天山北部拟南芥种群间形态和物候特征的选择分化
2011-01-05沈雪莹刘彤曾晓玲张卫宾
沈雪莹,刘彤 ,曾晓玲,张卫宾
天山北部拟南芥种群间形态和物候特征的选择分化
沈雪莹,刘彤 ,曾晓玲,张卫宾
(石河子大学生命科学学院,石河子832003)
将从天山北部不同地理区域采集的7个拟南芥种群在同质园中种植,研究干湿2种处理下不同种群的表型分化特点及其自然选择特征,分析拟南芥在不同地区表型分化的可能机制。结果表明:实验观测的株高、株高/根长、分支数等9个特征在种群间差异显著,其中分枝数、果实总数、株高/根长、成熟时间以及间隔时间5个特征在交互作用下呈现出显著差异。聚类分析发现2种处理下都将干旱地区种群归为一类,湿润地区的种群归为另一类,种群间特征的多重比较结果也进一步证明了这一点。回归分析结果表明,无论干旱还是湿润处理,自然选择对拟南芥各种群的株高、分枝数、莲座叶大小、株高/根长、叶片数等特征的选择均定向增加;干旱处理下,对额敏种群的开花物候的选择定向减小,而湿润条件下对青河2繁殖时间(间隔时间)的选择则为定向增加。总结认为:天山北部拟南芥种群间株高、分枝数、株高/根长、果实总数、开花时间、成熟时间、间隔时间、莲座叶大小和开花时叶片数已经发生了遗传特化,降水是其发生的主要环境因子,自然选择是种群间表型分化主要动力,但不排除地理隔离的因素作用。
拟南芥;自然选择;表型分化;遗传特化
植物表型是基因与环境相互作用的结果,在长期选择压力下形成稳定遗传而产生进化[1]。由于环境是波动的,而植物个体可以根据内外部而调整基因表达、翻译、转录,进而产生表达不同表型的能力-表型可塑性对于实现植物当代的繁殖成功具有重要意义[2],而如果长期地经历同样的选择,则可能导致表型分化进而产生进化[1,3]。
开花时间是植物生活史的关键特征,它决定了花部发育、授粉和种子生产[4],显著的贡献于植物适合度[5]。在自然选择下,植物形成了对水分、光照、温度以及生物因子等环境信号整合调节机制,最终形成现存最优的开花时间。众多研究表明,不同区域降雨量的差异是导致种群间物候的提前或延迟的重要原因[6-8],即在干旱条件下,植物花期倾向于定向选择,向着早开花的方向进化[4,9-11]。但由于环境波动的异质性、表型间特征整合性、系统发育约束性,以及物种的环境特化等,在干旱条件下开花时间的进化快慢和方向存在诸多变化。
拟南芥(Ar abidopsis thaliana)是植物生物学的模式植物,目前拟南芥表型进化的机制研究引起广泛重视,其中不同种群表型特征变化、所经受的自然选择以及变化趋势等是了解表型进化的重要内容,至今学者们已经对食草动物取食[12]、风[13]、水淹[14]、光照[15-17]、养分胁迫[18]、纬度[19]等对拟南芥表型分化的影响进行了研究,但是不同降水条件下拟南芥种群表型及物候的空间变异研究还很少。水分是限制拟南芥分布的重要因素[20],尤其在干旱区,水分更是决定了植物物种的分布格局[21-22],所以水分对拟南芥表型的影响以及导致种群间的差异尤其应该引起重视。
新疆的天山山脉是世界拟南芥及其近缘种的分布中心之一[23],我们调查发现不仅天山山脉,而且新疆北部的塔尔巴哈台山脉、阿尔泰山脉均有拟南芥分布[24-25]。天山北部拟南芥种群分布地区降水量不同,且土壤理化性质、地形等环境因素差异较大,自然居群的表型存在差异[26],这表明不同地区拟南芥种群表型分化经历了不同的进化过程。
自然选择是物种进化的主要动力,而突变、漂变、迁移、地理隔离等同样对物种进化产生作用。从天山北部拟南芥种群的空间分布来看,各种群间隔距离远近不同,生境多样且地理结构也不尽相同,所以天山北部拟南芥各种群表型分化的研究为深入分析拟南芥适应与进化过程提供了良好条件。那么,天山北部拟南芥种群表型分化的动力是什么?拟南芥种群的表型是否发生了分化?导致这种表型分化的主导因素是什么?各种群表型特征进化趋势是什么等问题极有必要深入研究。为此,我们将从天山北部3个地区采集的7个种群在同质园中种植,设置干旱和湿润2种环境条件,研究不同种群的表型分化特点和自然选择趋势,以期为分析天山北部拟南芥的表型进化机制奠定基础。
1 材料与方法
1.1 材料
1.1.1 实验样地
实验样地设置在天山北坡垂直带下石河子大学实验站的试验田中(同质园)。这一区域是短命植物主要的分布地带,且与当地的自然种群只相距十几公里的自然生境,N=44°15′,E=56°21′,海拔442 m,年降雨量约200 mm,年均温度8℃。
1.1.2 实验材料
2009年6月初,我们在天山北部5个区域的7个样地(表1)中收集拟南芥种子,每个样地均选择植株间相距3 m以上,约50株进行单株采种。
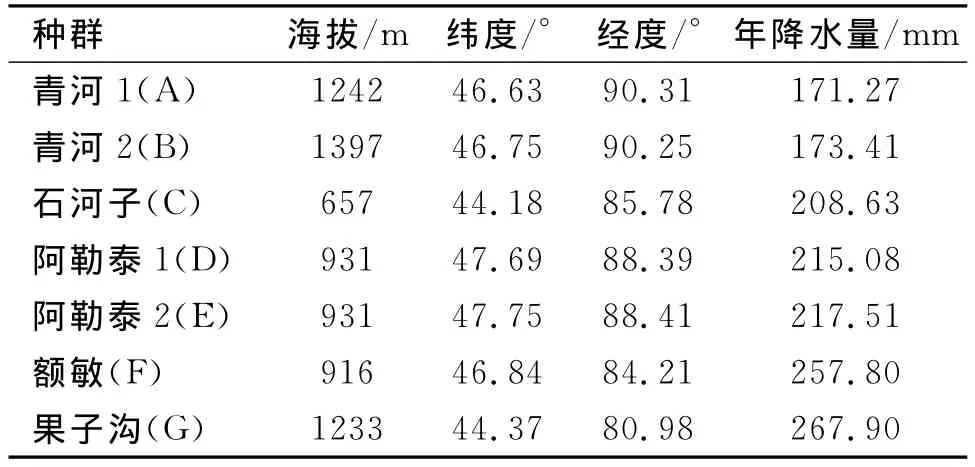
表1 天山北部拟南芥各种群的地理特征Tab.1 Geographical features on the origins of the seven studied populations of A.thaliana at north of Tianshan Mountains
根据表1中不同种群的降水特点以及李磊等[25]的研究结果,我们将青河1、青河2、石河子、阿勒泰1分成干旱种群,果子沟、额敏和阿勒泰2划为湿润种群,这是由于野外调查发现阿勒泰2种群生长在较陡的山体岩石旁或缝隙里,因有岩石庇荫,因此土壤比阿勒泰1的潮湿,所以将其划分为湿润种群。
1.2 方法
2010年3月底我们在同质园中进行实验,种植来自不同地理区域的种子,并在植物个体发生的不同时期进行干湿2种梯度的水分处理。试验采用裂区设计,每个小区撒播不同种群的拟南芥种子50粒,出苗2周后间苗保证每个小区5个植株,每个种群每种水分处理下10个小区,因此共有供试植株700株(7个种群×2种处理×10个小区×5个植株)。
自第1株幼苗出土起就开始观察各种群的出苗情况。对每个抽薹植株进行挂牌标记,并开始进行干湿的水分处理(类似于自然生境的降水时期),每天观察其物候和生长发育特征。水分处理时让湿润样地的浇水量是干旱样地的2倍(在试验样地铺设喷灌带,用半径为1.5 m的雾化喷头以喷灌的方式进行浇水,而且在干湿样地的地头各设置了1个开关,通过控制浇水时间来控制降水量)。采用搭建临时遮雨棚形式,防止自然降雨对实验结果造成影响。在各种群85%的植株成熟时进行统一采收,在实验室测量不同种群植株个体的形态特征。
我们供观测了3组特征:营养特征、结构特征和繁殖特征。
营养特征:开花时莲座叶大小、莲座叶的数量;
结构特征:株高、分枝数;
繁殖特征:第一开花时间(简称开花时间)、第一成熟时间(简称成熟时间)、间隔时间(表示从开花到成熟之间的天数)和果实总数。
用游标卡尺测量果实长度(依每个植株果实多少,选取上中下各3个(共9个)有代表性果实测量果实长度取平均值进行分析)和根长,直尺测量株高。在开花时用直尺测量其莲座叶的最长直径(每个植株任取3片莲座叶进行测量,取平均值)作为开花时莲座叶的大小。
1.3 数据处理
对株高、株高/根长、分枝数、果实总数、开花时莲座叶大小、开花时叶片数以及花期、成熟期、间隔时间等重要形态和物候与水分处理进行二元方差分析,研究拟南芥种群间和处理间形态和物候的差异性。分别用干、湿处理下的上述9个重要形态和物候特征进行聚类分析,研究拟南芥种群表型分化的主要自然选择压力是水分。为了探讨不同环境下拟南芥的自然选择类型和方向,将以上9个特征分别与适合度(果实总数)进行回归分析[27-28]。其符合形如Y=a X+b的线性模型,其中Y代表适合度;X代表各个表型特征;a表示每个变量X(特征)和Y(适合度)之间关系的偏回归斜率,b为截距。
所有数据均利用SPSS 16.0进行数据分析,Origin Pro 7.5辅助作图。
2 结果与分析
2.1 拟南芥种群间的遗传变异
由表2可见,实验所观测的9个特征在种群间都存在显著差异。除了间隔时间和开花时叶片数在干湿处理间变化不显著外,其余特征在干湿处理下变化均达到极显著差异。在种群与处理的交互作用下9个特征中有5个特征(分枝数、果实总数、株高/根长、成熟时间以及间隔时间)均呈现出显著差异,表明种群间的表型可塑性特征发生了遗传分化[14]。
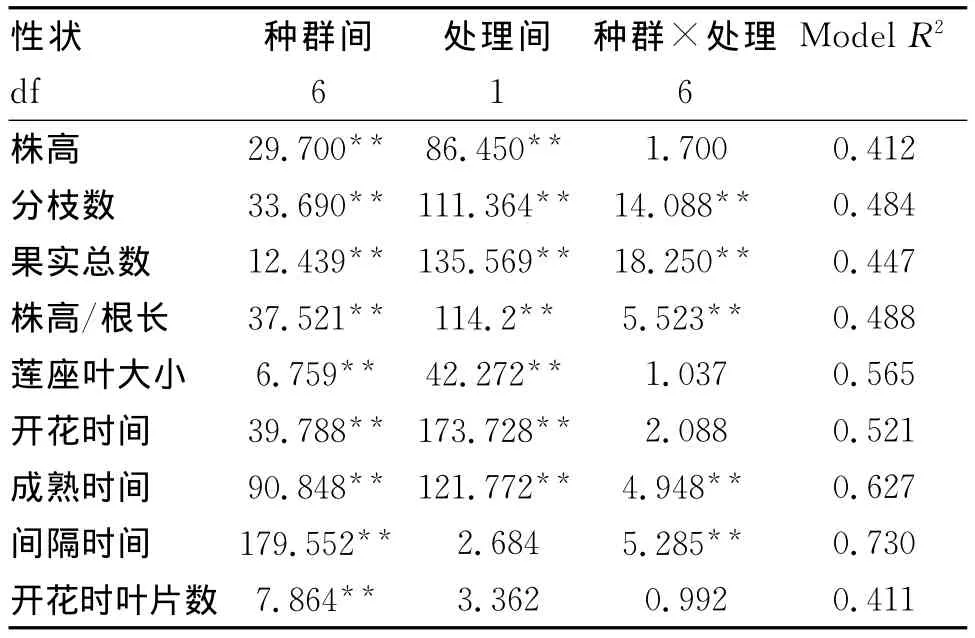
表2 不同种群干湿处理下9个特征的方差分析Tab.2 ANOVA of nine traits among populations at dry and wet treat ments
多重比较结果(表3)显示,干旱处理下,株高、株高/根长、开花时间、成熟时间和间隔时间这9表型个特征中有5个特征都表现出:干旱种源的4个种群之间几乎无显著差异,湿润种源的3个种群间差异不显著。尤其是比较果子沟、额敏和阿勒泰2这3个湿润种群时,种群间所有特征均无显著差异。
由表4可见,9个表型特征中有5个特征将所有种群分成两部分(即分枝数、果实总数、开花时间、成熟时间、间隔时间),这5个特征在湿润种源的3个种群间无显著差异,而在干旱种源间4个种群间各特征也无显著差异。除了株高/根长和开花时叶片数外,干旱种源种群间各特征均无显著差异;而湿润种群间除了株高/根长外,其余所有特征也均为差异不显著。
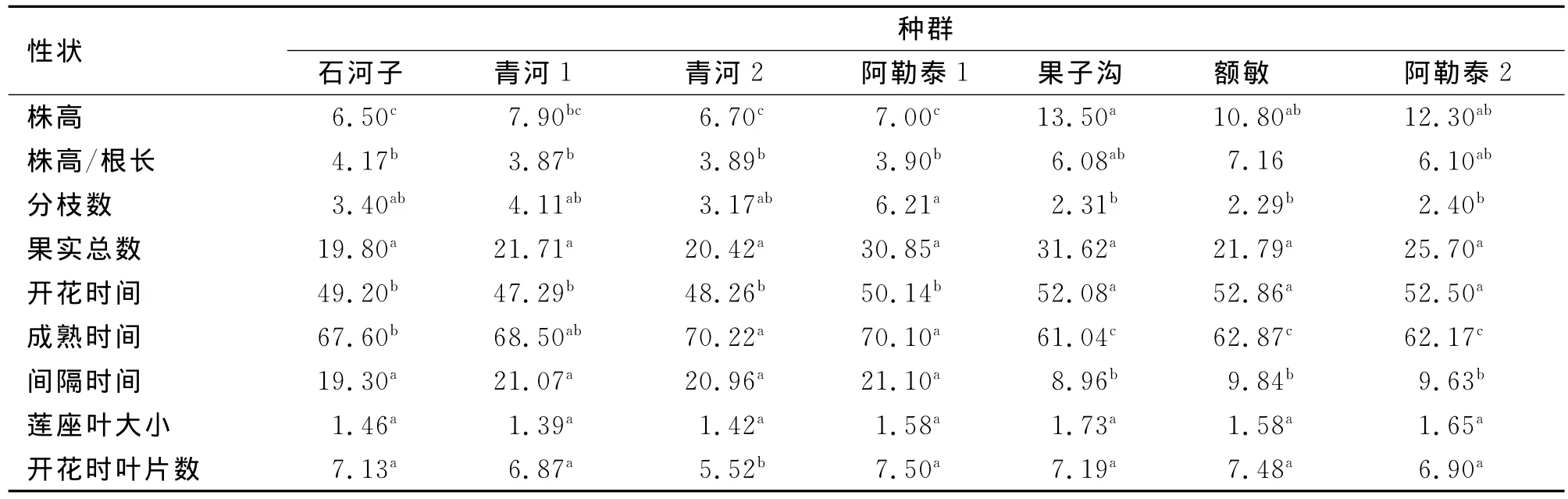
表3 干旱处理下拟南芥表型性状的方差分析Tab.3 Variance analysis of phenotypic characters among A.thaliana populations at dry treat ment
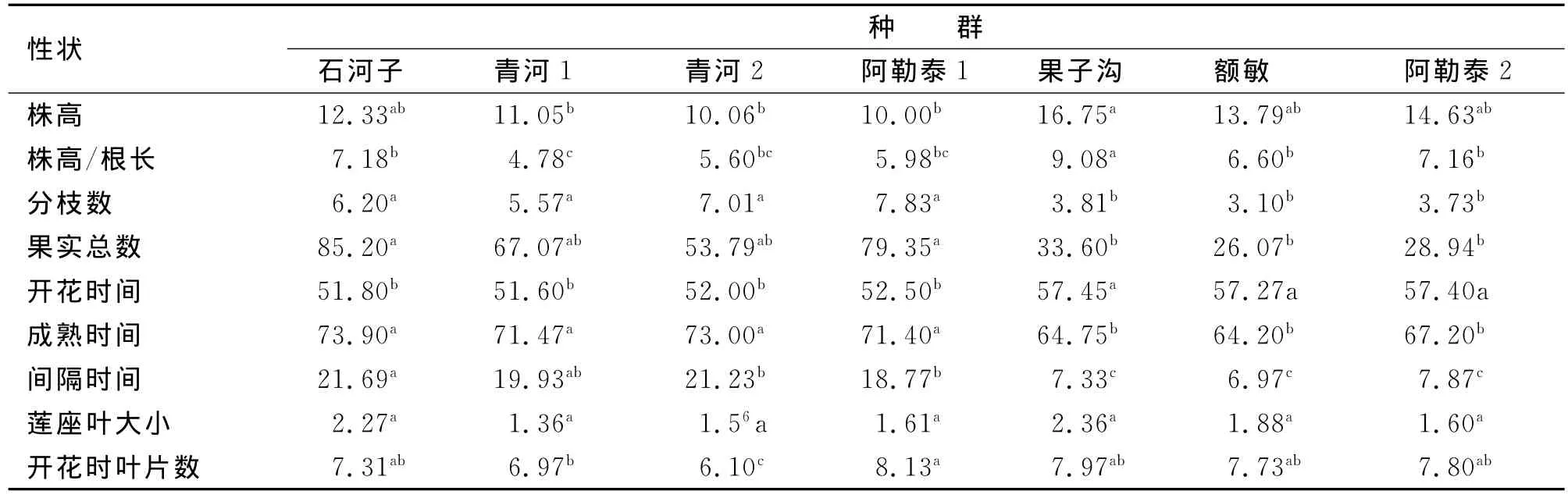
表4 湿润处理下拟南芥表型性状的方差分析Tab.4 Variance analysis of phenotypic characters among A.thaliana populations at wet treat ment
2.2 水分处理下拟南芥种群表型特征的整体相似性
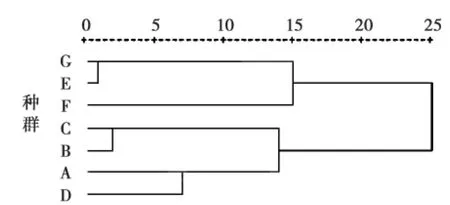
图1 干旱处理下各种群表型特征的相似性聚类分析Fig.1 Hierarchical cluster analysis based on similarity of phenotypic traits of seven populations of A.thaliana at dry treat ment
通过对包括物候、形态共9个表型特征进行聚类分析,结果(图1、图2)显示虽然单个特征在种群间差异都显著,但是整体特征在有些种群间差异却并不显著,如阿勒泰2和果子沟最先聚在一起,这表明这2个种群间植株的整体相似性最强。另外,无论是干旱处理还是湿润处理都将干旱种源的种群归为一类(青河1、青河2、石河子、阿勒泰1),湿润种源的种群归为另一类(额敏、果子沟、阿勒泰2),此结果与干、湿处理下种群间各特征的多重比较的结果相一致。这也反映出在干旱种源的种群间或湿润种源的种群间其整体表型均无显著差异,表明导致本地拟南芥各种群表型进化的主要动力为水分。
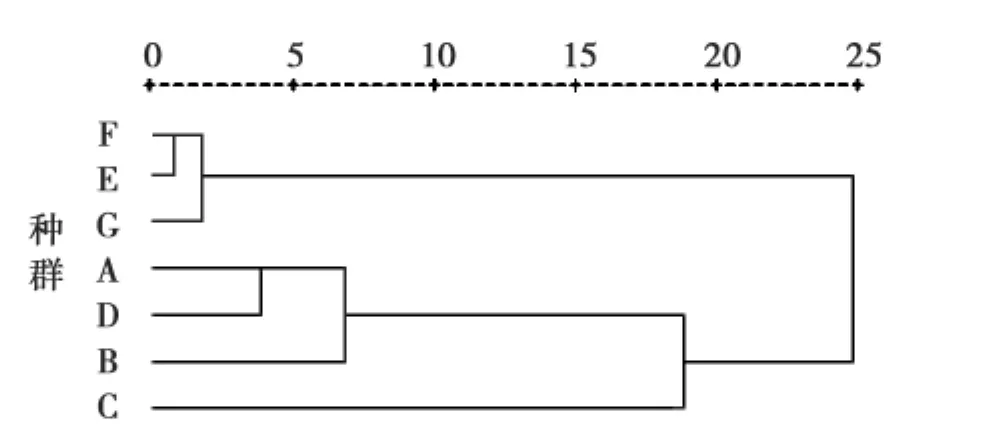
图2 湿润处理下各种群表型特征的相似性聚类分析Fig.2 Hierarchical cluster analysis based on similarity of phenotypic traits of seven populations of A.thaliana at wet treat ment
2.3 干湿条件下各特征的自然选择类型及方向
回归分析结果(表5、表6)显示,无论干旱还是湿润处理下,自然选择对拟南芥各种群的株高、分枝数、莲座叶大小、株高/根长、叶片数等相同特征的选择均是定向增加的,即植株向增加株高方向发展、分枝数、叶片数向增多的方向选择而莲座叶大小、株高/根长也向增大方向发展。此外在干旱处理下,对额敏种群的开花物候的选择是定向减小的,而在湿润条件下对青河2繁殖时间(间隔时间)的选择则是定向增加即朝延迟方向选择。
综合表5和表6,我们还发现:绝大多数种群的株高和分枝数,无论在干旱还是湿润条件下都与适合度具有极显著相关。青河1和青河2在干旱下与适合度相关的特征较少,而在湿润下时却较多;果子沟、额敏以及阿勒泰2与之相反,在湿润条件下对适合度有显著贡献的特征比在干旱下相对较多;而石河子和阿勒泰1在干湿条件下对适合度有选择贡献的特征相同,均是株高和分枝数。

表5 干旱处理下对拟南芥表型特征的自然选择分析Tab.5 Natural selection analysis of phenotypic characters of A.thaliana in arid condition
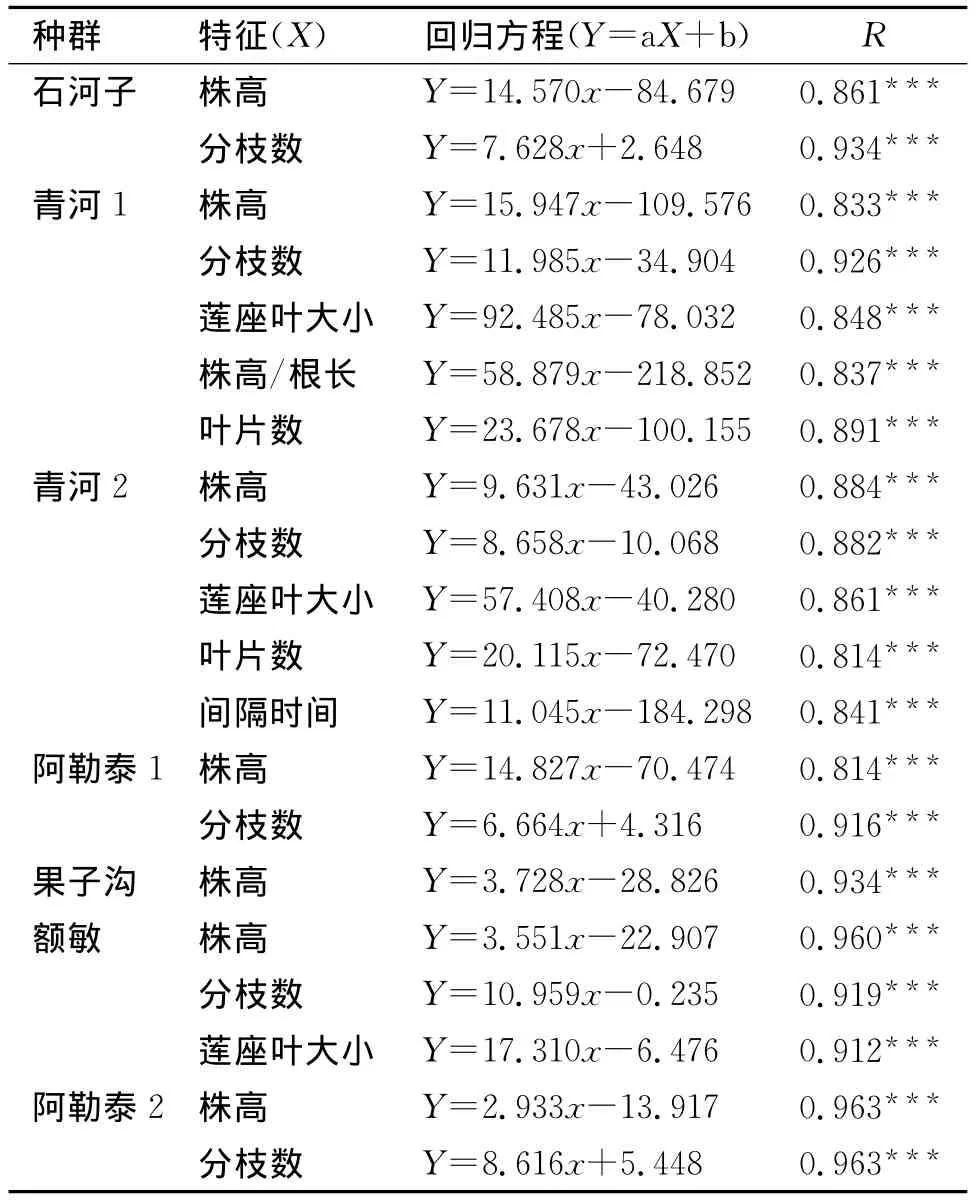
表6 湿润处理下对拟南芥表型特征的自然选择分析Tab.6 Natural selection analysis of phenotypic characters of A.thaliana in wet condition
3 讨论与结论
3.1 拟南芥各种群的表型分化特点
我们通过将野外自然条件下降雨量和空间结构存在差异的拟南芥种群种植到同质园中进行观察研究,发现这些种群间的形态和物候特征均存在显著差异,这表明各种群通过长期的自然选择均发生了显著的遗传分化;除了花时叶片数和间隔时间外,各特征在处理间也存在显著差异,这就说明基因型和降雨量均对拟南芥的这些特征具有显著影响,且这些特征也具有广泛的表型可塑性。9个特征中有5个表现出了可塑性的遗传分化,这表明各种群已经对当地环境产生了遗传特化。
在野外自然生境下植株的生长状态,反映了各种群在自然选择下适应状态,研究发现各种群间的株高和果实总数均达到显著变化,而分枝数差异不显著[25],但在同质园种植,去除环境因素干扰后发现株高和果实总数与野外自然生境相同,也达到了显著差异[26];而分枝数在种群间、处理间以及交互作用下均表现出极显著的差异。我们通过标准样地实验发现干旱地区种群的分枝数在干湿处理间变化显著(可塑性较大),而湿润种群在干湿处理下分枝数变化却不显著(可塑性相对较小)。这表明分枝数不仅具有较强的可塑性,而且在不同地点发生了不同的遗传特化,自然生境下表现的差异不显著很可能由于种群内特征值变化较大,导致组内方差大而形成的种群间差异不显著。
开花时间可以通过多样化的方式影响个体和种群的适合度,开花和种子产生必须符合一定季节,开花和生长必须在条件恶劣之前结束[29]。开花物候的高度变异,明显有助于该物种扩张其生存范围[30],并且在气候较为恶劣的时期也能存活。有研究表明植物对季节信号的最佳响应可能随地理变化而存在不同,从而导致开花时间存在地理差异,如在不同地理区域随纬度、海拔和气候在不同种群的物候存在明显的分化[29,31]。
我们的研究结果显示,开花天数在种群间和处理间变异显著,交互作用并不显著,这说明种群间开花时间的变异格局在2种水分处理下并没有改变,虽然干旱条件下,各种群的开花时间有所提前,但是这种差异依然较小。这可能是对于像拟南芥这样的早春开花的1年生植物而言,其开花时间除了受降水影响外,还与光周期、温度等因素有关[32-34]。
尽管在我们的研究中开花时间没有可塑性的遗传变异,但是成熟时间的可塑性遗传变异却很明显,表明拟南芥在胁迫环境下,除开花时间调整外,从开花到果实成熟之间的间隔也可以发生可塑性变化,这是以往研究所忽略之处。
3.2 天山北部拟南芥种群间表型分化的动力
自然选择是环境作用于生物的综合体,尽管降水、经纬度、坡度、坡位等构成了拟南芥的自然选择因素,但是李磊等[26]研究发现拟南芥主要分布于p H值和HCO3-含量低,有机质丰富,且有一定坡度的沙土地块上,其生长和分布还受到微环境的影响。
干旱和湿润对植物来说是两种截然不同的环境压力,于是会引出植物表型不同的变异与协变机制[35-36],最终通过表型变异呈现出来。虽然拟南芥在生长过程中可能经历波动的环境变化,但该物种通过1年生生活史特征、丰富的表型可塑性、控制种子扩散等方式适应环境。我们通过表型的聚类分析发现来自干旱地区的种群归为一类,而来自湿润地区的种群归为另一类,这表明干旱地区种群间或湿润地区种群间其整体特征差异不明显,这也证明了在水分波动的长期作用下,各种群形成了相似的整合格局,拟南芥对环境的适应是多个特征共同作用的整体适应,种群间的形态和物候分化是由区域性异质性降水所驱动。
3.3 天山北部拟南芥自然选择的类型及方向
干湿环境下对株高、分枝数、莲座叶大小等特征均产生递增的定向选择,说明无论环境如何,植物对空间结构(aerial str uctures)的投资均有利;同时也从一定程度上表明水分(降水)处理对这些特征产生的影响较大,进而会通过上述特征显著地影响繁殖适合度。
对于株高、分枝数来说,在所有种群所有处理中只有干旱处理下青河2的株高和湿润处理下果子沟的分枝数对适合度贡献不显著。这可能反映出它们在对各自局部生境的适应过程中发生了特化,形成了自身独特的适应对策。
尽管各种群的物候在干旱条件下提前,湿润条件下相对延迟,但是在选择分析中只有对额敏开花时间的选择在干旱条件定向减少,对青河2的间隔时间在湿润条件下定向增加。其余种群的物候没有表现出此类选择,这可能是其它种群物候与自身其它特征的协同作用减弱了其对适合度的直接贡献[37-38]。这也体现了拟南芥对干湿环境的适应不仅仅是物候与适合度的直接关系,而是自身的适应特性与形态特征以及物候等特征的综合作用的结果。
此外,我们发现的青河的2个种群在干旱下与适合度相关的特征较少,而在湿润下时却较多;果子沟、额敏以及阿勒泰2与之相反。这可能是由于来自干旱地区的种群对干旱环境已经形成了很好的适应,而突如其来的湿润对它来可能是一种胁迫,于是它必需通过定向的增加其它特征来共同调节适合度使其达到最优;对来自湿润地区的种群同理。石河子和阿勒泰1这2个种群在干湿下对适合度有选择贡献的特征始终相同,这可能和它们所处地理区域的降雨量有关,因为从表1中可以看到这2个地区的降雨量是这些种群中居中的(特殊的阿勒泰2除外),2个地区的降雨波动性可能较大,于是它们无论是对干旱环境还是湿润环境都能较好的适应,相对于其它种群,因此表现出此现象。不同种群对干湿环境变化的响应不同,这正反映出不同种群在过去一直存在的降水不同的情况下已经发生了遗传特化,由此对相同环境产生了不同的适应对策。
综上所述,拟南芥当前表型特征是过去自然选择的产物,又是未来变化的基础。因此我们从同质园中干旱、湿润处理下的表型变化特点,可以推断:促使天山北部各种群发生表型分化的主要环境因子是降水,主要动力是不同强度的自然选择。
[1]Pigliucci M,Murren C.Phenotypic plasticity and evolution by genetic assi milation[J].Jour nal of Experi mental Biology,2006,209(12):2362-2367.
[2]Schlichting C D,Smith H.Phenotypic plasticity:linking molecular mechanis m with evolutionary outco mes[J].Evolutionary Ecology,2002,16,189-211.
[3]高乐旋,陈家宽,杨继.表型可塑性变异的生态-发育机制及其进化意义[J].植物分类学报,2008,46(4):441-451.
[4]Shindo C,Ber nasconi G.Natural genetic variation in Arabidopsis:tools,traits and pr ospects for evolutionary ecology[J].Annals of Botany,2007,99(6):1-12.
[5]Stearns S C.The evolution of life histories[M].Oxford:Oxf ord University Press,1992.
[6]Shinoda M,Ito S,Nachinshonhor G U,et al.Phenology of mongolian grasslands and moisture conditions[J].J Meteorol Soc Jpn,2007,85:359-367.
[7]Palta J A,Tur ner N C,French R J,et al.Physiological responses of lupin genotypes to ter minal drought in a Mediterranean-type environ ment[J].Ann Appl Biol,2007,150:269-279.
[8]Bull-Hereñu K,Arr oyo M T K.Phenological and morphological differentiation in annual Chaetanthera moenchioides(Asteraceae)over an aridity gradient[J].Plant Syst Evol,2009,278:159-167.
[9]Callahan H S,Pigliucci M.Shade-induced plasticity and its ecological significance in wild populations of Ar abidopsis thaliana[J].Ecology,2002,83:1965-1980.
[10]Mc Kay J K,Richar ds J H,Mitchell-Olds T.Genetics of drought adaptation in Ar abidopsis thaliana:I.Pleiotropy contributes to genetic correlations among ecological traits[J].Molecular Ecology,2003,12:1137-1151.
[11]Pigliucci M.Selection in a model system:ecological genetics of flowering ti me in Ar abidopsis thaliana[J].E-cology,2003,84:1700-1712.
[12]Arany A M,De Jong T J,Van der Meijden E.Her bivor y and local genetic differentiation in nat ural populations of Ar abidopsis thaliana (Brassicaceae)[J].Herbaceous Plant Ecology,2009,201(2):287-295.
[13]Bossdorf O,Pigliucci M.Plasticity to wind is modular and genetically variable in Arabidopsis thaliana[J].Evol Ecol,2009,23:669-685.
[14]Pigliucci M,Kolodynska A.Phenotypic plasticity and integration in response to flooded conditions in natural accessions of Arabidopsis thaliana (L.)Heynh(Brassicaceae)[J].Annals of Botany,2002,90(2):199-207.
[15]Pigliucci M.Phenotypic plasticity to light intensity in Arabidopsis thaliana:invariance of reaction nor ms and phenotypic integration[J].Evolutionary Ecology,2002,16:27-47.
[16]Hans K.Stenøien.Quantifying latitudinal clines to light responses in natural populations of Arabidopsis thaliana(Brassicaceae)[J].American Journal of Botany,2002,89(10):1604-1608.
[17]Petrov A P,Petrosov V A.Variability in some lines of Arabidopsis thaliana (L.)Heynh.under different lighting conditions[J].Genetika,1981,16,1596-1602.
[18]Pigliucci M,Schlichting C D.Reaction nor ms of Ar abidopsis.V.Flowering ti me contr ols phenotypic architecture in response to nutrient stress[J].J Evol Biol,1998,11:285-301.
[19]Stinchcombe J R,Weinig C,Ungerer M,et al.A latitudinal cline in flowering ti me in Ar abidopsis thaliana modulated by the flowering ti me gene FRIGIDA[J].Proc Natl Acad Sci USA,2004,101:4712-4717.
[20]Hoff mann M H.Biogeography of Arabidopsis thaliana(L.)Heynh.(Brassicaceae)[J].Journal of Biogeography,2002,29,125-134.
[21]Gutter man Y.Seed ger mination in desert plants[M].Berlin:Springer-Verlag,1993:20-21.
[22]Mustart P J,Cowling R M.Effects of soil and seed characteristics on seed ger mination and their possible roles in deter mining field emergence patterns of four Agulhas Plain(South Africa)Proteaceae[J].Canadian Journal of Botany,1993,71:1363-1368.
[23]Price R A,Pal mer J A,A1-Shehbaz I A.Systematic relationships of Ar abidopsis:a molecular and mor phologyical perspective.In:Meyerowitz E,So merville C R.eds.Ar abidopsis[C].New Yor k:Cold Spring Har bour Laboratory Press,1994:7-19.
[24]陶冶,王丹,刘彤,等.天山北部拟南芥生存群落特征及其与环境的关系[J].生物多样性,2009,17(1):51-61.
[25]李磊,刘彤,刘斌,等.新疆北部拟南芥种群内和种群间的表型变异与协变[J].生物多样性,2010,18(5):497-508.
[26]李磊,刘彤,刘斌,等.天山北部拟南芥自然居群表型变化的环境依赖特征[J].植物生态学报,2010,34(11):1310-1318.
[27]Lande R,Aausher S J.The measurement of selection on correlated characters[J].Evolution,1983,37:1210-1226.
[28]Chevin L M,Lande R,Mace G M.Adaptation,plastici-ty,and extinction in a changing environ ment:towards a predictive t heor y[J].PLoS Biology,2010,8(4):1-8.
[29]Riihi mäki M,Savolainen O.Environ mental and genetic effects on flowering differences bet ween northern and souther n populations of Ar abidopsis Lyr ata (Brassicaceae)[J].American Journal of Botany,2004,91(7):1036-1045.
[30]Ehr man T,Cocks P S.Reproductive patterns in annual legume species on an aridity gradient[J].Vegetatio,1996,122:47-59.
[31]Olsson K,ÅÅgren J.Latitudinal population differentiation in phenology,life histor y and flower morphology in the perennial herb Lythr um salicaria[J].J Evol Biol,2002,15:983-996.
[32]Simpson G G.The Bald win effect[J].Evolution,1953,7:110-117.
[33]Stinchco mbe J R,Caicedo A L,Hopkins R,et al.Ver-nalization sensitivity in Ar abidopsis thaliana (brassicaceae):t he effects of latitude and flc variation[J].A-merican Jour nal of Botany,2005,92(10):1701-1707.
[34]Tan F,Swain S M.Genetics of flower initiation and development in annual and perennial plants[J].Physiol Plant,2006,128:8-17.
[35]Zhang J,Van-Toai T,Huynh L,et al.Deveop ment of flooding-tolenrant Arabidopsis thaliana by autore-gulated cytokinin production[J].Molecular Breeding,2000,6:135-144.
[36]Galen C.High and dry:drought stress,sex-allocation tradeoffs,and selection on flower size in the alpine wildflower Polemonium viscosum (Polemoniaceae)[J].American Naturalist,2000,156,72-83.
[37]王丹,刘彤,蒋成国,等.天山北麓拟南芥表型特征的自然选择[J].生态学杂志,2009,28(5):804-810.
[38]刘彤.拟南芥表型选择及适应性可塑性[D].南京:南京林业大学,2004.
The Select Differentiation of Mor phology and Phenology among Ar abidopsis thaliana Populations in North Tianshan
SHEN Xueying,LIU Tong,ZENG Xiaoling,ZHANG Weibin
(College of Life Science,Shihezi University,Shihezi 832003,China)
We planted the seeds of seven populations from different geographical regions in northern Tianshan in the common garden,and set the wet and dry treat ments.In order to investigate the characteristics of the phenotypic differentiation and the trend of natural selection in different populations,analyzing possible mechanis m that phenotypic differentiation of Arabidopsis thaliana in different regions.The results show there are nine characteristics of experi mental observations have significant differences among populations,including shoot height,branch nu mber,nu mber of fr uits and so on.Of which five features are branch nu mber,nu mber of fr uits,shoot height/root lengt h,ti me to maturation and inter val days respectively,in the interaction showed significant differences.Cluster analysis f ound that populations of arid regions classified as a class,wet populations classified as another group under dry and wet treat ment both are.Multiple comparisons bet ween the characteristics of populations f urther proved t his point.Regression analysis showed that both dr y or wet treat ments,natural selection of plant height,branch nu mber and other feat ures among populations in Ar abidopsis thaliana was directed to increase;dry treat ment,to the choice of Emin population's flowering phenology is directional reduce,however,to natural selection of Qinghe2 breeding ti me (the interval in days)was directed to increase under wet treat ment.The conclusion is that shoot height,branch number,number of fruits,shoot height/root length,rosette size,time to flowering,time to maturation,interval days and leaf nu mber at flowering have occurred genetic specialization among populations of Ar abidopsis thaliana in norther n Tianshan Mountains,precipitation is t he main environ mental factors of its occurrence,natural selection is the main driving force of phenotypic differentiation among populations.
Ar abidopsis thaliana;natural selection;phenotypic differentiation;genetic specialization
Q948
A
1007-7383(2011)04-0408-08
2011-01-20
国家自然科学基金项目(30760047)
沈雪莹(1984-),女,硕士生,专业方向为植物生态学;e-mail:shenxueying999@163.co m。通讯作者:刘彤(1968-),男,教授,从事植物生态学研究;e-mail:liutong1968@yahoo.com.cn。
