啮齿动物种群数量与年降水量的灰色关联度分析
2011-03-31包俊江武晓东付和平张福顺李颖班晓瑞
包俊江,武晓东,付和平,张福顺,李颖,班晓瑞
(1.内蒙古农业大学生态环境学院,内蒙古 呼和浩特 010019;2.内蒙古工业大学能源与动力工程学院,内蒙古 呼和浩特 010062;3.内蒙古鄂尔多斯市气象局,内蒙古 鄂尔多斯 017000;4.内蒙古自治区烟草质量监督检测站,内蒙古 呼和浩特 010010)
啮齿动物种群数量和气候因子之间的关系一直是生态学家们关心的问题,尤其以北美、南美和澳大利亚地区由El Nio现象引起的降水量变化对啮齿动物的影响研究较多[1-2],但所得结论并不一致。有研究认为啮齿动物数量不受降水变化的影响[3],而有研究表明啮齿动物的数量暴发与降水高峰是相关的[4]。这些研究往往仅对单一物种进行分析[5-7],缺少对多个物种的综合研究。
阿拉善荒漠是内蒙古西部典型的温性荒漠,啮齿动物作为荒漠草地中重要的功能群,其数量大小和生态功能对生态系统的平衡和稳定具有重要意义。降水稀少而且极不均匀是荒漠生态系统的主要特点[8],对啮齿动物种群数量大小存在直接或间接作用。本研究地区天然草地存在围栏禁牧、适度轮牧和过度放牧3种利用方式,但划区轮牧是该地区的主要利用方式,轮牧草地面积较大。因此,本研究对轮牧区中啮齿动物和降水量的相关性进行分析。降水量与啮齿动物种群数量之间的关系是非线性的[9],运用线性分析方法很难揭示事物的内在联系。而灰色关联度分析主要是通过对灰色系统中不同事物间的相关分析,根据因素之间发展趋势的相似或相异程度,衡量因素间关联程度的一种非线性方法[10]。本研究采用该方法计算群落中主要鼠种种群数量与年降水量的关联度大小,分析各鼠种对降水量变化的反应,为揭示啮齿动物群落格局变动规律提供依据。
1 研究区概况与研究方法
1.1 研究区自然概况研究区位于内蒙古阿拉善左旗南部的嘉尔格勒赛汗镇,地理坐标为104°10′~105°30′E,37°24′~38°25′N,地处腾格里沙漠东缘。该地区的草地类型是典型的温性荒漠,植被稀疏,结构单调,覆盖度低,一般为1%~20%。植物种类贫乏,主要以旱生、超旱生和盐生的灌木和小半灌木为主。建群植物以藜科(Chenopodiaceae)、菊科(Compositae)和蒺藜科(Zygophyllaceae)为主,其次为蔷薇科(Rosaceae)、柽柳科(Tamaricaceae)。地形起伏不平,丘陵、沙丘与平滩相间。气候为典型的高原大陆性气候,冬季严寒、干燥,夏季酷热,昼夜温差大,极端最低气温-36℃,最高气温42℃,年平均气温8.3℃,无霜期156 d。年降水量45~215 mm,且降水极不均匀,主要集中在7-8月。年蒸发量3 000~4 700 mm。土壤为棕漠土,淋溶作用微弱,土质松散、瘠薄,表土有机质含量1.0%~1.5%,含有较多的可溶性盐。
1.2 研究方法2000年,研究区域开始实行划区轮牧制度,2002年选择500 hm2的天然草地作为试验区,划分为 4个轮牧小区,放牧 300只羊,各小区每月放牧一次。为了保证结论的准确性,研究期间,没有更换试验区,并且放牧制度始终保持一致。
2002-2008年每年的4、7、10月的上旬采用铗日法对啮齿动物种群数量进行调查。每次取样时,在每个轮牧小区随机选择1个铗捕样方,样方面积10 hm2。每个样方布设5条铗线,每铗线100铗,铗距5 m,铗线距50 m,共计500铗日。以啮齿动物的捕获率代表种群的相对密度,用各年度4、7、10月的平均捕获率表示该年度的捕获率。2002-2008年试验区的降水量数据来源于内蒙古阿拉善盟孪井滩气象站。
1.3 分析方法采用灰色系统分析方法中的灰色关联分析。其原理是依据空间理论为数学基础,按照规范性、偶对称性、整体性和接近性的原则,对信息部分确定和部分不确定的灰色系统的发展势态进行定量描述和比较,以确定参考数列和若干比较数列之间的灰色关系。具体分析步骤如下。
将降水量与各鼠种的种群密度视为一个整体,即灰色关联系统。并将降水量设定为参考数列X0,记作:X0={X0(1),X0(2),…,X0(n)},捕获鼠种的种群密度分别设定为比较数列X1~Xi,记作:Xi={Xi(1),Xi(2),…,Xi(n)},(i=1,2,…,n),n为鼠种数。对各数列进行均值化无量纲处理,即将同一个数列的所有数据均除以该序列均值得到一个新数列,计算公式表示为:
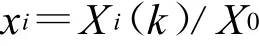
求出参考数列X0与比较数列Xi各对应点的绝对差值,即 Δi(k),

关联系数的计算:

关联度(ri)的计算:

式中,ri为比较数列与参考数列的关联度,是具体反映X0与Xi数列之间关联性的度量[7]。
2 结果与分析
2.1 啮齿动物群落的物种组成2002-2008年共布放 42 000铗日,捕鼠2 861只(表1),分属4科11种。它们分别为跳鼠科(Dipodidae):三趾跳鼠(Dipussagitta)、五趾跳鼠(Allactaga sibirica)、蒙古羽尾跳鼠(Stylodipus andrewsi)、三趾心颅跳鼠(Salpingotus kozlovi);仓鼠科(Cricetidae):小毛足鼠(Phodopus roborovskii)、黑线仓鼠(Cricetulusbarabensis)、短耳仓鼠(Allocricetuluseversmanni)、子午沙鼠(Meriones meridianus)、长爪沙鼠(M.unguiculatus);松鼠科(Sciuridae):阿拉善黄鼠(Spermophilus alaschanicus);鼠科(Muridae):小家鼠(Musmusculus)。子午沙鼠、三趾跳鼠、小毛足鼠、五趾跳鼠、阿拉善黄鼠和黑线仓鼠为群落中的主要鼠种(表1),各年中的捕获数量较大,所以本研究仅对以上鼠种进行了种群数量和降水量的灰色关联度分析。其中子午沙鼠和小毛足鼠为群落的优势种,7年中的捕获量明显高于其他鼠种。

表1 2002-2008年啮齿动物群落的物种组成 只
2.2 啮齿动物种群数量与降水量的年动态2002-2008年群落中总的啮齿动物数量与年降水量的变动规律如图1所示,啮齿动物种群数量和降水量的年动态波动均较大,变动趋势基本一致。啮齿动物的种群数量在2003年达到一个数量高峰,随后降低,2006达到最低点,总捕获率仅为1.18%,随后逐渐上升,2008年达到另一个数量高峰,总捕获率高达21.02%。降水量的第1个高峰也出现2003年,2005年降至最低,仅为63.3mm,随后上升,2007年达到另一高峰,为288 mm。2002-2008年啮齿动物的捕获率与年降水量的变动趋势是一致的,仅2008年有差异,2008年降水量较2007年降低,而啮齿动物的捕获率明显高于2007年。但是经Spearman相关分析发现,2002-2008年总的啮齿动物数量与年降水量呈显著正相关(r=0.390 5,P=0.032 4)。
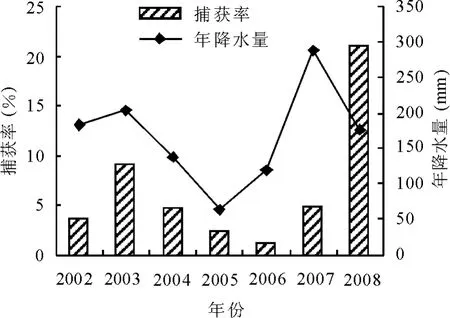
图1 啮齿动物捕获率和降水量的年动态
2.3 啮齿动物种群数量与年降水量的灰色关联度分析
2.3.1数据无量纲化处理 将年降水量设定为参考数列X0,五趾跳鼠、三趾跳鼠、阿拉善黄鼠、小毛足鼠、子午沙鼠和黑线仓鼠的年捕获率分别设定为比较数列X1~X6。由于啮齿动物捕获率与年降水量的计量单位不同,且数值差异较大,需对各原始数据进行均值化变换进行标准化无量纲处理(表2)。

2.3.3关联系数和关联度 根据关联系数计算公式(2),可以求出各鼠种捕获率与该地区年降水量的关联系数(表4)。各鼠种捕获率与年降水量的关联系数在0.541 8~1.000 0。
将表4中的各关联系数代入公式(3),计算出各鼠种的年捕获率与年降水量的关联度(表5)。关联度的大小反映了年降水量对不同鼠种的作用大小,关联度越大,则表示年降水量对该鼠种的影响越大。年降水量与群落中主要鼠种的关联度大小顺序为:三趾跳鼠>五趾跳鼠>小毛足鼠>阿拉善黄鼠>子午沙鼠>黑线仓鼠(表5)。因此,可以看出群落中跳鼠对降水量的反应较明显,三趾跳鼠和五趾跳鼠的灰色关联系数分别为0.855 6和0.818 9;子午沙鼠和黑线仓鼠对降水量的反应较弱,关联系数仅为0.706 7和0.706 4。
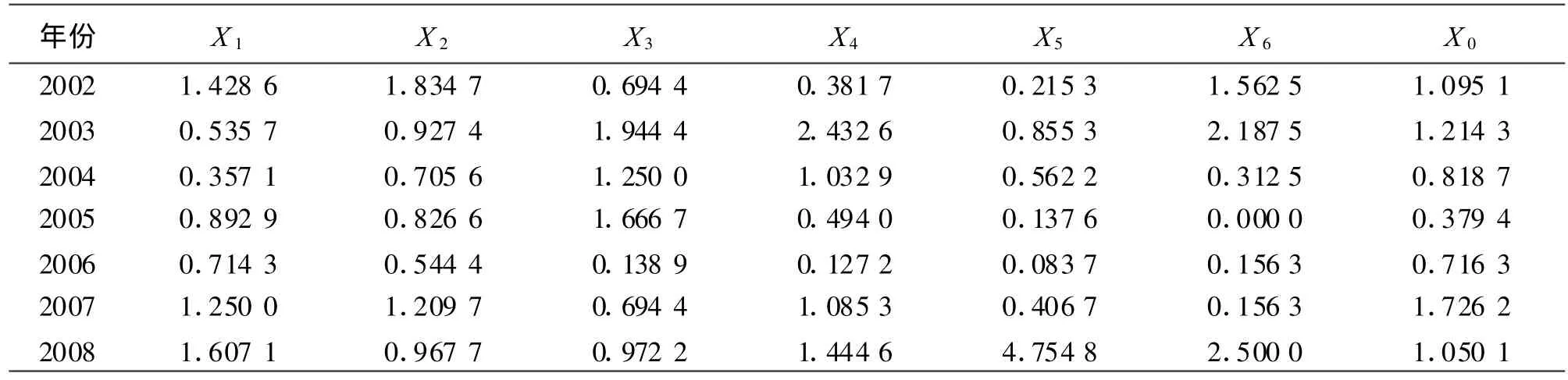
表2 数据标准化处理
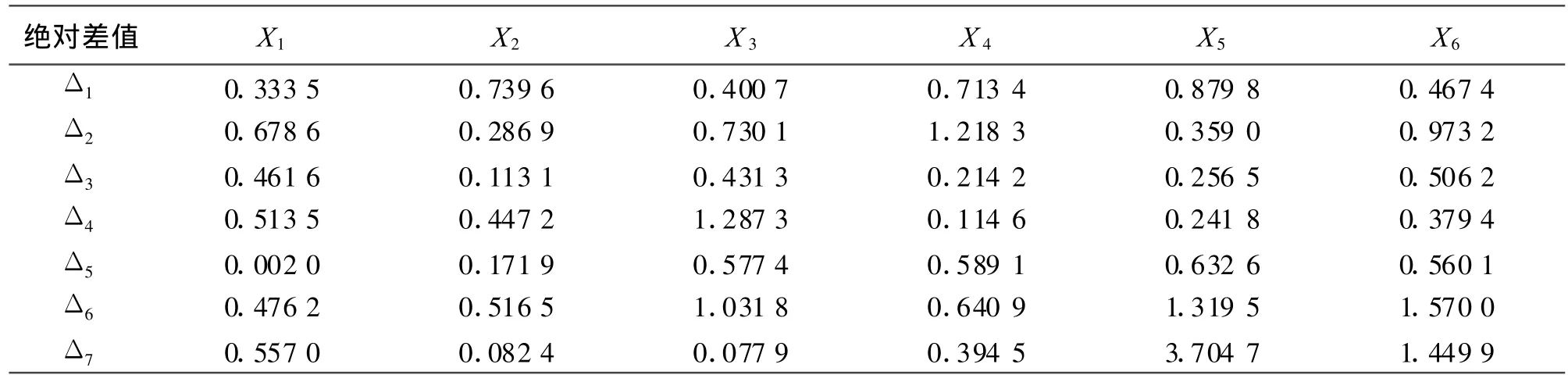
表3 年降水量与各鼠种捕获率的绝对差值

表4 年降水量与各鼠种捕获率的关联系数

表5 群落中主要鼠种与全年降水量的关联度
3 讨论与结论
灰色系统理论是由我国学者邓聚龙于1982年提出的[11]。近年来,该理论在工业、农业、生态和生物等许多领域得到广泛应用[12-19],但国内外关于该方法在啮齿动物研究上的应用鲜见报道。灰色关联度分析是该理论中的重要分析方法,它的优点在于,对样本量的多少和有无规律都同样适用,不要求样本服从某个典型概率分布,不要求各因素数据与系统特征数据之间呈线性关系,弥补了采用数理统计方法作系统分析所导致的缺憾。啮齿动物种群数量与降水量之间的关系是复杂的,降水通过影响啮齿动物的繁殖和食物资源等,影响其种群的大小。它们之间的关系是模糊的、灰色的,符合灰色理论中的灰色系统,因此灰色关联度分析是揭示啮齿动物与环境因子之间关系的一种有效方法。该方法的不足之处在于,关联度分析法中只有正关联度,无负关联度,其范围为0~1。而在数理统计学中的相关系数,有正、负和零,这符合生物界各种因素的相互影响有正、负相关性的客观事实[13]。
本研究7年中在研究区共捕获11个鼠种,选取了其中6个主要鼠种进行了分析。因为其他鼠种在各年度的捕获数量较低或仅在个别年份中被捕获,加以分析会影响关联分析的准确性。例如,2008年长爪沙鼠的捕获数量较大,为179只,而其他年份基本为0。长爪沙鼠在该地区主要生存在开垦农田生境中,2008年为长爪沙鼠的高峰年,可能由于其发生由农田向天然草地的扩散现象,造成该年天然草地中长爪沙鼠的数量明显高于其他年份。因此,长爪沙鼠不是轮牧区中的主要组成物种,本研究中没有对其进行灰色关联度分析。
国内外关于啮齿动物与降水量关系进行了大量研究,Ernest等[20]研究表明,美国Sevilleta地区的啮齿动物种群大小和降水量是相关的。T riggs[5]发现小家鼠的种群数量与降水量变化有关。赵天飙等[6]研究表明大沙鼠(Rhombomys opimus)的种群数量与降水量显著相关,其种群密度和当年降水量的变化基本一致 。本研究与以上研究结果相似,啮齿动物种群数量与年降水量呈显著正相关,说明降水量是影响啮齿动物数量变动的重要因素之一。但是并不能忽视其他生态因子的作用,因为影响生物的因子是多方面的,多个因子共同作用决定生物的种群数量。因此,本研究中啮齿动物数量的大小是由降水与其他生态因子共同作用决定的,有关各因子的作用大小还有待进一步研究。
近年来,生态学家们提出了资源脉冲理论[21],资源脉冲作用广泛存在于生态系统中[22]。资源脉冲理论对于理解荒漠生态系统中啮齿动物与降水量之间的关系是十分重要的。降水在荒漠生态系统中作为重要的脉冲资源,首先对植物种子的萌发、生长和繁殖产生作用,而植物又作为脉冲资源促进了啮齿动物的生存和繁殖[20],决定了啮齿动物的数量大小。因此,资源脉冲理论可以用来解释本研究中啮齿动物数量与年降水量呈显著正相关的结果。本研究中2002-2007年,啮齿动物数量与年降水量可以表现出正相关的变动趋势,而2008年降水量较2007年降低,啮齿动物的捕获率却明显高于2007年。笔者认为滞后效应可能是造成该现象的原因,Letnic等[2]研究表明,在澳大利亚中部Simpson荒漠地区,啮齿动物数量对于El Nio现象引起的降水变化表现出滞后效应。Ernest等[20]也发现啮齿动物数量变化至少滞后于降水一个季节。因此,本研究中由于2007年的降水量较高,啮齿动物数量对其反应表现出滞后效应,导致2008年啮齿动物达到较高的数量。
不同啮齿动物正常代谢过程中所需的水分平衡条件不同[23],导致其对降水量的反应也可能不同[24]。例如,内蒙古荒漠草原地区长爪沙鼠的种群密度与降水量是负相关的,而达乌尔鼠兔(Ochotona dauurica)种群增长率与降水量正相关[25-26]。本研究表明,群落中总的啮齿动物种群数量与年降水量呈显著正相关,但是各鼠种与年降水量的相关性并不一致。群落中主要鼠种与年降水量的关联度顺序为:三趾跳鼠>五趾跳鼠>小毛足鼠>阿拉善黄鼠>子午沙鼠>黑线仓鼠。子午沙鼠和小毛足鼠为群落中的优势鼠种,它们与降水量的关系相对较弱,而跳鼠的种群大小对降水量的反应最明显。可见,啮齿动物对降水量变化的反应与其在群落中生态位的高低无关,可能与物种在长期进化中形成的生态适应机制有关,具体原因尚需进一步探讨。
[1]Jaksic F M.Ecological effects of El Nio in terrestrial ecosystems of western South America[J].Ecography,2001,24(3):241-250.
[2]Letnic M,Tamayo B,Dickman C R.The responses of mammals to La Nia(El Nio southern oscillation)-associated rainfall,predation,and wildfire in central Australia[J].Journal ofMammalogy,2005,86(4):689-703.
[3]Meserve P L,Yunger J A,Gutié rrez J R,et al.Heterogeneous responses of small mammals to an El Nio southern oscillation event in northcentral semiarid Chile and the importance of the ecological scale[J].Journal of Mammalogy,1995,76(2):580-595.
[4]Jaksic F M,Lima M.M yths and facts on ratadas bamboo blooms,rainfall peaks and rodent outbreaks in South America[J].Austral Ecology,2003,28(3):237-251.
[5]Triggs G S.The population ecology of house mice(Musdomesticus)on the lsle of May,Scotland[J].Journal of Zoology,1991,225(3):449-468.
[6]赵天飙,杨持,周立志,等.大沙鼠种群密度与降水量的关系[J].兽类学报,2007,27(2):195-199.
[7]姜运良,卢浩泉,李玉春,等.山东阳谷县黑线仓鼠种群数量预测预报[J].兽类学报,1994,14(3):195-201.
[8]Walsberg G E.Small mammals in hot deserts:some generalizations revisited[J].BioScience,2000,50(2):109-119.
[9]Lima M,M arquet P A,Jaksic F M.El Nio events,precipitation patterns,and rodent outbreaks are statistically associated in semiarid Chile[J].Ecography,1999,22(2):213-218.
[10]郭娜,郭科,吴金炉,等.灰色关联度分析法在土地评价中的应用[J].成都理工大学学报(自然科学版),2007,34(6):626-629.
[11]邓聚龙.灰色系统基本方法[M].武汉:华中理工大学出版社,1992.
[12]刘玉华,史纪安,贾志宽.气候因子与苜蓿草产量的灰色关联度分析[J].草业科学,2009,26(8):101-106.
[13]杨诗兴.试论灰色系统关联度分析法在生物科学研究中应用的合适性[J].家畜生态,2003,24(4):10-13.
[14]杨曌,张新全,李向林,等.应用灰色关联度综合评价17个不同秋眠级苜蓿的生产性能[J].草业学报,2009,18(5):67-72.
[15]苏才旦,周雷.共和盆地温性草原物候期草地生产力与气候因素的灰色关联度分析[J].草业科学,2009,26(5):38-43.
[16]刘玉成,杜道林,岳泉.绪云山森林次生演替中优势种群的特性与生态因子的关联度分析[J].植物生态学报,1994,18(3):283-289.
[17]蔡丽艳,石凤翎,高翠萍,等.苜蓿雄性不育系杂交制种产量灰色关联度分析[J].种子,2010,29(3):19-23.
[18]李云乐,乔玉辉,孙振钧,等.不同土壤培肥措施下农田有机物分解的生态过程[J].生态学报,2006,26(6):1933-1939.
[19]马姜明,李昆,张昌顺.元谋干热河谷苏门答腊金合欢、新银合欢人工林天然更新初步研究[J].应用生态学报,2006,17(8):1365-1369.
[20]Ernest S K M,Brown J H,Parmenter R R.Rodent,plant,and precipitation:spatial and temporal dynamics of consumers and resource[J].Oikos,2000,88(3):470-482.
[21]Holt R D.Theoretical perspectives on resource pulses[J].Ecology,2008,89(3):671-681.
[22]Orland M C,Kelt D A.Responses of a heteromyid rodent community to large-and small-scale resource pulses:diversity,abundance,and home-range dynamics[J].Journal of Mammalogy,2008,88(5):1280-1287.
[23]Tirado C,Corté s A,Bozinovic F.Water balance in two South American Phyllotis desert rodents,P.xanthopygus rupestrisandP.darwini darwini[J].Journal of Arid Environments,2008,72(5):664-670.
[24]Reed A W,Slade N A.Environmental correlates of survival and reproduction in old-field rodents[J].Journal of Mammalogy,2009,90(3):680-685.
[25]Wang G M,Zhong W Q.Mongolian gerbils and Daurian pikas responded differently to changes in precipitation in the Inner Mongolian grasslands[J].Journal of Arid Environments,2006,66(4):648-656.
[26]Liu W,Wan X,Zhong W Q.Population dynamics of the Mongolian gerbils:Seasonal patterns and interactions among density,reproduction and climate[J].Journal of Arid Environments,2007,68(3):389-397.
