谷氨酸棒杆菌关键酶活性与苏氨酸高产的关系
2011-03-10张君胜杨晓志
张君胜,王 扬,杨晓志,张 力
(江苏畜牧兽医职业技术学院动物科技学院,江苏 泰州 225300)
苏氨酸(Threonine)是仅次于蛋氨酸、赖氨酸和色氨酸的第4种限制性氨基酸,在医药和畜牧业中有非常重要的作用[1]。近年来随着人们对于苏氨酸需求量的日益增加,苏氨酸生产的研究也日益成为了热点。目前,发酵法以其生产成本低、资源节约、环境污染小等优点逐渐成为工业化生产L-苏氨酸的主要方式。
在谷氨酸棒杆菌代谢过程中,葡萄糖经糖酵解途径生成磷酸烯醇式丙酮酸,磷酸烯醇式丙酮酸经二氧化碳固定反应生成四碳二羧酸,经氨基化反应生成天冬氨酸;天冬氨酸在天冬氨酸激酶催化作用下,生成天冬氨酸半醛;天冬氨酸半醛在高丝氨酸脱氢酶的催化下生成高丝氨酸;高丝氨酸在高丝氨酸激酶的催化下生成苏氨酸[2-3]。在微生物发酵合成L-苏氨酸的整个代谢途径中,高丝氨酸脱氢酶和天冬氨酸激酶控制着整个发酵过程中碳源和氮源的走向,它们是决定能否获得高产L-苏氨酸的关键酶[4]。笔者采用诱变选育的一系列谷氨酸棒状杆菌菌株,测定其高丝氨酸脱氢酶和天冬氨酸激酶两个关键酶的活性,研究了谷氨酸棒杆菌两个关键酶与发酵产L-苏氨酸的关系,并对比了诱变处理前后酶活的大小,旨在从更深入的代谢途径研究L-苏氨酸产量提高的原因,为L-苏氨酸的育种工作提供研究依据。
1 材料与方法
1.1 材 料
1.1.1 菌 种 谷氨酸棒状杆菌(Cornebacterium glutamicum):WS4006、WS4006-1(L-Met-)、D-6(LMet-+L-Lys-)、K-11(L-Met-+L-Lys-+AHVr)、Y-1(L-Met-+L-Lys-+AHVr+Ile-),全部由笔者实验室诱变获得[5]。
1.1.2 试 剂 L-天冬氨酸、5-三磷酸腺苷二钠盐(ATP)、异丙基硫代-β-D-半乳糖苷、S-2-氨基乙基半胱氨酸、β-巯基乙醇、三羟甲基氨基甲烷、氯代三苯基四氮唑均为上海生工产品;2,3,5-氯化三苯基四唑(即TTC)为Sigma公司产品;其他试剂均为国产分析纯。
1.1.3 培养基 培养基的配制参照文献[5]。
1.2 方 法
1.2.1 高丝氨酸脱氢酶活性的测定方法 高丝氨酸脱氢酶活性测定参照文献[6]略有改动,通过测定人造受氢体为2,3,5-氯化三苯基四唑(即TTC)的脱氢酶活性来反映高丝氨酸脱氢酶活性。将谷氨酸棒状杆菌在肉汤培养基中,30℃,180 r/min培养9 h,使其全部达到对数生长期,此时其酶活性最高。将培养好的新鲜菌液,5 000 r/min离心4min,弃去上清液,加蒸馏水洗涤,摇匀,4 000 r/min离心5 min,弃去上清液(反复3次)。取等体积蒸馏水将菌体配成悬液待用。取若干25mL比色管,分别加入7.5mL Tris-HCl缓冲液、2.5mL 0.4%的TTC溶液、2.5mL 0.36%的Na2CO3溶液和2.5mL纯水,并混合均匀。将比色管置于37℃恒温水浴锅中振荡5 min。加入5mL粗酶液以0.5mL甲醛作为空白对照。37℃恒温水浴锅培养一段时间后,加入人造受氢体TTC,TTC在细胞呼吸过程中接受氢以后,其还原产物三苯基甲膳(即TF)以红色结晶存在于细胞内,开始显色,记下显色时间。当培养达到一定时间后(一般2 h),加入甲醛作为终止剂,混合均匀,即可终止反应。用5mL 80%丙酮萃取TF,在波长485 nm处测出吸光度值。吸光度的大小就反映了高丝氨酸脱氢酶活性的大小。把1 h产生1μg TF的量作为一个酶活力单位。根据所测得的吸光度值,查标准曲线找TTC-脱氢酶活性,就是高丝氨酸脱氢酶的活性。
1.2.2 天冬氨酸激酶(AKI)活性的测定 天冬氨酸激酶(AKI)的活性测定参照文献[7]略有改动。将活化后的谷氨酸棒状杆菌于30℃摇床培养9 h,使其达到对数生长期,然后加入ITPG(终浓度1 mmol/mL)。4 h后收集菌体,6 000 r/min 离心 10min,弃上清液,把菌体悬浮于0.02 mol/L,pH 7.5,含0.03 mol/Lβ-巯基乙醇的冷冻磷酸钾缓冲液中,超声破碎后的悬浊液15 000 r/min离心30min,取上清液作为粗酶液。取粗酶液50μL加入到1mL反应液(底物为L-天冬氨酸10 mmol/L,Tris-HCl缓冲液(pH 8.1)94 mmol/L,ATP 10.4 mmol/L,MgSO41.6 mmol/L,β-巯基乙醇 10 mmol/L,NH4OH 800 mmol/L,KCl 800mmol/L)中。反应体系混合均匀后,在30℃恒温水浴锅中反应30 min后,加入0.5 mL 12%三氯乙酸中止反应。加入0.5mL 3mml/LHCl和 0.5mL FeCl3·6H2O(溶于 0.1mmo1/LHCl中)。混合物400 r/min离心5 min去沉淀,上清液在600 nm波长处测光密度,以反应液中不加L-天冬氨酸为对照。AKI的测活由在600 nm下反应液的吸光度的大小决定。OD600即为天冬氨酸异羟酸离子的光学密度,酶活则表示为:1 000×OD600值。
2 结果与分析
2.1 TTC标准曲线的制作
取8支20mL的带有塞子的试管,在试管中依次加入 Tirs-HCl 2 mL,蒸馏水2 mL,TTC系列溶液2mL,对照管中不加TTC,然后再各试管中加入10%Na2S 1mL,充分震荡显色后,加入甲醛5 mL,萃取3 min,取上层有机溶液在分光光度计485 nm处比色测定吸光度。所绘制标准曲线如图1所示。
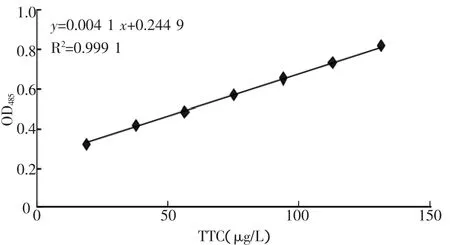
图1 TTC脱氢酶活性标准曲线图
由图1可以看出TTC脱氢酶活性(TTC-DHA)与吸光度A的关系,得回归曲线公式:

y表示吸光度A值,x表示TTC脱氢酶活性,R2表示回归系数。
2.2 基因突变对谷氨酸棒状杆菌高丝氨酸脱氢酶活性的影响
将谷氨酸棒状杆菌 WS4006、WS4006-1、D-6、K-11、Y-1分别经过菌液的培养后,用分光光度计测定其吸光度A值,每个测定做3个重复,其吸光度A值如表1所示。

表1 各菌株高丝氨酸脱氢酶的活性
从表1可以看出,选育出的目的菌株随着遗传特性的增多,其吸光度A值也逐渐增大。将这些数据代入回归方程1可以得到各个目的菌株的TTC浓度。原始菌株WS4006脱氢酶的活性很低,0.124 9 U,随着缺陷型菌株和AHV抗性的选育,其脱氢酶的活性也随之增高,从WS4006、WS4006-1、D-6、K-11 到 Y-1,高丝氨酸脱氢酶的活 性 由 0.124 9、0.179 3、0.220 0、0.395 6 U 到0.416 0 U,分别是出发菌株的 1.44、1.76、3.17、3.33倍,与此相对应L-苏氨酸产量由0.31 g/L提高到1.04、1.21、2.43、4.35 g/L,分别是出发菌株的 3.35、3.90、7.84、14.03倍。由此可见,酶活性提高相对应L-苏氨酸产量也提高。
2.3 基因突变对谷氨酸棒状杆菌天冬氨酸激酶活性的影响
谷氨酸棒状杆菌的目的菌株,经过细胞破碎后提取粗酶液,模拟底物和反应体系后,在分光光度计600 nm处测定光密度。具体结果见表2。
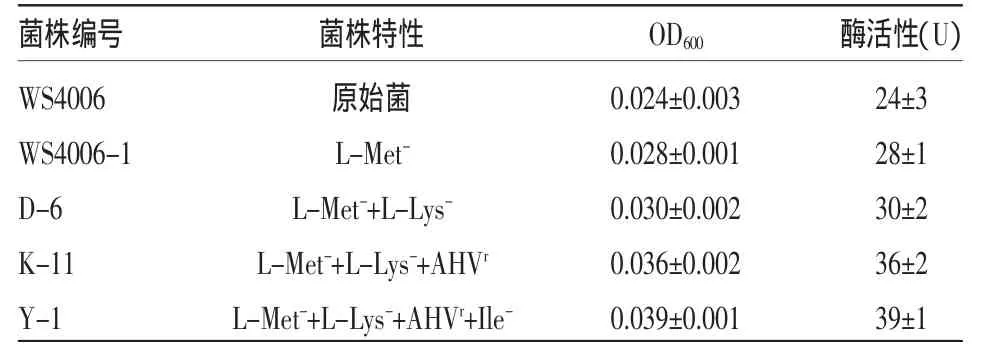
表2 各菌株天冬氨酸激酶的活性
原始菌种天冬氨酸激酶的活性只有23.6 U,随着菌种选育工作的进行,天冬氨酸激酶的活性也逐渐提高,从 WS4006、D-6、K-11 到菌株 Y-1,酶活已经由24 U达到39 U,天冬氨酸激酶的活性分别是出发的原始菌株的 1.17、1.25、1.5、1.65 倍。其中,抗性菌株K-11天冬氨酸激酶活性增幅最大,主要是由于解除了苏氨酸对天冬氨酸激酶的反馈抑制作用。与L-苏氨酸合成相对应,可见天冬氨酸激酶活性提高有利于L-苏氨酸的合成。
3 讨 论
本试验中蛋氨酸缺陷型菌株WS4006-1的选出,解除了蛋氨酸对高丝氨酸脱氢酶的反馈阻遏作用,使高丝氨酸脱氢酶的活性有了一定的提高,但提高不明显。沈琼等[7]在研究苏氨酸操纵子的克隆表达与天冬氨酸激酶的测活时,用分子方法克隆得到thr操纵子,将结构基因装载于pET-lla质粒上的T7启动子下游,得到了表达质粒pTHlla-08,经测定天冬氨酸激酶的活性为含载体质粒pET-lla的对照菌的100倍,但L-苏氨酸产量并没有大量提高[8]。这与本试验结论,天冬氨酸激酶活性提高不明显有很大出入,但L-苏氨酸产量提高不明显结果相似。另外李敬坡等[9]研究发现赖氨酸和甲硫氨酸对天冬氨酸激酶活性没有抑制作用,当苏氨酸浓度大于0.5mmol/L时,对其活性有明显抑制作用,并且抑制作用与苏氨酸呈现浓度依赖性。张雪等[10]经5 L发酵罐发酵产酸实验,携带含苏氨酸操纵子质粒的W3110菌株L2苏氨酸产量为2.590 g/L,质粒上thr A解除反馈抑制后,L-苏氨酸的产量增加到9.223 g/L。此两人研究结果与本研究相近,由此可见,苏氨酸反馈抑制不解除的情况下,提高天冬氨酸激酶活性和L-苏氨酸产量几乎是不可能的。另外解除蛋氨酸时高丝氨酸脱氢酶的反馈抑制作用没有完全解除,因为高丝氨酸脱氢酶除了蛋氨酸的反馈阻遏作用外还受到L-苏氨酸本身的反馈抑制。饶志明等[11]在研究谷氨酸棒状杆菌高丝氨酸脱氢酶编码基因hom的敲除来提高L-赖氨酸产量时,发现赖氨酸产量也没有明显提高,这也与本试验结果类似。可能原因是出发菌为原始菌种,没有经过突变育种改造,影响到了氨基酸产量。
赖氨酸缺陷型D-6(L-Met-+L-Lys-)的选育从一定程度上减弱了L-赖氨酸对于天冬氨酸激酶的反馈抑制作用,使得天冬氨酸激酶活性由原来的23.6 U提高到29.8 U,提高不明显,可能与L-苏氨酸自身对于天冬氨酸激酶的反馈抑制有很大关系。苏氨酸的结构类似物AHV的选育,一方面可以解除苏氨酸自身对于天冬氨酸激酶的反馈抑制,另一方面也解除了对高丝氨酸脱氢酶的反馈抑制,使得这两个酶的活性都有了很大提高。从表2也可以看出,这一步高丝氨酸脱氢酶和天冬氨酸激酶的活性提高最为明显。虽然如此,但L-苏氨酸产量仍然没有大幅提高,可能是抗性育种试验不够彻底,没有能够从根本上解除其自身的反馈抑制作用。
最后选育L-异亮氨酸,是为了不让积累的L-苏氨酸进一步流失,原理上对天冬氨酸激酶和高丝氨酸脱氢酶的活性没有影响,甚至由于分支代谢被切断,有可能间接抑制他们的活性。Hua等[12]曾报道能够过量表达天冬氨酸激酶的重组菌在合成培养基中培养,赖氨酸产量提高,但生长受到影响(减弱);Jetten等[13]的研究结果与其相似。而Koffas等[14]的工作表明重组菌不能在以葡萄糖为碳源的合成培养基上生长。Koffas等认为这可能是由于较高的天冬氨酸激酶活性与回补途径中回补酶的活性不平衡造成的。
谷氨酸棒杆菌是氨基酸发酵生产的主要菌株,也是氨基酸发酵的模式菌株。本研究对谷氨酸棒杆菌的发酵产L-苏氨酸和两个关键酶活性的关系进行了研究,由结果可以看出,这两种关键酶的活性还是有了一定的提高。本研究对利用谷氨酸棒杆菌发酵生产L-苏氨酸菌株的选育具有一定的指导意义。
[1] DebabovV G.The threonine story[J].Adv Biochem Eng Biotechno,2003,79:113-136.
[2]陈殿林.微生物工程技术原理 [M].北京:化学工业出版社,2007.
[3] 张克旭,陈 宁,张 蓓,等.代谢控制发酵[M].北京:中国轻工业出版社,1998.
[4]Thierbach G,Halinoswski J,Bachmann B.Cloning of a DNA fragment from Corynebacterium glutamicum conferring aminoethyl cysteine resistance and feedback resistance to aspartokinase[J].Appl Microbiol Biotechnol,1990,32:443-448.
[5] 王 扬,张 力,李伟星,等.亚硝基胍诱变选育L-苏氨酸高产菌的研究[J].贵州农业科学,2010,38(4):107-109.
[6] 赵 智,刘阳剑,王 宇,等.抗反馈抑制的天冬氨酸激酶基因在钝齿棒杆菌中的表达[J].微生物学报,2005,8(45):4,530-533.
[7] 沈 琼,黄雪峰,吴海珍,等.苏氨酸操纵子的克隆表达及天冬氨酸激酶的测活[J].药物生物技术,2003,10(3):133-136.
[8] 天津轻工业学院.氨基酸工艺学[M].北京:中国轻工业出版社,1983.
[9] 李敬坡,韩宏岩,夏 勇,等.极端嗜热菌Thermus thermophilus HB27中天冬氨酸激酶在大肠杆菌中表达、纯化及酶学性质研究[J].山东农业大学学报(自然科学版),2009,40(4):479-483.
[10]张 雪,闫继爱,于 雷,等.含苏氨酸操纵子重组质粒的构建及其对大肠杆菌L-苏氨酸积累的影响 [J].微生物学报,2009,49(5):592-597.
[11]饶志明,沈 微,张君胜,等.谷氨酸棒杆菌高丝氨酸脱氢酶编码基因 hom 的敲除[J].中国生物工程杂志,2007,27(1):59-63.
[12]Qiang H,Chen Y,Kazuyuki S.Metabolic control analysis for lysine synthesis using Corynebacterium glutamicum and experimental verification[J].Biosci Bioeng,2000,90(2):184-192.
[13]Jetten M.Follettie M.Sinskey A.Effect of different levels of aspartokinase on the lysine production of Corynebacterium lactofermentum[J].Appl Microbiol Biotechnol,1995,41:76-821.
[14]KoffasM A,Jung GY,StephanopoulosG.Engineeringmetabolism and product formation in Corynebacterium glutamicum by coordinated gene overexpression[J].Metab Eng,2003,5(1):32-41.
