松嫩平原扁秆藨草种群生殖株构件表型可塑性及生长分析
2011-04-25张经经杨允菲陈永强
张经经,杨允菲,陈永强
(东北师范大学草地科学研究所 植被生态科学教育部重点实验室,吉林 长春 130024)
植物的生长发育及形态结构建成是一个顺序发生过程,但植物的物质生产和分配以及所形成的器官是否能够继续发育却具有较大的可调剂性,从而普遍出现表型可塑性[1]。表型可塑性是植物对环境的适应,具有调节生长和物质分配的功能和意义[2-5]。而籽实成熟期的不同植株,是生殖构件形成、生长和整个植株物质生产与分配长期调剂的结果[1]。因此,深入研究植物各数量性状的表型可塑性与植株物质生产和分配的内在联系及其变化规律,不仅是植物种群生活史策略的核心内容,也是植物种群生态学研究的重要方面。
扁秆藨草(Scirpusplaniculmis)是我国东北稻田边湿地和低湿草甸广泛分布的无性系植物,以往对扁秆藨草的研究主要集中在与土壤生境的相互关系[6-8]、无性繁殖规律[9]、危害及其防治[10]等方面,而有关种群生长与生殖分配的可塑性及规律性的研究,迄今尚未见报道。本研究根据对松嫩平原草甸生境野生扁秆藨草生殖株的大样本取样,就种群中的生殖株进行了生长和生殖分配的定量分析,旨在揭示种群物质和能量分配策略,为扁秆藨草的合理利用和有效防治提供理论依据,为植物种群生殖生态的深入研究奠定基础。
1 材料与方法
1.1研究区概况 研究地点位于松嫩平原南部、吉林省长岭种马场、东北师范大学草地生态研究站的天然碱化草甸(44°38′ N,123°41′ E)。该区属于温带半湿润、半干旱气候,年平均气温4.6~6.4 ℃,年平均降水量313~581 mm,主要集中在6-9月;年蒸发量约为降水量的2~3倍。无霜期136~163 d。6-8月平均气温超过20 ℃,7月气温最高[11-12]。
研究样地设在扁秆藨草种群呈镶嵌分布的杂类草群落低洼地段,其中心为扁秆藨草并伴有少量的鹅绒藤(Cynanchumchinense)、拂子茅(Calamagrostisepigeios)、星星草(Puccinelliatenuiflora)、虎尾草(Chlorisvirgata)、西伯利亚蓼(Polygonumsibiricum)等,边缘分布着较多的羊草(Leymuschinensis)、芦苇(Phragmitescommunis)和寸草苔(Carexduriuscula)。土壤为碱化草甸土。进入生长季中后期,扁秆藨草一般高50~80 cm。研究样地为割草场,每年8月上旬割草,备制冬贮干草。此时扁秆藨草处于籽实成熟期。
1.2研究材料 扁秆藨草是莎草科(Cyperaceae)多年生草本。根状茎具地下匍匐枝,其顶端变粗成块茎状。秆单一,高30~80 cm,较细,三棱形,平滑,具多数秆生叶。长侧枝聚伞花序短缩成头状或有时具1~2个短的辐射枝,通常具1~6个小穗;小穗卵形,具多数花;小坚果倒卵形或广倒卵形。块茎及根状茎含淀粉可造酒;块茎可代荆三棱供药用[7]。同时扁秆藨草也是我国稻田以无性繁殖方式为主的恶性杂草之一,严重危害水稻的生长[9]。
1.3取样与测定 2010年8月初,于扁秆藨草籽实成熟期,采用大样本取样测定方法,在10 m×10 m的样地内,对扁秆藨草种群的生殖株随机取样,样本数为30株。取样时,将每个生殖株齐地面剪下,分置,保持各株完整性。测定植株高度后于80 ℃烘干,分别测定植株生物量、花序生物量和种子生物量。
1.4数据处理 对所测的各数量性状作统计分析,各项测定数据均采用实测值。用Excel软件计算各项数量指标的平均数(Mean)、标准差(SD)和变异系数(CV)。性状间的关系,在线性、指数、乘幂和对数函数中,选择相关性最好的作为其规律性变化的描述模型。分别用花序生物量和种子生物量占株生物量的百分比作为生殖分配的数量指标。其计算公式如下:
2 结果与分析
2.1数量性状的表型可塑性 在籽实成熟期,扁秆藨草生殖株构件各数量性状的统计结果见表1。7个数量性状中,如果把变异系数和观测值的范围作为衡量表型可塑性的数量指标[1],则种子生物量的表型可塑性最大,其变异系数高达81.63%,最大值是最小值的19倍;然后依次是生殖分配Ⅱ、花序生物量、茎叶生物量、植株生物量和生殖分配Ⅰ,而以植株高度的表型可塑性最小,其变异系数为13.11%,最大值是最小值的1.6倍。可见,扁秆藨草种群生殖株不同个体生长与生殖分配均存在着不同程度的表型可塑性。
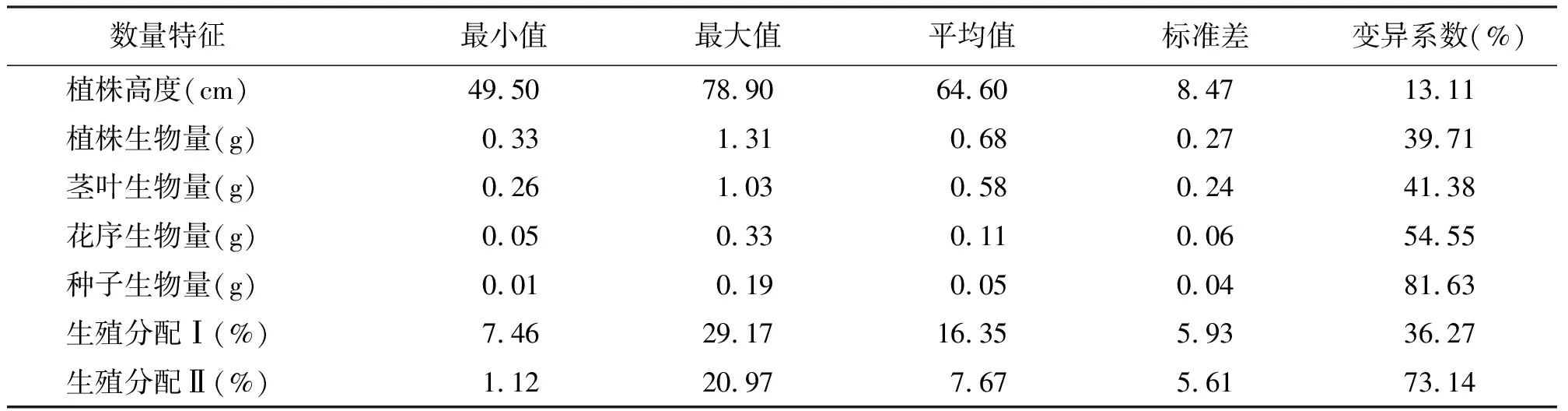
表1 扁秆藨草种群生殖株构件数量性状的统计值及观测值范围
2.2构件生物量与株高及株生物量的关系 经统计分析,在30个样本中,扁秆藨草种群生殖株的茎叶生物量分别与株高和株生物量呈极显著(P<0.01)的幂函数正相关关系;花序生物量与株生物量呈极显著(P<0.01)的线性正相关关系,而与株高呈幂函数增加的趋势(图1)。反映了扁秆藨草种群生殖株茎叶生物量与株高和株生物量之间具有异速增长规律,但是增长速率不尽相同;花序生物量与株高具异速生长规律,与株生物量具同速生长规律。说明扁秆藨草种群不同高度和不同生物量生殖株的养分积累及其向营养器官和生殖器官的分配是协调有序的,但所遵循的规律和速率不尽相同,具有两种不同过程的表型可塑性调节。
2.3生殖分配与株高及株生物量的关系 经统计分析,在30个样本中,扁秆藨草种群生殖株生殖分配Ⅰ与株高呈极显著(P<0.01)的指数函数负相关关系,与株生物量呈显著(P<0.05)的幂函数负相关关系;生殖分配Ⅱ与株高和株生物量分别呈极显著(P<0.01)的幂函数负相关关系和幂函数下降的趋势(图2)。表明生殖株株高和株生物量的增加要以减少其生殖分配为代价,将更多的能量投人到茎叶的营养生长上。
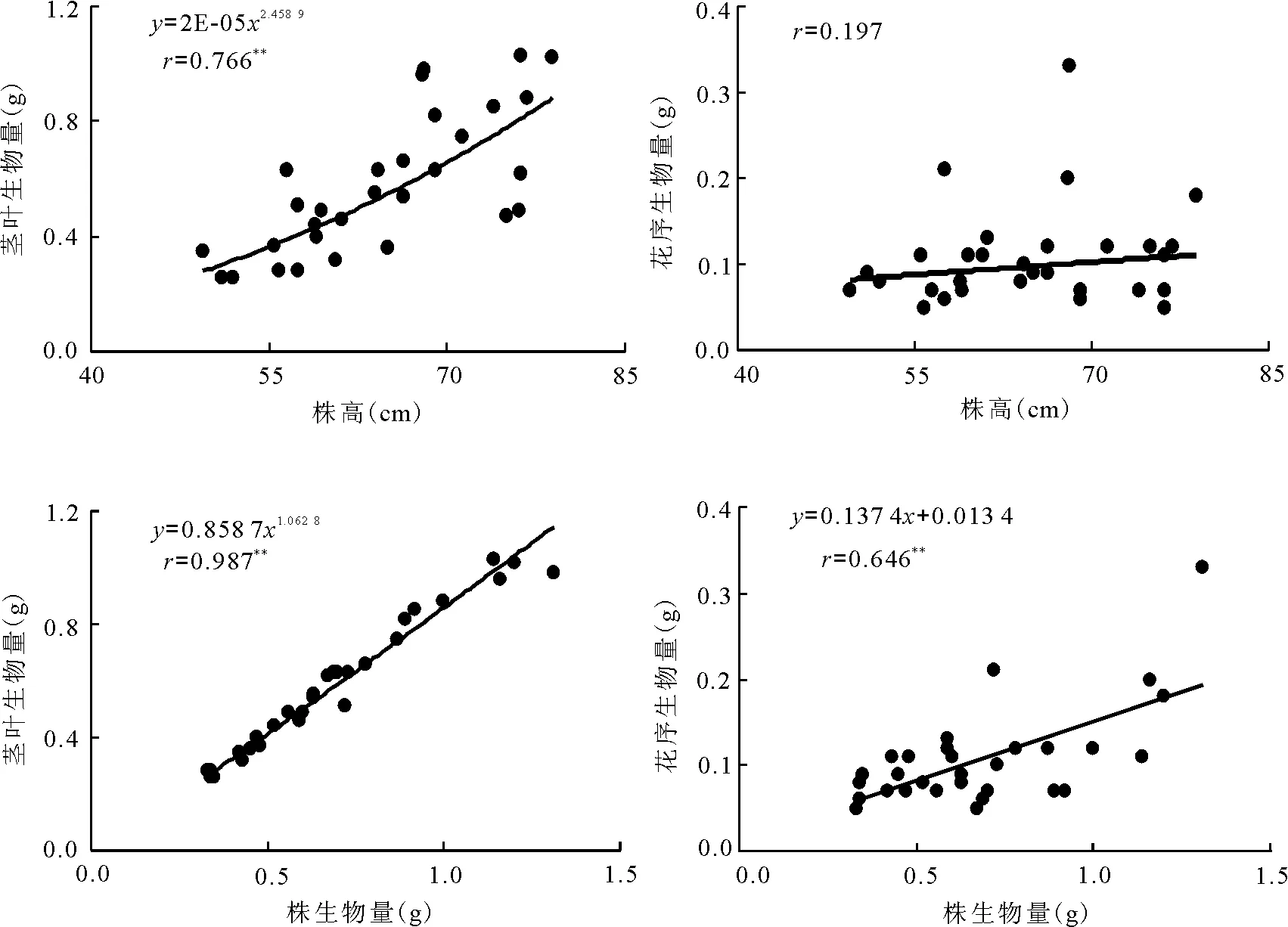
图1 扁秆藨草种群生殖株构件生物量与株高和株生物量的观测值及拟合曲线
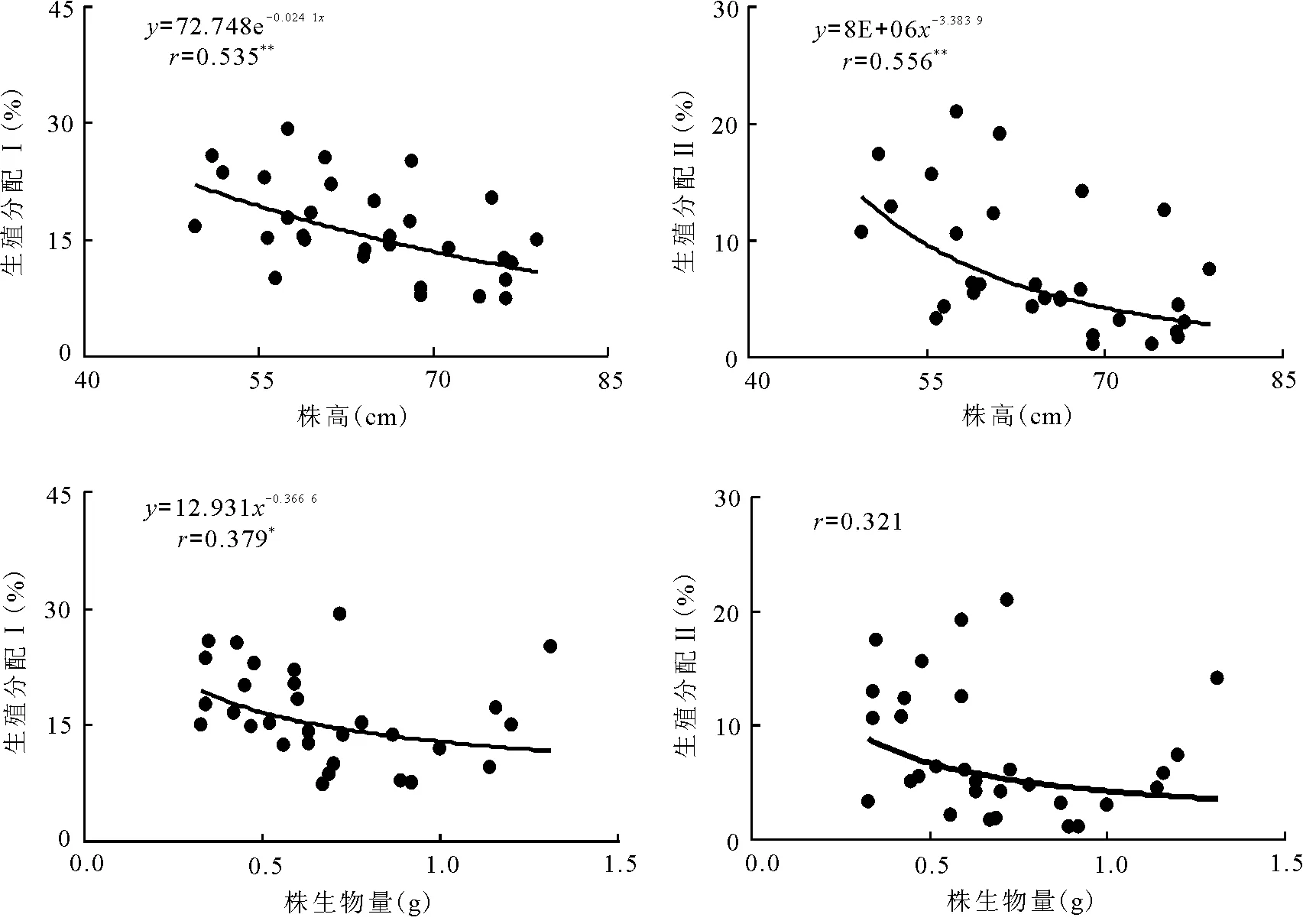
图2 扁秆藨草种群生殖株的生殖分配与株高和株生物量的观测值及拟合曲线
3 讨论与结论
3.1数量性状的表型可塑性 植物的表型是基因型与环境共同作用的结果,相同基因型控制的性状,因环境条件的差异而存在着可塑性。在理论上,同质环境不存在表型可塑性,但天然群落即使在极小范围的样地内,表观植被组成与分布已大体一致,也存在微地形、土壤质地的均匀性、土壤小动物的干扰、不同植株返青的时间等差异。所以,天然植物在异质群落内存在表型可塑性,在相对同质的群落内也存在表型可塑性[1]。植物数量性状的表型可塑性,是植物对环境应力的适应性反应,适应性越广,生态幅度越大,其可塑性也越大[13]。在松嫩平原,扁秆藨草生殖株构件在籽实成熟期,如果用变异系数来衡量各数量性状的表型可塑性,其最小为13.11%,最大为81.63%,反映出扁秆藨草种群个体生长与分配存在环境效应。对于这种环境效应是微环境所致,还是个体出生和生长时间的差异所致有待于进一步研究。
3.2株高与株生物量对构件生物量的影响 植物种群生殖株的地上生物量包括茎叶生物量和花序生物量。植物生殖株株高不同,分配给营养器官和生殖器官的生物量也不尽相同,即随着生殖株株高和株生物量的增加,营养器官和生殖器官的生物量增加规律及相同规律的增长速率均不尽相同,因而表现出不同的生长型,即同速生长型和异速生长型。同速生长型往往表现在数量性状间的关系为线性函数形式,而异速生长型表现在数量性状间的关系为非线性函数形式[14-15]。扁秆藨草种群生殖株地上部茎叶生物量与株高和株生物量均呈极显著幂函数正相关;花序生物量与株高呈幂函数增加的趋势,与株生物量呈极显著的线性正相关,反映了扁秆藨草种群不同高度和不同生物量生殖株的养分积累及其向营养器官和生殖器官的分配是协调有序的,但是所遵循的规律和速率却不尽相同。扁秆藨草种群生殖株同时具有同速和异速两种不同过程的表型可塑性调节。
3.3株高与株生物量对生殖分配的影响 生殖分配是指植物一年所同化的能量中用于生殖的比例,实际上指总能量供给生殖器官的比例,是植物对有性生殖投入的重要指标,也是植物适应环境的策略[16]。植物在其生命的过程中,必须把它们的一部分能量用于生殖,不同类型的植物常常采取不同的生殖对策。处于不同生长阶段的某些植物把较多的能量用于营养结构的生长,而分配给生殖部分的能量较少,这些植物的生殖能力比较低[14]。扁秆藨草种群生殖分配Ⅰ与生殖株株高呈极显著的指数函数负相关关系,与株生物量呈显著的幂函数负相关关系;生殖分配Ⅱ与株高呈极显著的幂函数负相关关系,与株生物量呈幂函数下降的趋势。表明扁秆藨草种群生殖株高度和株生物量的增加,是以减少生殖分配为代价,将更多的能量分配给茎叶的营养生长,以提高其现实竞争力,更有利于植株个体的生长和存活。通过对生长与生殖的权衡,实现其对资源的最佳利用,体现了在不同生长状况下,生殖株对有限资源所采取的不同分配策略[17-19]。
[1]杨允菲,张宝田.松嫩平原草甸生境甘草种群生殖构件表型可塑性及变化规律[J].草业学报,2006,15(2):14-20.
[2]Pigliucci M.Phenotypic Plasticity, Beyond Nature and Nurture[M].Baltimore:The Johns Hopkins University Press,2001:11-23.
[3]Bradshaw A D,Hardwick K.Evolution and stress genotypic and phenotypic components[J].Biological Journal of the Linneon Society,1989,37:137-155.
[4]Schlichting C D,Pigliucci M.Phenotypic Evolution:A Reaction Norm Perspective[M].Sunderland, Massachusetts:Sinauer Press,1998:9-15.
[5]干友民,任婷,陈燕,等.西南地区野生马蹄金无性繁殖特性研究[J].草业科学,2009,26(8):163-171.
[6]Wang L Y.Effect ofScirpusplaniculmison the remediation of heavy metals of municipal sludge[A].In:2009 3rd International Conference on Bioinformatics and Biomedical Engineering[C].Beijing:ICBBE,2009:1-3.
[7]郑志华.莫莫格湿地扁秆藨草繁育特性与生态特性研究[D].北京:北京林业大学,2008:10-11.
[8]惠淑荣,李响,张志翔,等.土壤含盐量对扁秆藨草生长的影响[J].辽宁林业科技,2009(1):25-28.
[9]康学耕,富力,唐恩全,等.松辽生态区扁秆藨草无性繁殖规律的数量研究[J].植物学报,1993,35(6):466-471.
[10]韩逢春,李尧.黑龙江省稻田扁秆藨草和日本藨草的发生危害及其防治对策[J].黑龙江农业科学,1993(4):31-35.
[11]杨允菲,张宝田.松嫩平原贝加尔针茅无性系构件的结构及生长规律[J].应用生态学报,2005,16(12):2311-2315.
[12]张丽辉,李海燕,杨允菲.松嫩平原返青期2种碱茅种群构件的数量特征研究[J].安徽农业科学,2006,34(21):5527-5528.
[13]杨允菲,李建东.东北草原羊草种群单穗数量性状的生态可塑性[J].生态学报,2001,21(5):752-758.
[14]张春华.松嫩平原披碱草种群生殖分蘖株生长及分配研究[J].草原与草坪,2007(5):42-45.
[15]徐飞.不同生境下麻栎和刺槐幼苗整株及叶性状的表型可塑性研究[D].济南:山东大学,2009:20-22.
[16]李小钌,刘晓东,邢军会,等.毛果苔草生殖分蘖株生长及生殖分配的表型可塑性调节[J].东北林业大学学报,2009,37(9):54-56.
[17]张春华,杨允菲.松嫩平原寸草苔种群生殖分株的种子生产与生殖分配策略[J].草业学报,2001,10(2):7-13.
[18]陈学林,梁艳,齐威,等.一年生龙胆属植物的繁殖分配及其花大小、数量的权衡关系研究[J].草业学报,2009,18(5):58-66.
[19]宋智芳,安沙舟,孙宗玖.刈割和放牧条件下伊犁绢蒿生物量分配特点[J].草业科学,2009,26(12):118-123.
