南海北部浮游生态系统要素季节变化的模拟研究*
2011-01-05高会旺高振会
张 冲,史 洁,高会旺**,高振会
(1.中国海洋大学海洋环境与生态教育部重点实验室,山东青岛266100;2.国家海洋局北海环境监测中心,山东青岛266100;3.国家海洋环境监测中心,辽宁大连116023)
南海北部浮游生态系统要素季节变化的模拟研究*
张 冲1,3,史 洁1,高会旺1**,高振会2
(1.中国海洋大学海洋环境与生态教育部重点实验室,山东青岛266100;2.国家海洋局北海环境监测中心,山东青岛266100;3.国家海洋环境监测中心,辽宁大连116023)
本文基于南海SEATS站的观测资料,利用垂直一维海洋生态系统动力学模式(MEM-1D)模拟了南海北部生态要素垂直结构的季节变化。结果表明,南海北部表层Chla浓度具有明显的季节变化特征,夏季呈现低值(<0.05 mg.m-3),冬季呈现高值,12月为0.38 mg.m-3。春季,风速相对变小,垂直混合变弱,次表层Chla浓度极大值现象形成(80~100 m),并可一直持续到10月末。冬季,海表温度降低,东北季风爆发,垂直混合增强,出现表层水华现象。浮游动物和细菌的垂直结构相似,冬季上层水体中浮游动物和细菌出现最大值,分别为13 mg.m-3和18 mg.m-3。3~10月细菌(12 mg.m-3)同浮游动物(14 mg.m-3)一样出现次表层最大值,其存在深度与浮游动物相似。
海洋浮游生态系统;台风;叶绿素a;水华;模拟研究
浮游生物是海洋生态系统中最基础的组成部分,在整个食物链物质循环和能量流动中扮演着重要的角色,而且浮游植物每年贡献了全球一半的氧气[1],因此,对浮游生态系统的研究一直是海洋科学研究关注的焦点问题[2]。我国南海地处东亚季风区和西太平洋暖池区,水文特征和生态系统结构特点鲜明,是研究气候变化与海洋生态响应的天然实验室[3]。卫星资料和船载观测资料分析表明,由于受季风作用,南海初级生产力呈明显季节性变化,冬季高,夏季低[4]。Liu等[3]的模拟结果表明,南海初级生产力年际变化与季节性变化相比,并不明显,其年平均值为196 mg.m-3.d-1。
南海北部与人类活动关系密切,近些年随着科考航次的增多,对南部北部生化要素研究也逐渐深入。Tang等[5]利用卫星数据分析了南海北部湾初级生产力的时空变化;Chen等[6]通过2001—2004年5个航次的观测数据计算了南海北部的新生产力,分析影响其变化的因素;基于南海SEATS站(18°N,116°E)的观测资料,诸多学者对南海北部生态系统中各要素(如浮游植物生物量季节变化和碳循环等)的季节变化做了一些研究[7-10],然而对南海北部浮游生态系统中各要素变化之间相互关系研究较少。数值模型计算可以得到各个状态变量的连续变化,是定量研究生态系统动力学的有力手段,在我国其他海区已有广泛的应用[11]。南海生化要素水平分布较均匀,其垂直结构变化受到更多的关注[7]。本文利用垂直一维海洋生态系统动力学模式(MEM-1D)模拟了南海北部浮游生态系统在风和太阳辐射等因素强迫下的季节变化,定量分析了营养盐浓度、浮游植物、浮游动物及其细菌生物量的变化特征,以及它们之间相互影响和制约关系,并且与其他海域生态要素季节变化特征作了对比。
1 物理-生物耦合模型(MEM-1D)
本文采用的海洋生态动力学模式为一维物理-生物耦合模式(Modular Ecosystem Model-1D,MEM-1D)[12],其物理亚模型采用一维POM模式(Princeton Ocean Model),生物亚模型采用ERSEM模式(EuropeanRegionalSeaEcosystemModel,ERSEM-II,1997)。
1.1 物理模型
由于POM模型的湍混合模型采用传统的2阶湍流闭合模型(Mellor and Yamada),未考虑表层海浪混合作用,模拟出的混合层较浅[13],夏洁等[14]在该模式垂直混合系数的计算中考虑了海浪的作用,较好的模拟了南黄海混合层季节变化规律。本文物理模型部分采用相同的修改方案。另外,本文所选用POM一维模式垂向上只考虑湍扩散过程。
1.2 生物模型
生物模型为欧洲区域海洋生态模式ERSEM[15],其状态变量包括浮游植物(P)、浮游动物(Z)、营养盐(N)、底栖碎屑(D)和微生物(B)等。
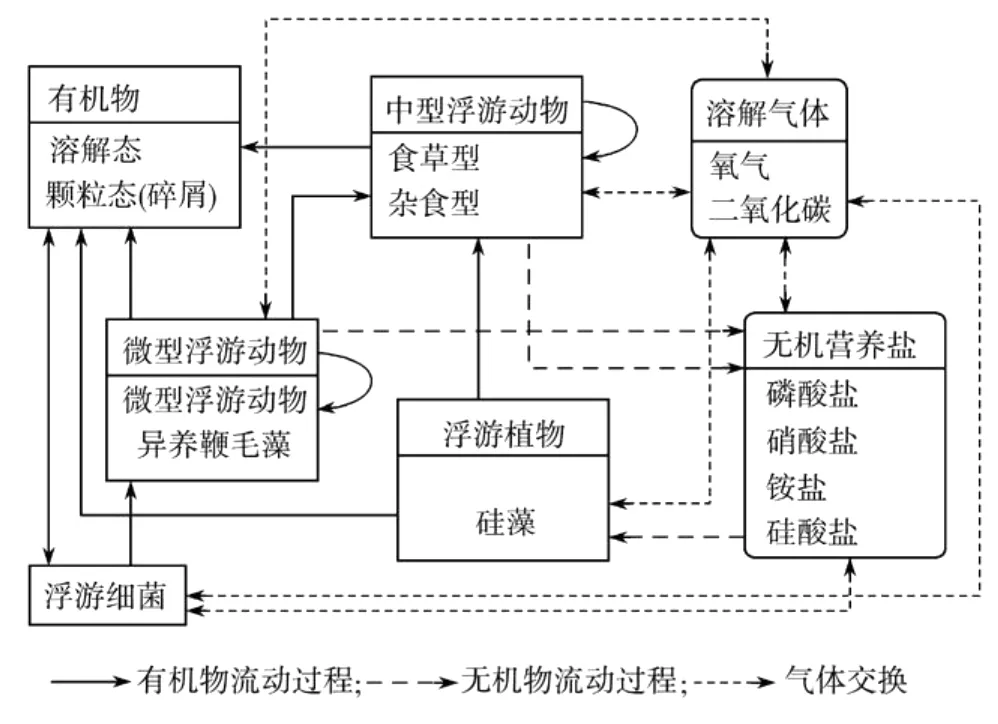
图1 模型食物网结构图Fig.1 Food web structure in the model
以往研究表明硅藻为南海的优势种群,冬季硅藻约占浮游植物的95.7%,夏季占75.9%[16]。因此,本模式中浮游植物仅考虑硅藻。浮游动物包括:杂食中型浮游动物、微型浮游动物。营养盐考虑硝酸盐、铵盐、磷酸盐和硅酸盐。图1是以文献[15]为基础而建立的生态系统概念模型。另外南海属于热带海域,终年平均温度较高,因此选用了浮游生物生长温度修正的Q10法则[4]。
1.3 参数和初、边界条件的确定
1.3.1 参数的确定 模式参数的合理选取对海洋生态系统基本特征的准确模拟非常重要。由于南海北部模式研究还不多,本文中模式参数主要来自ERSEM模拟中的常用参数[17],部分选自南海及邻近海域的研究结果[4]。具体参数取值见表1~2。

表1 模式基本参数及取值Table 1 Basic parameters and value for the model

表2 模式浮游动物参数及取值Table 2 Zooplankton parameters and value for the model
1.3.2 热力强迫和动力强迫 海表面物理强迫主要考虑为风和太阳辐射。海表面平均风速值数据采用NECP 2001—2008年平均风场再分析资料(见图2)。海表面太阳辐射数据通过SBDART(Santa Barbara Disort Atmosphere Radiative Transfer)软件计算得到[18](见图3)。水文条件初值取自南海上层物理海洋学气候图集[19](18°N,116°E);生源要素初值取自南海海洋图集(化学分册)[20](18°N,116°E)。
南海地处东亚季风区[21],季风和太阳辐射是南海水文要素变化的主要驱动力。如图2所示,每年6~9月盛行暖湿的西南季风(夏季风),11月至次年3月盛行干冷的东北季风(冬季风),4~5月及9~10月则为季风转换时期。东北季风的平均风速为10 m.s-1,相对较弱的西南季风的平均风速则为6 m.s-1。如图3所示,南海北部纬度较低,其辐射终年维持在200~360 W.m-2。
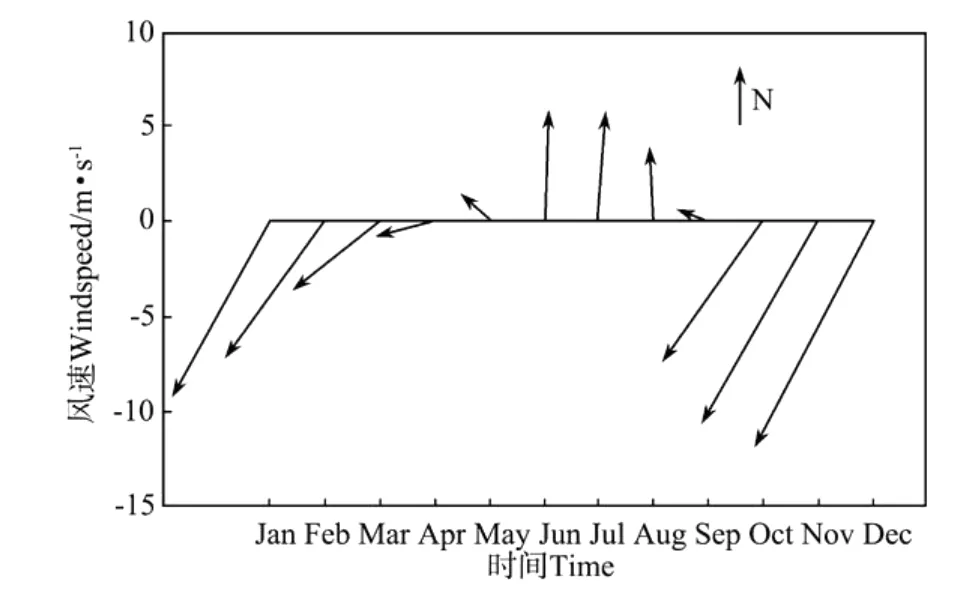
图2 海表面月平均风速、风向随时间的变化Fig.2 Monthly long-term mean of wind speed and direction at the surface
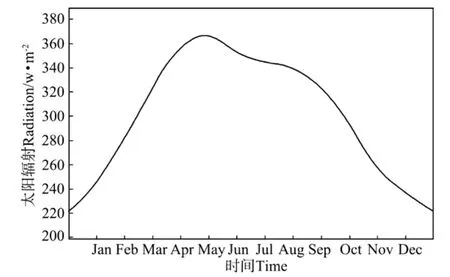
图3 海表面太阳辐射量随时间的变化Fig.3 The monthly solar radiation at the surface
1.3.3 模型配置 本文模拟了南海北部生态要素的季节变化特征,并利用SEATS站资料进行模式结果验证。模式从1月1日开始,运行720 d,时间步长为216 s。模式计算水深1 200 m(南海平均水深),分40层。由于重点关注上层海洋浮游生态系统的季节变化,故水面至水深150 m处采用上密下疏的非均匀分35层,而水深150 m以下均匀分5层。选取第2年模拟结果与实测数据进行比较验证。
2 结果与分析
2.1 模式结果验证
SEATS站是1个设置在副热带边缘海的观测站,从1998年8月运行至今已经完成20多个航次的观测,获得了不同季节水文、化学和生物变量的垂直剖面[22],为模式验证提供了很好的数据基础。
本文选取文献[22]SEA TS站2000年1、5、7、11月的温度、盐度和叶绿素a浓度的观测资料验证模式结果。如图5所示,冬季(11~1月)南海北部受控于东北季风[4],混合层较深,Chla也呈全年最高值(0.38 mg.m-3)。夏季(5~7月)海面风速较小,海水层结稳定,跃层深度大约在20~30 m,Chla浓度出现次表层最大值现象,深度大约为80~100 m。由于模式的风场强迫选取的是多年平均资料,变量初值也是来自图集,使得模拟结果与SEATS站2000年的实测值数据对比存在一定差别(如1月和11月Chla的模拟结果),但整体上来说,模式较好的反映了该站150 m以上各个变量垂向分布的季节变化特征。
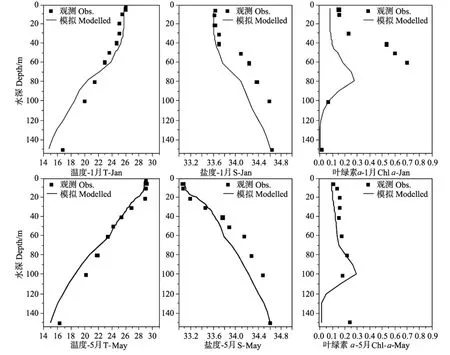
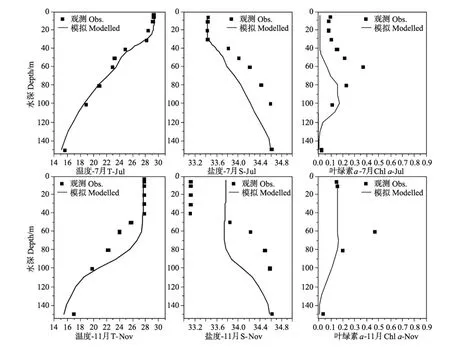
图5 温度、盐度和Chla垂直结构季节变化的模拟和实测资料比较Fig.5 Comparison of modelled vertical seasonal changes of temperature,salinity and chlorophyll-a(mg.m-3)with observation data
2.2 温度和盐度垂直结构的季节变化
模拟得到的温度和盐度垂直结构的年循环如图6所示。南海北部地处亚热带地区,SST(Sea Surface Temperature)终年在24.0~29.2℃之间。3月底,随着太阳辐射的不断增强(见图3),风速逐渐变小(见图2),海水表面开始增温,温跃层变浅。7月SST达到最大值29.2℃。之后,由于太阳辐射的减弱,表层海水温度降低,加之风速的增大使得混合层逐渐加深。12月混合均匀的水层最深可达到100 m。盐度模拟结果显示,受南海北部盛行季风影响,冬夏季风时期海表面盐度升高,8月和12月表层盐度呈现2个高值,分别为33.59,33.60。
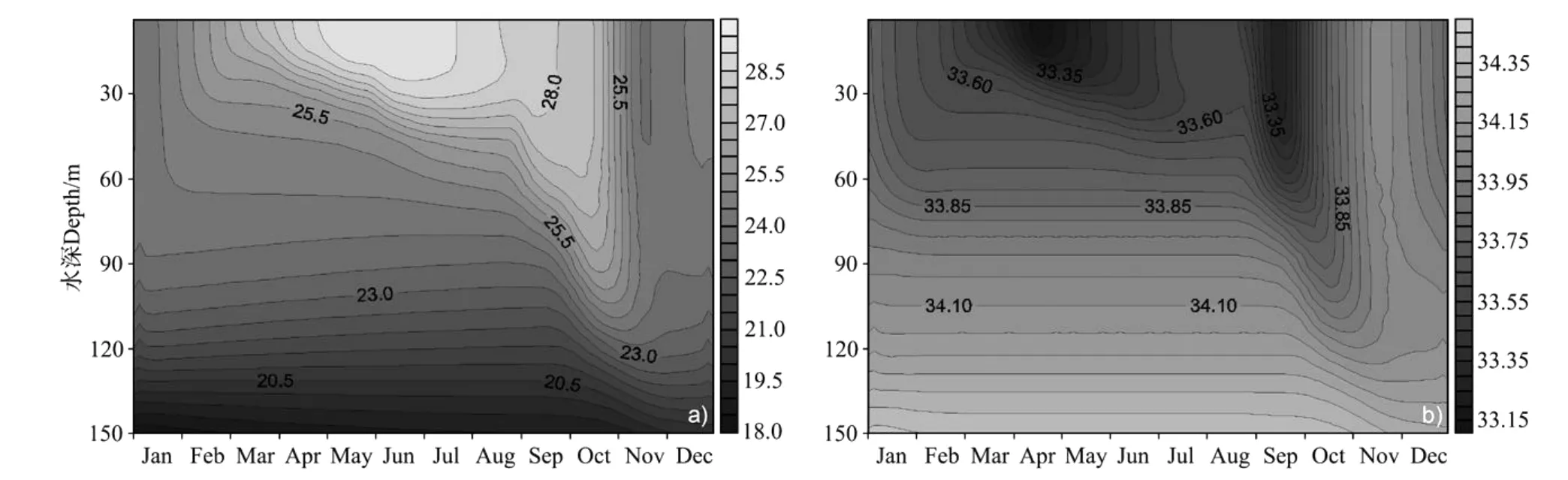
图6 温度、盐度季节变化的模拟结果Fig.6 Simulated seasonal cycles of temperature and salinity
根据温度模拟结果计算的混合层深度的变化如图7。本文定义比10 m以浅平均温度低1℃的位置为混合层深度[23]。模拟结果表明,南海北部混合层终年存在,夏季混合层较浅,4月混合层最浅为15.6 m;南海北部冬季受控东北季风,平均风速达到11.5 m.s-1(见图2),在此作用下,12月混合层深度可达120 m(见图7)。王东晓[19]研究认为南海大部分海区水体层化明显,上混合层的深度通常只有30~50 m,但南海北部浅水陆架区水体混合均匀,混合层厚度可达75 m以上,而混合层的时空变化较大,但很少超过150 m。
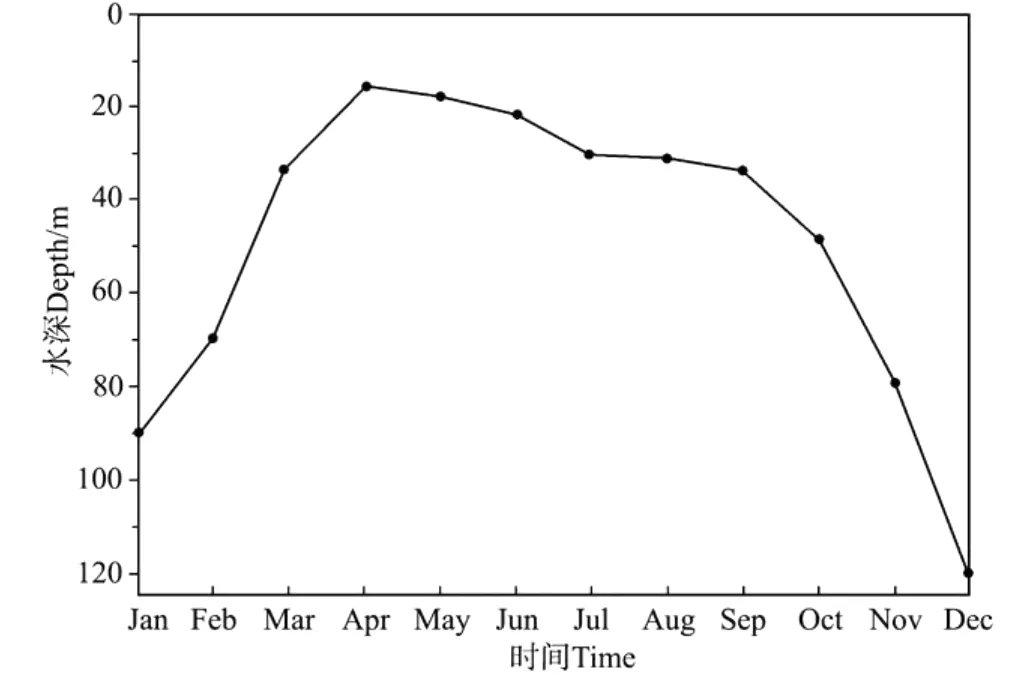
图7 混合层深度变化的模拟结果Fig.7 Simulated seasonal cycle of mixed layer depth
2.3 浮游植物垂直结构的季节变化
南海具有热带开阔大洋寡营养的特征,虽然太阳辐射对浮游植物生长限制较小,但生产力却很低[24]。模拟结果所示,南海北部表层Chla浓度存在明显的季节性变化,夏季呈现低值(<0.05 mg.m-3),冬季呈现高值,12月为0.38 mg.m-3。模拟得到的Chla浓度的季节变化趋势与现场和卫星观测结果相类似[9-10,25]。Chla浓度季节变化呈单峰分布,与我国黄海中部浮游植物生物量的双峰分布特征不同[14]。
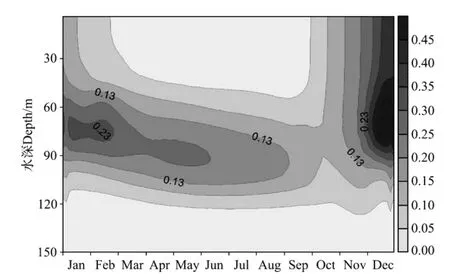
图8 Chla浓度垂直结构季节变化的模拟结果(单位:mg.m-3)Fig.8 Simulated seasonal cycle of chlorophyll-a(mg.m-3)
春季,南海风速相对变小,垂直混合变弱,由于跃层的阻碍作用,下层丰富的营养盐很难到达上层,再加上南海太阳辐射终年较强,海水透明度高,Chla浓度次表层最大值现象得以形成,位置处于温跃层之下(80~100 m)。在寡营养的地区,次表层绿叶素最大值现象(SCM)已有诸多报道[26-31](见表3),虽然表3列举的海区同属于开阔大洋区域,但是SCM发生的深度和强度都不尽相同,原因是不同海区的物理和生物环境都存在差异。Liu等[32]对南海初级生产力模拟研究发现,光适应性(Photo-adaptation)是SCM现象形成的关键因素。这种现象一直持续到10月末,随后由于海面开始降温,同时风向转为东北风,风速增大,垂直混合增强,使下层营养盐向上补充,表层海水浮游植物快速生长,出现水华,且混合层内均维持高值(见图8)。

表3 不同海域次表层叶绿素最大值对比Table 3 SCM in different seas
2.4 营养盐垂直结构的季节变化
图9所示为模拟的硝酸盐、铵盐、磷酸盐和硅酸盐浓度垂直结构的季节变化,其变化趋势与南海图集化学分册[20]中的资料基本吻合。冬季11月~次年1月,海面的降温和东北季风盛行使垂直混合增强,把深层的营养盐带到上层,营养盐垂直梯度减小。真光层中营养盐的补充为浮游植物生长提供了条件,加之南海冬季温度对浮游植物生物生长并不限制,因此出现浮游植物水华现象(见图8)。浮游植物水华过后,表层水体中的硝酸盐、磷酸盐和硅酸盐迅速减少,开始出现营养盐跃层,其跃层位置和浮游植物极值深度一致,大约为80~90 m。春末夏初,浮游生物生长大量消耗上层营养盐,同时,海表面风速降低,温跃层形成,深层营养盐被阻碍在温跃层之下,出现层化现象。
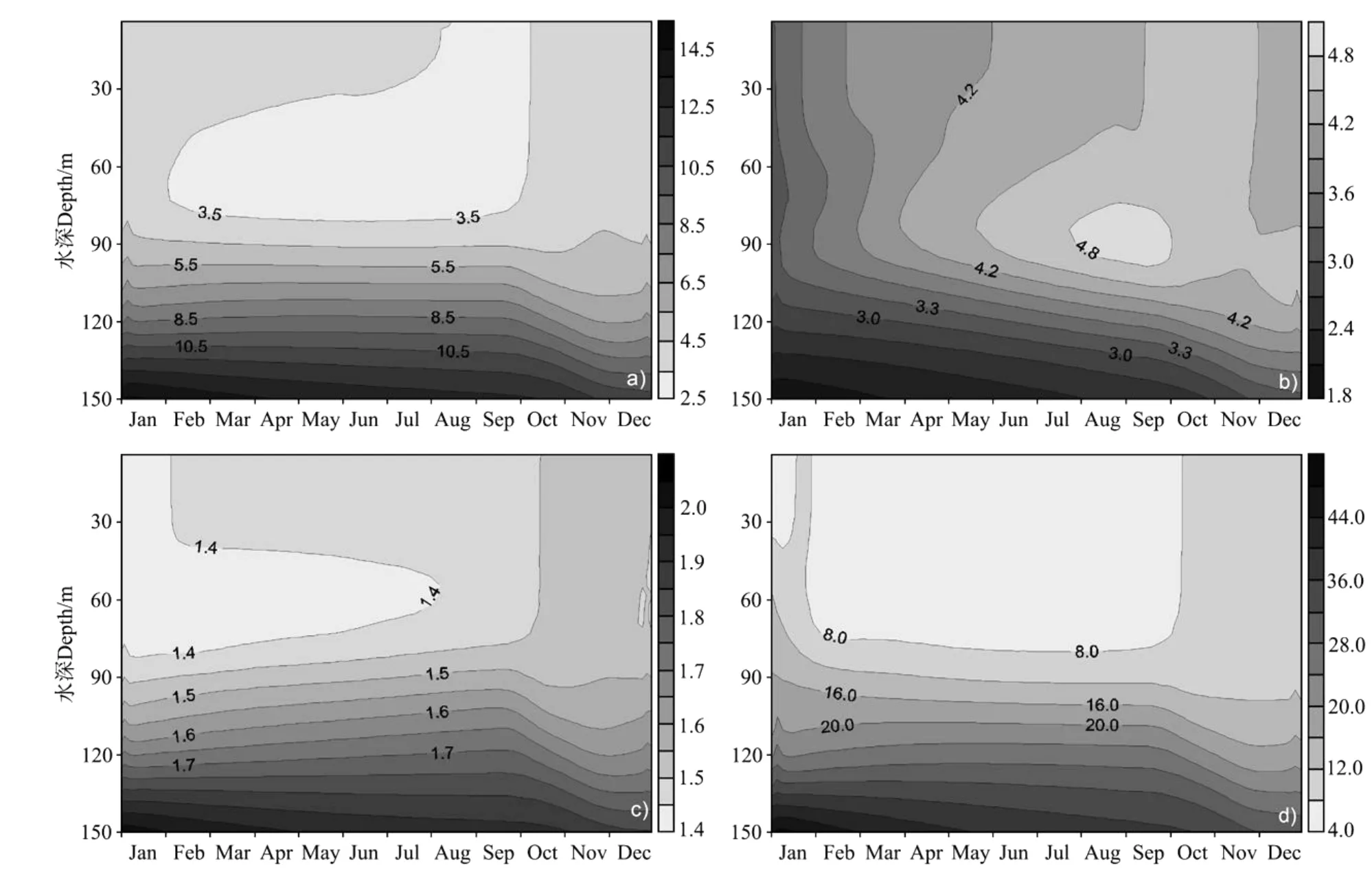
图9 营养盐垂直结构季节变化的模拟结果Fig.9 Simulated seasonal cycle of nutrient
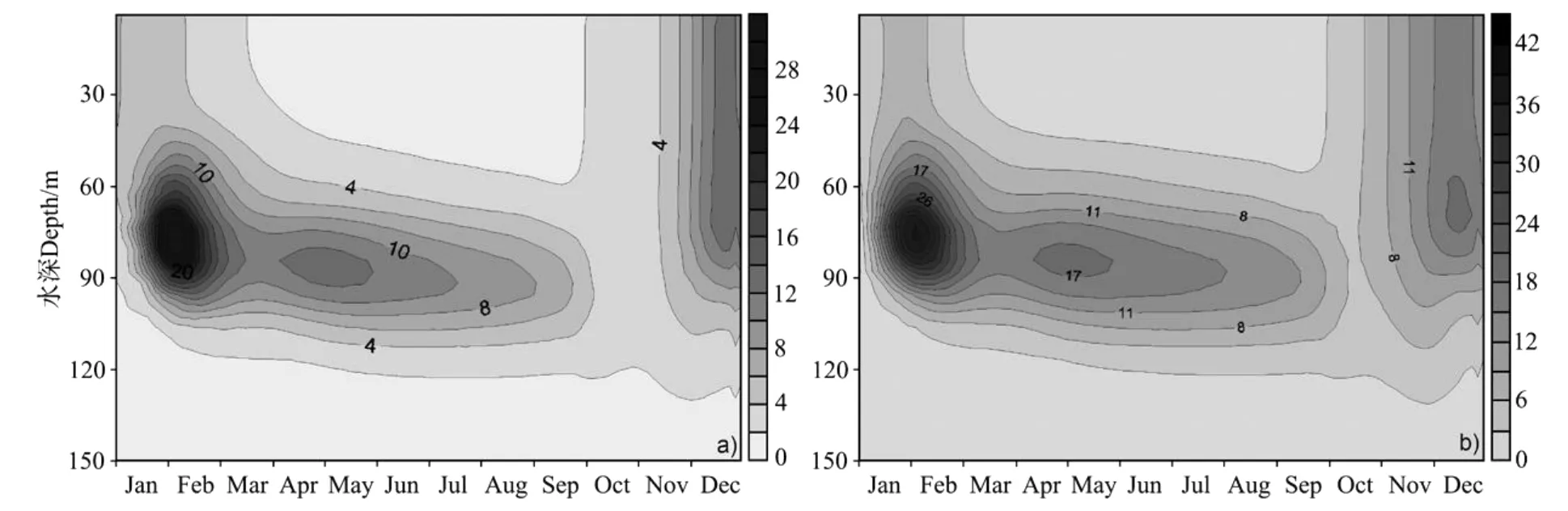
图10 浮游动物和细菌垂直结构季节变化的模拟结果(单位:mg.m-3)Fig.10 Simulated seasonal cycle of zooplankton and bacteria(mg.m-3)
铵盐的垂直分布在4~9月出现了深层最大值现象,这种现象体现了氮的再生过程。浮游植物生长把无机氮转化成有机氮,并通过浮游动物的摄食,及各级动物之间的捕食继续在食物链中传递,在这个过程中浮游植物的胞外溶出、浮游动物的溶出与排泄及细菌对有机碎屑降解的产物主要为铵氮[14],故铵盐浓度升高。另外,冬季水华过后,通过浮游生物和细菌的作用,一部分硝酸氮再生转化为氨氮,使得铵盐的浓度再次增加。
2.5 浮游动物和细菌垂直结构的季节变化
图10a为浮游动物垂直结构的季节变化。浮游植物的现存量除了受上行控制(Bottom up),还受控于下行控制(Top down),即浮游动物对浮游植物的摄食调节作用[33],反之,浮游植物的多寡也会影响浮游动物的浓度变化。在冬季12月底叶绿素a浓度出现峰值后,1月表层浮游动物出现峰值,最大值为13 mg.m-3。3~10月,由于南海表面风速变小,垂直混合减弱,浮游动物浓度高值出现在次表层60~100 m处,其最高值为12 mg.m-3。南海北部浮游动物四季总生物量变化范围[34]为18~38 mg.m-3,与本次模式计算结果大致相当。图10a与图8比较,可以看出浮游动物与Chla浓度的时空分布有很好的一致性,浮游动物的生长滞后于浮游植物。大西洋BATS站(31.5°N,64.1°W)的分析表明,浮游动物与浮游植物的量呈线性关系,其生长滞后于浮游植物[35],与南海北部的结果类似。
细菌和浮游动物的垂直结构相似,冬季上层水体中细菌出现最大值,为18 mg.m-3,且混合趋于均匀。3~10月细菌同浮游动物一样出现次表层最大值(14 mg.m-3),其深度与浮游动物位置相当。
3 结语
本文利用垂直一维海洋生态系统动力学模式(MEM-1D)对南海北部各要素垂直结构的季节变化进行了模拟,并选取南海北部SEATS站(18°N,116°E)实测资料对模拟结果进行验证。虽然模拟结果和实测数据有一定差异,但整体上来说,模式还是较好的反映了该站150 m以上水层中各要素垂向分布特征的季节变化。
南海北部表层叶绿素浓度季节性变化明显,夏季呈现低值(<0.05 mg.m-3),冬季呈现最高值(0.38 mg.m-3)。春季,风速相对变小,垂直混合变弱,下层丰富的营养盐很难到达上层,次表层叶绿素a浓度最大值现象形成(80~100 m),这种现象一直持续到10月末。冬季,海温降低,东北季风暴发,垂直混合增强,使下层营养盐向上输运,表层水华现象发生。浮游动物和细菌的垂直结构相似,冬季上层水体中浮游动物和细菌出现最大值,分别为13 mg.m-3和18 mg.m-3。2~10月细菌同浮游动物一样出现次表层最大值(12 mg.m-3和14 mg.m-3),其深度与浮游动物位置相当。
本文通过模拟研究定量分析了南海北部在风和太阳辐射强迫下,温度、盐度和生态要素的季节变化过程,从而刻画了浮游生态系统各要素垂向分布变化的相关关系,为今后开展南海生态系统的过程研究提供了参考。然而,本文模型中的初始条件和边界条件的时空分辨率较低,模型参数的针对性不强,导致模拟结果存在一些不确定性。
[1] Lin I,Liu W T,Wu C C,et al.New evidence for enhanced ocean primary production triggered by tropical cyclone[J].Geophys Res Lett,2003,30(13):1718.
[2] Lee Chen Y.Spatial and seasonal variations of nitrate-based new production and primary production in the South China Sea[J].Deep Sea Research Part I:Oceanographic Research Papers,2005,52(2):319-340.
[3] Liu G,Chai F.Seasonal and interannual variability of primary and export production in the South China Sea:a three-dimensional physical-biogeochemical model study[J].ICES Journal of Marine Science,2009,66(2):420.
[4] Liu K K,Chao S Y,Shaw P T,et al.Monsoon-forced chlorophyll distribution and primary production in the South China Sea:observations and a numerical study[J].Deep Sea Research Part I:Oceanographic Research Papers,2002,49(8):1387-1412.
[5] Tang D,Kawamura H,Lee M,et al.Seasonal and spatial distribution of chlorophyll-a concentrations and water conditions in the Gulf of Tonkin,South China Sea[J].Remote Sensing of Environment,2003,85(4):475-483.
[6] Lee Chen Y,Chen H.Seasonal dynamics of primary and new production in the northern South China Sea:The significance of river discharge and nutrient advection[J].Deep Sea Research Part I:Oceanographic Research Papers,2006,53(6):971-986.
[7] Wong G T F,Ku T,Mulholland M,et al.The South East Asian time-series study(SEATS)and the biogeochemistry of the South China Sea--An overview[J].Deep Sea Research Part II:Topical Studies in Oceanography,2007,54(14-15):1434-1447.
[8] Tseng C M,Wong G T F,Chou W C,et al.Temporal variations in the carbonate system in the upper layer at the SEATS station[J].Deep Sea Research Part II:Topical Studies in Oceanography,2007,54(14-15):1448-1468.
[9] Chun-Mao Tseng G T F W.A unique seasonal pattern in phytoplankton biomass in low-latitude waters in the South China Sea[J].Geophysical Research Letter,2005,32:L08608.
[10] Liu H,Chang J,Tseng C M,et al.Seasonal variability of picoplankton in the Northern South China Sea at the SEATS station[J].Deep Sea Research Part II:Topical Studies in Oceanography,2007,54(14-15):1602-1616.
[11] 韩君,赵亮,魏皓.近岸海域浮游植物水华动力机制研究进展和展望[J].中国海洋大学学报:自然科学版,2008,38(4):527-532.
[12] Ricci L,Pinardi N,Zavatarelli M,et al.Numerical Modelling of the Mediterranean sapropel S1 ecosystem structure[J].Geolines Journal,2000,11:20-23.
[13] 张书文.黄海冷水团夏季叶绿素垂向分布结构的影响机制[J].海洋与湖沼,2003,34(2):179-186.
[14] 夏洁,高会旺.南黄海东部海域浮游生态系统要素季节变化的模拟研究[J].安全与环境学报,2006,6(4):59-65.
[15] Allen J I,Blackford J C,Radford P J.An 1-D vertically resolved modelling study of the ecosystem dynamics of the middle and southern Adriatic Sea[J].Journal of Marine Systems,1998,18(1-3):265-286.
[16] Ning X,Chai F,Xue H,et al.Physical-biological oceanographic coupling influencing phytoplankton and primary production in the South China Sea[J].J Geophys Res,2004,109:C10005.
[17] Vichi M,Baretta J W,Baretta Bekker J G,et al.European regional seas ecosystem model III:Review of the biogeochemical equations[M].[s.l.]:[s.n],2004.
[18] Liang S.Narrowband to broadband conversions of land surface albedo I-Algorithms[J].Remote Sensing of Environment,2001,76(2):213-238.
[19] 王东晓,杜岩,施平.南海上层物理海洋学气候图集[M].北京:气象出版社,2002.
[20] 国家海洋局第三海洋研究.南海海洋图集(化学分册)[M].北京:海洋出版社,2005.
[21] 李敏.南海海洋叶绿素α时空分布及对季风变动响应的研究[D].北京:中国气象科学研究院,2009.
[22] 陳英傑.南海初級生產力之估算與SEATS測站顆粒態有機質之觀測:應用於南海物理-生地化耦合模式之驗證與改進[D].台湾:国立台湾大学海洋研究所,2004.
[23] 孙成学,刘秦玉,贾英来.南海混合层深度的季节变化及年际变化特征[J].中国海洋大学学报:自然科学版,2007,37(2):197-203.
[24] Chen C,Wang S L,Wang B J,et al.Nutrient budgets for the South China Sea basin[J].Marine Chemistry,2001,75(4):281-300.
[25] 赵辉,齐义泉,王东晓,南海叶绿素浓度季节变化及空间分布特征研究[J].海洋学报.2005,27(4):45-52.
[26] Huisman J,Thi N,Karl D M,et al.Reduced mixing generates oscillations and chaos in the oceanic deep chlorophyll maximum[J].Nature,2006,439(7074):322-325.
[27] Hodges B A,Rudnick D L.Simple models of steady deep maxima in chlorophyll and biomass[J].Deep Sea Research Part I:Oceanographic Research Papers,2004,51(8):999-1015.
[28] Hopkinson B M,Barbeau K A.Interactive influences of iron and light limitation on phytoplankton at subsurface chlorophyll maxima in the eastern North Pacific[J].Limnology and Oceanography,2008,53(4):1303-1318.
[29] Naik H,Naqvi S,Suresh T,et al.Impact of a tropical cyclone on biogeochemistry of the central Arabian Sea[J].Global Biogeochemical Cycles,2008,22(3):1-11.
[30] Pérez V,Fernández E,Maraón E,et al.Vertical distribution of phytoplankton biomass,production and growth in the Atlantic subtropical gyres[J].Deep Sea Research Part I:Oceanographic Research Papers,2006,53(10):1616-1634.
[31] Fennel K,Boss E.Subsurface maxima of phytoplankton and chlorophyll:Steady-state solutions from a simple model[J].Limnology and Oceanography,2003,48(4):1521-1534.
[32] Liu K,Chen Y,Tseng C,et al.The significance of phytoplankton photo-adaptation and benthic-pelagic coupling to primary production in the South China Sea:Observations and numerical investigations[J].Deep Sea Research Part II:Topical Studies in Oceanography,2007,54(14-15):1546-1574.
[33] 乐凤凤,宁修仁.南海北部浮游植物生物量的研究特点及影响因素[J].海洋学研究,2006,24(2):60-69.
[34] 李纯厚,贾晓平,蔡文贵.南海北部浮游动物多样性研究[J].中国水产学报,2004,11(2):139-146.
[35] Madin L P,Horgan E F,Steinberg D K.Zooplankton at the Bermuda Atlantic Time-series Study(BATS)station:diel,seasonal and interannual variation in biomass,1994-1998[J].Deep Sea Research Part II:Topical Studies in Oceanography,2001,48(8-9):2063-2082.
Numerical Study on Seasonal Variations of the Vertical Structures of the Plankton Ecosystem in the Northern Area of South China Sea
ZHANG Chong1,3,SHI Jie1,GAO Hui-Wang1,GAO Zhen-Hui2
(1.Key Laboratory of Marine Environment and Ecology,Ministry of Education,Ocean University of China,Qingdao 266100,China;2.North China Sea Monitoring Center,SOA,Qingdao 266100,China;3.National Marine Envirohmental Monitoring Center,Dalian 116023,China)
One-dimensional Modular Ecosystem Model(MEM-1D)was used to simulate the seasonal variations of the vertical structures of the plankton ecosystem in the northern area of South China Sea.Model results indicate that the concentration of chlorophyll-a at surface presents obviously seasonal variation with the minimum value in summer(<0.05 mg.m-3)and the maximum value in winter(0.38 mg.m-3).In spring,vertical mixing is weaker due to the weaker wind.It is hard for nutrients to reach the upper layer,so the sub-surface chlorophyll-a concentration maximum(SCM)(80~100 m)maintains until the end of October.In winter,the vertical mixing becomes stronger due to the decrease of the sea surface temperature and the strong northeast monsoon.As a result,nutrients can reach the upper layer,which induce the phytoplankton bloom.The vertical structures of the biomasses of zooplankton and bacteria are similar and the maximum values both appear in winter with 13 mg.m-3and 18 mg.m-3respectively.
marine plankton ecosystem;typhoon;chlorophyll-a;bloom;numerical study
P735
A
1672-5174(2011)03-011-08
国际合作项目(2010DFA91350);中国海洋大学青年教师科研专项基金项目(20103031);国家重点基础研究发展计划项目(2010CB428900)资助
2010-05-10;
2010-05-21
张 冲(1984-),男,硕士生,从事海洋生态系统动力学研究。
**通讯作者:E-mail:hwgao@ouc.edu.cn
责任编辑 庞 旻
