低总体呈现上升后下降趋势,耐寒性强的的品种(系)电导率较小,丙二醛含量较低,表明其细胞受损程度较小。在11月15日温度回升期,各狗牙根品种电导率出现缓慢下降,可能是温度回升使植株对低温有了一定的适应。但随着12月低温的到来,电导率值出现了迅速增高的现象,表明此时植物随着低温的加强受到的伤害加剧。丙二醛含量出现小的下降然后平缓上升,这与电导率表现一致,说明植物体内丙二醛对温度的变化反应敏感,并随着温度的下降含量逐渐增高。
一般认为脯氨酸在植物抵抗低温胁迫时起到平衡细胞代谢的作用,以保持细胞环境的相对稳定,其含量的增加有利于提高抗寒性[13]。低温胁迫下脯氨酸含量逐渐增加,可以降低细胞水势,以此避免冻害的发生[14-15]。可溶性糖能够增加细胞液浓度,降低冰点,并与水结合,会使细胞内束缚水含量增高,增强细胞液的流动性,原生质粘性增强,细胞耐结冰脱水能力和耐冻性增强[16]。可溶性糖含量越高,植物组织细胞液浓度也越高,植物耐寒能力越强[17]。本研究表明,脯氨酸含量随着低温的到来呈现先上升后下降的趋势,可溶性糖含量随着低温的到来呈现整体上升又下降的趋势,耐寒性强的品种,脯氨酸和可溶性糖含量较大。11月4日-11月15日的温度回升期,各品种(系)的可溶性蛋白含量迅速下降,11月15日-12月15日的降温期间可溶性蛋白含量又开始升高,表明植物体内可溶性蛋白对温度的变化反应敏感。各品种(系)的可溶性蛋白含量在11月4日和12月15日的低温期出现两次峰值,说明植物体内可溶性蛋白随着温度的变化做出了适应。
低温对叶绿素影响有两个方面,一是叶绿素的生物合成过程绝大部分有酶参与,低温影响酶的活性,进而影响叶绿素的合成;二是低温作用于植物,使植物本身形成水分逆境,因组织缺水引起气孔关闭,叶绿体受伤,与光合作用有关的酶失活或变性[18-19]。因此在植物耐寒生理研究中,叶绿素含量常作为反映低温胁迫对植物造成伤害程度的生理指标。本研究表明,在自然降温过程中,叶绿素含量随着温度的降低总体呈现下降的趋势,耐寒性强的品种,叶绿素含量较高。
超氧化物歧化酶是防御细胞膜系统受活性氧损害的保护酶之一[20],能以超氧阴离子为基质进行歧化反应,进而清除植物组织和细胞内的超氧阴离子自由基,同时产生歧化物H2O2,从而减缓超氧化物自由基对细胞膜的伤害。低温下植物体内SOD酶活性的变化与其抗寒性密切相关,超氧化物歧化酶是植物抗氧化系统的第一道防线,是酶促清除系统中的核心酶[21-22]。本研究表明,超氧化物歧化酶活性随着温度的降低呈先上升又下降的趋势,耐寒性强的品种,超氧化物歧化酶活性相对较高。在降温初期各品种(系)的超氧化物歧化酶活性急速上升,温度回升期又急速下降,随后超氧化物歧化酶活性随着低温的加剧而缓慢下降,表明植物内超氧化物歧化酶对低温反应敏感,各品种(系)在11月4日达到最高峰,表明11月4日的低温已是植物体内超氧化物歧化酶能适应的极限温度。
在整个自然降温过程中,4个狗牙根品种(系)的生理生化指标变化趋势比较一致,通过差异显著性分析可以看出品种上各生理指标存在显著差异。综合来看,‘矮败狗牙根’的电导率值最小,可溶性糖含量、可溶性蛋白含量和超氧化物歧化酶活性最大,‘公主’的脯氨酸、可溶性糖和叶绿素含量较大,‘天堂419’的丙二醛含量和叶绿素含量较大。‘HN019’的电导率值较大,脯氨酸和可溶性糖含量较小。在时间上可以看出各品种(系)在12月受低温伤害最严重。通过模糊隶属函数法综合评价得出,4个狗牙根品种(系)的耐寒性强弱顺序为‘矮败狗牙根’>‘公主’>‘天堂419’>‘HN019’。
[1] 郑玉红,刘建秀,陈树元,等.中国狗牙根[Cynodondactylon(L.)Pers.]耐寒性及其变化规律[J].植物资源与环境学报,2002,11(2):48-52.
[2] 刘玲珑,吴彦奇.狗牙根种质资源及抗寒性研究进展[J].中国草地,2000(6):45-50.
[3] 刘志雄,周高峰,廖启荣,等.低温胁迫对几个狗牙根品种生理生化特征的影响[J].安徽农学通报,2007,13(4):23-25.
[4] 张伟锋,陈平,姬承东.广州地区几种草坪草低温胁迫效应研究[J].草业科学,2006,23(8):85-88.
[5] 代会平,杨知建,胡勇.狗牙根抗寒性研究进展[J].草业科学,2007,24(11):87-91.
[6] 萧浪涛,王三根.植物生理学实验技术[M].北京:中国农业出版社,2006.
[7] 李合生.植物生理生化实验原理和技术[M].北京:高等教育出版社,2005.
[8] 邵文鹏.几种常绿阔叶植物抗寒性研究[D].青岛:山东农业大学,2009.
[9] Lyons J M.Chilling injury in plants[J].Annual Review of Plant Physiology,1973(24):445-448.
[10] 王洪春.植物抗逆性与生物膜结构功能研究进展[J].植物生理学通讯,1985(1):60-66.
[11] 郑小林,胡木林,罗晓莹,等.假俭草低温胁迫的伤害与适应[J].草业科学,2002,19(7):55-57.
[12] 张朝阳,许桂芳.利用隶属函数法对4种地被植物的耐热性综合评价[J].草业科学,2009,26(2):57-60.
[13] Dunn J H,Nelson C J.Chemical changes occurring in three bermudagrass turf cultivars in relation to cold hardiness[J].Agronomy Journa1,1974(6):28-31.
[14] 龚明,刘友良,朱培仁.低温下稻苗叶片中蛋白质及游离脯氨酸的变化[J].植物生理学通讯,1989(4):18-22.
[15] 陈杰忠,徐春香,梁立峰.低温对香蕉叶片中蛋白质及脯氨酸的影响[J].华南农业大学学报,1999,20(3):54-56.
[16] 宋海鹏,刘君,李秀玲,等.干旱胁迫对5种景天属植物生理指标的影响[J].草业科学,2010,27(1):11-15.
[17] 何若韫.植物低温逆境生理[M].北京:中国农业出版社,1995:13-35.
[18] 刘兵,张世权,郭家保.植物耐逆研究概况[J].安徽农学通报,2006,12(8):39-121.
[19] 杜秀敏,殷文漩.超氧化物歧化酶(SOD)研究进展[J].中国生物工程杂志,2003,23(1):48-51.
[20] 杨鑫光,傅华,牛得草.干旱胁迫下幼苗期霸王的生理响应[J].草业学报,2007,16(5):107-112.
[21] 王齐,孙吉雄,安渊.水分胁迫对结缕草种群特征和生理特性的影响[J].草业学报,2009,18(2):33-38.
[22] 王静,杨持.冷蒿耐寒生理特性的研究[J].内蒙古大学学报(自然科学版),2002,33(6):673-676.
