AT1受体在脑胆碱能刺激引起的延髓腹外侧部 TH-IR变化中的作用
2011-01-23姚齐颖夏满莉周新妹姜春玲
王 敏,姚齐颖,夏满莉,周新妹,姜春玲
AT1受体在脑胆碱能刺激引起的延髓腹外侧部 TH-IR变化中的作用
王 敏1,姚齐颖2,夏满莉1,周新妹1,姜春玲2
(1.浙江嘉兴学院医学院生理学教研室,浙江嘉兴 314001;2.大连医科大学生理学教研室,辽宁大连 116027)
目的:探讨大鼠侧脑室注射胆碱能激动剂氨甲酰胆碱 (carbachol,CBC)后引起的肾排钠量、肾小球滤过率、肾血浆流量的改变和延髓腹外侧部酪氨酸羟化酶免疫反应活性 (thyrosine hydroxylase-immunoreactivity,TH-IR)的变化,以及氯沙坦 (losartan,Los)阻断脑血管紧张素能 AT1受体后对上述变化的影响。方法:选用 SD雄性大鼠,随机分成 4组:NS+CBC组、Los+CBC组、Los+NS组、NS+NS组,侧脑室注射氨甲酰胆碱 (0.5μg)和 /或氯沙坦 (20μg)后,利用整体实验方法测定肾排钠量、肾小球滤过率、肾血浆流量;采用免疫组织化学方法观察大鼠延髓腹外侧部酪氨酸羟化酶免疫反应活性的变化。结果:与 NS+NS组比,大鼠侧脑室注射氨甲酰胆碱 (0.5μg)后 40 min,肾排钠量显著增加,延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元数目明显增多,免疫染色强度明显增加 (P<0.05);与 NS+CBC组比,氯沙坦预处理后大鼠肾排钠量显著下降,延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元数目明显下降,免疫染色强度明显降低 (P<0.05)。此外,实验还观察到,侧脑室注射氨甲酰胆碱或氯沙坦对肾小球滤过率和肾血浆流量无明显影响 (P>0.05)。结论:大鼠侧脑室给予氨甲酰胆碱后,可显著引起促钠排泄反应,对延髓腹外侧部的儿茶酚胺能神经元有兴奋作用;阻断脑血管紧张素能 AT1受体可下调氨甲酰胆碱诱导的上述变化。
卡巴胆碱 /药理学;受体,血管紧张素,1型 /药理学;延髓 /药物作用;氯沙坦 /药理学
[J Zhejiang Univ(Medical Sci),2011,40(1):71-77.]
延髓腹外侧部是中枢参与循环血量调节的重要部位。血量扩张可引起清醒家兔延髓腹外侧部 c-fos基因表达增加及促钠排泄反应[1],但延髓腹外侧部何种神经元参与血量扩张引起的促钠排泄反应尚未见报道。有文献报道,侧脑室、内侧隔区、蓝斑给予胆碱能刺激或肾上腺素能刺激均能引起促钠排泄反应[2];而α-肾上腺素能受体阻断剂酚妥拉明可阻断上述效应[3],提示脑胆碱能系统和脑儿茶酚胺能系统在促钠排泄反应中有某种联系。我们以往的工作证实,下丘脑内儿茶酚胺能神经元参与脑胆碱能刺激引起的促钠排泄和利尿反应[4]。神经解剖学证实,延髓腹外侧部有丰富的儿茶酚胺能神经元,并接受来自下丘脑、蓝斑、孤束核等与水钠平衡相关脑区的纤维投射[5]。但延髓腹外侧部的儿茶酚胺能神经元是否参与脑胆碱能刺激引起的促钠排泄反应,迄今为止尚未见报道。
另有文献报道,中枢给予血管紧张素Ⅱ可引起升压、饮水及促钠排泄反应[6-7];脑血管紧张素能AT1受体参与大鼠脑室高张刺激引起的肾排钠量、排钾量、尿量增加和平均动脉压升高的效应[6,8],但脑血管紧张素能 AT1受体是否参与侧脑室给予氨甲酰胆碱引起的延髓腹外侧部儿茶酚胺能神经元活性变化的反应目前尚未见报道。酪氨酸羟化酶 (thyrosine hydroxylase,TH)是儿茶酚胺合成的限速酶,可作为儿茶酚胺能神经元活性的标志[5]。本研究应用整体实验和免疫组织化学的方法,观察侧脑室注射胆碱能激动剂氨甲酰胆碱 0.5μg后对肾排钠量、肾小球滤过率 (glomerular filtratior rate,GFR)和肾血浆流量 (renal plasma flow,RPF)的影响,延髓腹外侧部酪氨酸羟化酶免疫反应活性的变化,以及用氯沙坦 (20μg)阻断脑血管紧张素能 AT1受体后对上述变化的影响;探讨中枢儿茶酚胺能神经元和血管紧张素能神经元在脑胆碱能刺激引起的促钠排泄反应中的作用机制及可能的相互关系,为进一步研究肾排钠障碍的中枢机制,揭示其在高血压发生发展中的可能影响提供理论依据。
1 材料和方法
1.1 药品、试剂和仪器 氯沙坦 (losartan,Los)、氨甲酰胆碱 (carbachol,CBC)、TH一抗、二氨基联苯胺、菊粉、对氨基马尿酸为美国Sigma公司产品,其他免疫组化试剂为武汉博士德公司产品或国产分析醇。XY-86型振动切片机 (浙江象山科学精密仪器厂),Easylyte plus Na/K/Cl分析仪 (美国 Medica公司 ),Olympus A.H-B-L万用显微镜 (日本),754型分光光度计 (日本),M IAS-GOODJJ.BMP图像分析仪 (美国)。
1.2 动物分组 36只健康雄性 SD大鼠于一侧脑室埋置导管,1周后将动物随机均分为 4组 (n=9),①NS+NS组:侧脑室先注射 0.85%NaCl 5μl,20 min后再注射 0.85%NaCl 5μl;②NS+CBC组:侧脑室先注射 0.85%NaCl 5 μl,20 min后再注射 0.5μg氨甲酰胆碱 5μl;③Los+CBC组:侧脑室先注射 20μg氯沙坦 5 μl,20 min后再注射 0.5μg氨甲酰胆碱 5μl;④Los+NS组:侧脑室先注射 20μg氯沙坦 5 μl,20 min后再注射 0.85%NaCl 5μl。所有动物均由大连医科大学实验动物中心提供。
1.3 整体实验 实验时各组大鼠麻醉后固定,行气管插管、膀胱插管,分离颈外静脉供输液,分离颈总动脉供取血、监测血压。每 20 min收集尿样本 1次,用 Easylyte p lus Na/K/Cl分析仪测定尿钠浓度。
1.3.1 肾小球滤过率 (GFR)的测定 颈外静脉输注菊粉后,分别于侧脑室注射前 30 min,注射后 30 min和 50 min取血、尿样本。用分光光度计测定肾菊粉的清除率。
1.3.2 肾血浆流量 (RPF)的测定 颈外静脉输注对氨基马尿酸后,分别于侧脑室注射前 30 min,注射后 30 min和 50 min取血、尿样本。用分光光度计测定肾对氨基马尿酸的清除率。
1.4 免疫组织化学实验 侧脑室注药后 40 min,麻醉经心脏灌流,取脑后固定。用振动切片机切取 50μm厚冠状脑片,按 George Paxinos&Charles Watson脑立体定位图谱[8],用中性红染色在低倍镜下挑选前囟后 11.3~14.08 mm(Bregma:-11.3 mm~ -4.08 mm)部位的脑片即为延髓腹外侧部所在部位,进行免疫组织化学实验。①2%Triton孵育 5 min,PBS振荡漂洗;②2%BSA孵育 40 min;③小鼠抗大鼠酪氨酸羟化酶一抗 (1∶2 000)4℃孵育过夜;④PBS漂洗,2%的 BSA孵育 30 min;⑤生物素化的二抗室温孵育 1 h,PBS漂洗;⑥ SABC室温孵育 30 min,PBS漂洗;⑦二氨基联苯胺显色;⑧二甲苯透明,中性树胶封片;⑨光镜下观察酪氨酸羟化酶免疫反应阳性神经元的数目及免疫染色强度并拍片。
1.5 图像分析和统计学分析 选取 NS+NS组、NS+CBC组、Los+CBC组和 Los+NS组大鼠各 6只,用 M IAS-GOODJJ.BMP图像分析系统对延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元的个数和平均光密度进行测定。
1.6 统计学分析 实验数据用 SPSS 13.0进行统计学处理,实验指标用均数±标准差(¯x±s)表示,4组间进行单因素方差分析,并进行SNK两两比较,以 P<0.05为有统计学意义。
2 结 果
2.1 氯沙坦对侧脑室注射氨甲酰胆碱引起的促钠排泄作用的影响 各组间肾排钠量指标经方差分析有显著性统计学差异 (F=10.194,P<0.05)。图 1显示:NS+NS组大鼠注药后120 min排钠量基本稳定于注药前水平。NS+CBC组大鼠注药后排钠量明显增加,40 min达到高峰,与 NS+NS组相比,两组注射前排钠量无统计学差异 (P>0.05);注射后 21~40 min排钠量明显高于 NS+NS组,差异有统计学意义 (P<0.05)。Los+CBC组注射后排钠量也显著增加,但与 NS+CBC组相比,两组注射前排钠量无统计学差异 (P>0.05);注射后 21~40min内排钠量明显低于 NS+CBC组,差异有统计学意义 (P<0.05)。Los+CBC组与 NS+NS组相比,注射前排钠量无统计学差异 (P>0.05);注射后 21~40min内排钠量仍高于 NS+NS组,差异有统计学意义 (P<0.05)。
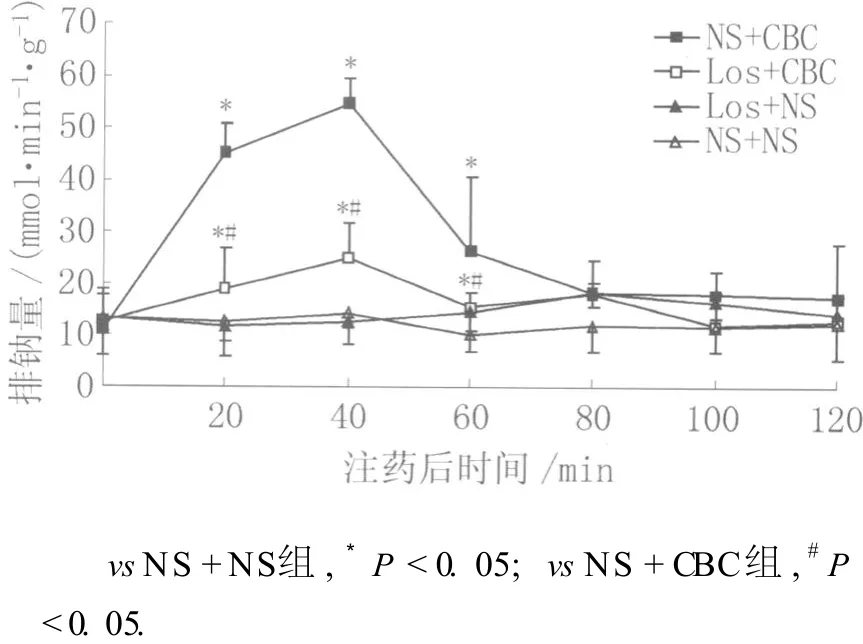
图1 氯沙坦对侧脑室注射氨甲酰胆碱引起的促钠排泄的影响Fig.1 Effect of losartan pretreatment on carbachol-induced natriuresis
2.2 侧脑室注射氨甲酰胆碱和/或氯沙坦对肾小球滤过率和肾血浆流量的影响 各组间肾小球滤过率指标经方差分析无统计学差异 (F=1.157,P>0.05)。表 1显示:NS+NS组大鼠注药后 30min和 50minGFR基本稳定于注药前水平。NS+CBC组大鼠 GFR注药后与注药前相比无统计学差异 (P>0.05),与 NS+NS组相比,注药后 30min和 50minGFR也无统计学差异 (P>0.05)。Los+CBC组大鼠注药后 30min和 50minGFR,与 NS+CBC组及 NS+NS组相比均无统计学差异 (P>0.05)。Los+NS组大鼠注药后 GFR基本稳定于注药前水平,与 NS+NS组相比也无统计学差异 (P>0.05)。
4组肾血浆流量指标经方差分析无统计学差异 (F=1.96,P>0.05,表 2)。而且 RPF的变化规律与 GFR的相似。
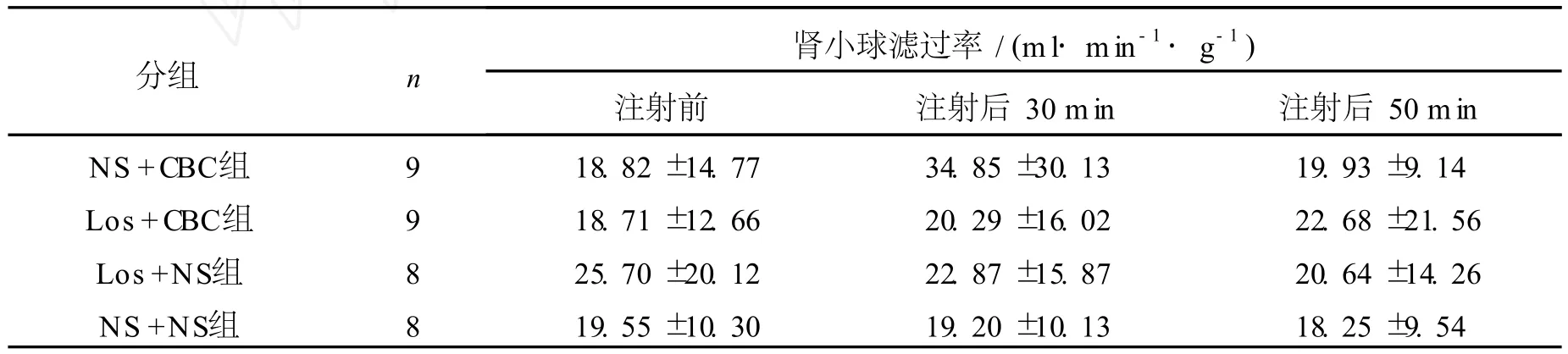
表1 侧脑室注射氨甲酰胆碱及氯沙坦对肾小球滤过率的影响Table1 Effect of carbachol and /or losartan on glomerular filtration rate(¯x±s)

表2 侧脑室注射氨甲酰胆碱及氯沙坦对肾血浆流量的影响Table2 Effect of carbacol and /or losartan on renal plasma flow(¯x±s)
2.3 氯沙坦对侧脑室注射氨甲酰胆碱引起的延髓腹外侧部酪氨酸羟化酶免疫反应活性变化的影响 本研究以延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元的数目和平均光密度作为酪氨酸羟化酶免疫反应强度的评价指标。侧脑室注药后各组间阳性神经元的数目经方差分析有显著性统计学差异 (F=3.924,P=0.024),阳性神经元平均光密度指标经方差分析也有显著性统计学差异 (F=4.596,P=0.013)。NS+NS组大鼠延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元散在分布,细胞形态不清晰,染色较浅 (图 2C)。与 NS+NS组相比,NS+CBC组大鼠延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元密集分布,胞浆深染,细胞轮廓清晰(图 2A),免疫染色强度明显增强 (表 3,P<0.05)。与 NS+CBC组相比,Los+CBC组大鼠延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元数目明显减少,散在分布,胞浆染色较浅,但细胞形态清晰 (图 2B),免疫染色强度明显降低(表 3,P <0.05)。Los+NS组与 NS+NS组相比免疫反应阳性颗粒数目和免疫染色强度无统计学差异 (图 2、表 3,P >0.05)。

图2 氯沙坦对侧脑室注射氨甲酰胆碱引起的延髓腹外侧部酪氨酸羟化酶免疫阳性反应变化的影响Fig.2 Effect of losartan on the changes of TH-IR in RVL induced by ICV injection of carbachol
3 讨 论
研究表明,脑中枢参与肾钠水排出的调节,下丘脑、蓝斑 (locus coeruleus,LC)给予胆碱能刺激可引起动物摄水、平均动脉压升高和肾的钠排泄反应增加[2-3]。本研究结果显示:大鼠侧脑室注射氨甲酰胆碱 (0.1、0.2、0.5μg)也可引起肾排钠增加,并且呈现明显的量效关系。这表明肾的钠排泄直接受到中枢胆碱能系统的调节。注射 0.5μg氨甲酰胆碱后动脉血压迅速升高,20 min后基本恢复,而肾排钠量增加持续 100 min,这表明血压升高不是侧脑室注射氨甲酰胆碱引起持续性促钠排泄反应的主要因素,可能有其他神经体液因素参与这种促钠排泄反应。有研究报道[9],用正常犬血量扩张后的血液,灌流游离肾,尿钠排出量显著增加;而用去头犬血量扩张后的血液,灌流游离肾,则不引起尿钠排出量增加的反应。这提示脑中枢在调控肾促钠排泄反应中是重要的参与者。
本实验还发现,侧脑室注射氨甲酰胆碱和 /或氯沙坦后,大鼠的肾小球滤过率和肾血浆流量均无明显改变,进一步表明,脑胆碱能刺激引起的促钠排泄反应不是通过改变肾血流动力学来实现的,可能是中枢通过神经体液机制作用于肾小管引起的。有研究发现,胆碱能刺激大鼠下丘脑外侧区等核团引起的促钠排泄反应与肾神经活动改变无关,与血压升高、肾小球滤过率和肾血浆流量的改变也无关,可能与某些体液因子有关[10]。Baldissera[10-11]发现,胆碱能刺激第三脑室前腹部在引起肾钠钾水排出增加的同时伴有血浆心房钠尿肽水平的升高,垂体和下丘脑内侧区的心房钠尿肽也有增高;损毁第三脑室前腹部后上述脑区心房钠尿肽水平降低,并且血浆心房钠尿肽含量明显降低。上述研究提示,脑区的心房钠尿肽能神经元参与了脑胆碱能刺激引起的血浆心房钠尿肽升高的反应,脑胆碱能刺激引起的促钠排泄反应与中枢的心房钠尿肽的释放有关。
另研究发现,侧脑室给予肾上腺素可引起与上述相似的促钠排泄反应[3],提示脑胆碱能刺激和脑儿茶酚胺能刺激引起的促钠排泄反应很可能具有或部分具有共同的神经传导通路。本实验观察到,侧脑室注射氨甲酰胆碱后 40 min,延髓腹外侧部酪氨酸羟化酶免疫反应阳性神经元数目明显增多,免疫染色强度明显增强。这表明延髓腹外侧部不仅参与应激、防御时交感紧张性冲动和心血管稳态的调控,而且参与机体水钠代谢的调节。饮食中钠摄入增加可引起延髓腹外侧部心血管反应增强[12],提示延髓腹外侧部通过调节水钠代谢从而影响心血管活动。免疫组织化学证实,血管紧张素能 AT1受体在许多脑区如蓝斑、孤束核、延髓腹外侧部的分布与儿茶酚胺能神经元的分布有显著的重叠性[13],提示脑血管紧张素能神经通路与脑儿茶酚胺能神经通路在功能上可能具有密切联系。本实验还发现:选用脑血管紧张素能 AT1受体拮抗剂氯沙坦预处理后,可明显减弱氨甲酰胆碱诱导的延髓腹外侧部儿茶酚胺能神经元活性的变化,表明内源性的血管紧张素Ⅱ作用于AT1受体后,可能通过或部分通过影响儿茶酚胺能神经元的活性,参与侧脑室注射氨甲酰胆碱引起的促钠排泄反应。本实验结果结合文献报道[14],推测侧脑室注射氨甲酰胆碱后,可能引起内源性血管紧张素Ⅱ的释放,作用于延髓腹外侧部的 AT1受体,引起延髓腹外侧部儿茶酚胺能神经元兴奋,释放儿茶酚胺,后者作为神经递质,通过某种神经液递机制引起肾脏的促钠排泄反应。
近几年的研究都在探讨中枢血管紧张素Ⅱ增强儿茶酚胺能神经元活性的机制,包括促进儿茶酚胺类递质的合成、释放以及转运体的活动。有文献报道,脑儿茶酚胺能神经元参与中枢给予血管紧张素Ⅱ引起的升压和饮水反应,其作用机制可能是通过血管紧张素Ⅱ作用于α-肾上腺素能受体或血管紧张素能 AT1受体,引起下丘脑去甲肾上腺素释放量明显增多,并呈显著的量效关系[15]。另研究发现,血管紧张素Ⅱ可激活磷脂酶 C和蛋白激酶 C传导通路,促进肾上腺素转运系统的活动[14]或增强酪氨酸羟化酶 mRNA、c-fos的表达[16],引起肾上腺素合成和释放增加。
综上所述,脑血管紧张素能通路和脑儿茶酚胺能神经通路共同参与侧脑室注射氨甲酰胆碱引起的肾促钠排泄反应。但其确切的细胞及分子生理机制还有待进一步的研究。
[1] CUNNINGHAM J T,BRUNO SB,HIGGS K A,et al. Intrapericardial procaine affects volume expansion-induced fos immunoreactivity in unanesthetized rats[J].Exper im ental Neurology,2002,174(2):181-192.
[2] DELUCA L A J R,FRANCIC R,SAAD W A,et al.Natriuresis,not seizures,induced by cholinergic stimulation of the locus coeruleus is affected by forebrain lesions and water deprivation[J].Brain Research Bulletin,1991,26(2):203-210.
[3] MCCANN SM,FRANCICR,FAVARETTO A L,et al.Neuroendocrine regulation of salt and water metabolism[J]. Brazilian Journal of Medical and Biological Research,2004,84(1):169-208.
[4] JIANG Chun-ling,ZHANG Wan-qin,L IN Maozhang(姜春玲,张万琴,林茂樟).The role of carbachol in natriuresis and the activity of TH-IR neurons in hypothalamus[J].Chinese Journal of Applied Physiology(中国应用生理学杂志),1999,15(1):35-38.(in Chinese)
[5] ZHU Chang-geng(朱长庚 ).Neuroanatomy(神经解剖学 )[M].Beijing:People’s Health Publishing House,2002:500-536.(in Chinese)
[6] SAAD W A,GUARDA IF,CAMARGO L A,et al.Adrenoceptors of the midial septal area modulate water intake and renal excretory function induced by central administration of angiotansin II[J].Brazilian Journal of Medical and Biological Research,2002,35(8):951-959.
[7] SAAD W A,LUIZ A C,CAMARGO L A,et al.Functional evidence that the central renninangiotensin system plays a role in the pressor response induced by central injection of carbachol[J]. Brazilian Journal of Medical and Biological Research,1997,30(4):493-496.
[8] MATHAL M L,EVERED M D,MCKINLEY M J.Central losartan blocks natriuretic,vasopressin and pressor response to central hypertonic NaCl in sheep[J].Regulatory,Integrative and Com parative Physiology,1998,275(2):548-554.
[9] KALOYANIDES G L,BALABANIAN M B,BOWMAN R L. Evidence that the brain participates in the humoral natriuretic mechanism of blood volume expansion in the dog[J]. The Journal of Clin ical Investigation,1978,62(6):1288-1295.
[10] ANTUNES-RODRIGUESJ,DE CASTROM,EL IAS L L,et al.Neuroendocrine control of body fluid metabolism[J].Physiological Rev iews,2004,84(1):169-208.
[11] BALDISSERA S,MENANIJW,DOS SANTOSL F,et al.Role of hypothalamus in the control of atrial natriuretic pep tide release[J].Proceedings of the National Academy of Sciences,1989,86(23):9621-9625.
[12] ITO S,GORDON F J,SVED A F.Dietary salt intake alters cardiovascular responses evoked from the rostral ventrolateral medulla[J].American Journal of Physiology,1999,276(6):1600-1607.
[13] BENARROCH E E,SCHMEICHEL A M.I mmunohistochemical localization of the angiotensin II type 1 receptor in human hypothalamus and brain stem [J]. Brain Research,1998,812(1-2):292-296.
[14] CAMARGO L A A,SAAD W A.Role of theα1-and α2-adrenoceptors of the paraventricular nucleus on the water and salt intake,renal excretion,and arterial p ressure induced by angiotensin IIinjection into the medial septal area[J].Bra in Research,2001,54(6):595-602.
[15] DE ARRUDA CAMARGO L A,SAAD W A,DE SOUZA V ILLA P. Interaction between paraventricular nucleus and medial septal area on the renal effects induced by adrenaline[J].Autonom ic Neuroscience:Basic and Clin ical,2004,111(2):135-139.
[16] YU K,LU D,ROWLAND N E,et al.Angiotensin II regulation of tyrosine hydroxylase gene exp ression in the neuronal culture of normotensive and spontaneously hypertensive rats[J].Endocr inology,1996,137(8):3566-3576.
Effect of AT1receptor on changes of thyrosine hydroxylase-immunoreactivity in rostral ven trolateral medulla induced by bra in cholinergic stimuli in rats
WANGM in1,YAO Qi-ying2,XIA Man-li1,ZHOU Xin-mei1,JIANG Chun-ling2
(1.Department of Physiology,Jiaxing University,School of Medicine,Jiaxing 314001,China;2.Department of Physiology,Dalian Medical University,Dalian 116027,Chian)
Objective:To investigate the effect of AT1receptor on the changes of thyro sine hydroxylase-immunoreactivity(TH-IR)in rostral ventrolateral medulla(RVLM)induced by brain cholinergic stimuli in rats.Methods:Male SD rats were randomly divided into 4 groups:NS+CBCgroup,Los+CBC group,Los+NS group and NS+NS group.AT1was blocked by pretreatment of 20μg losartan in Los+CBC and Los+NS groups;intracerebroventricular injection of 0.5μgcarbachol was used for cholinergic stimuli in NS+CBC and Los+CBC groups;normal saline(NS)was used for control.The output amount of natrium in kidney,glomerular filtration rate(GFR)and renal plasma flow(PRF)were observed.The changes of TH-IR in the RVLM were observed by immunohistochemistry.Results:In NS+CBC group carbachol induced potent natriuresis,after pretreatment of losartan the natriuretic effectwas partially inhibited in Los+CBC group.Both the number and op tical density of THIR positive neurons in NS+CBC group were markedly increased than those in NS+NS group(P<0.05);while those in Los+CBC group were significantly lower than those in NS+CBC group(P<0.05).Intracerebroventricular injection of carbachol and losartan had no effect on GFR and RPF(P>0.05).Conclusion:The results suggest that cholinergic stimuli can induce potent natriuresis and increase the activity of adrenergic neurons in the RVLM;the above effects can be down regulated by blockade of brain AT1recep tor.
Carbachol/pharmacol;Recep tor,angiotensin,Type 1/pharmacol;Medulla oblongata/drug eff;Losartan/pharmacol
R 334
A
1008-9292(2011)01-0071-07
http:∥www.journals.zju.edu.cn/med DO I:10.3785/j.issn.1008-9292.2011.01.013
2010-05-26
2010-11-24
浙江省嘉兴市科技局项目 (2008AY2039-4).
王 敏 (1979-),女,硕士 ,讲师;E-mail:wmwmwmtom@yahoo.com.cn
姜春玲 (1964-),女,教授,博导;主要从事神经和肾脏生理研究;E-mail:chunling_jiang2006@yahoo.com.cn
[责任编辑 黄晓花 ]
