运动对成长期骨骼肌神经肌肉接头N乙酰胆碱受体表达的影响
2011-01-02安楠
安 楠
运动对成长期骨骼肌神经肌肉接头N乙酰胆碱受体表达的影响
安 楠
研究目的:通过分析骨骼肌神经肌肉接头(NMJ)N乙酰胆碱受体(nAChR)亚基在成长期(离乳-性成熟)的表达特征,了解NMJ发育过程中的变化及运动对这一过程的影响。研究方法:对3周龄SD雄性大鼠进行运动增强(跑台运动)和运动减弱(尾部悬吊)干预,测定γ-、ε-、δ-nAChR m RNA在3~8周龄的表达水平。结果:1)大鼠腓肠肌γ-、ε-、δ-nAChR的表达水平在出生后3~5周迅速下降,至5周龄时仅有微量的表达;2)悬吊组γ-nAChR在5周龄时表达水平高于对照组;运动组γ-nAChR在8周龄时表达水平低于对照组;3)ε-和δnAChR表达水平在组间无显著性差异。结论:骨骼肌NMJ的发育在出生后早期具有典型的敏感期;运动训练可以促进胚胎型γ-nAChR的消退,限制运动则显著延缓这一过程;成熟后的NMJ具有一定的结构稳定性。
运动;神经肌肉接头;成长期;发育;N乙酰胆碱受体亚基;鼠;动物实验
1 前言
N型乙酰胆碱受体(nicotinic acetylcholine receptor, nAChR)是哺乳动物骨骼肌神经肌肉接头(neuromuscular junction,NMJ)与神经递质乙酰胆碱(acetylcholine,Ach)结合、完成神经肌肉信号传递的重要结构,由5种亚基组成,其中的α亚基是与ACh结合的部位。nAChR在发育的过程中需要完成由胚胎型(α2βγδ)向成熟型(α2βεδ)的转换,由ε亚基替代γ亚基,从而实现功能成熟[1]。γ-ε转换并非是nAChR形成所必须的,但对保持终板结构高度集中很关键[14]。研究发现,啮齿动物的这个过程主要在出生后的几周内完成,与NMJ由多神经支配向单神经支配的转化时间一致,成熟型nAChR集中地表达在已经具有胚胎型nAChR的运动终板上,然后逐渐取代胚胎型nAChR[17]。关于γ-ε亚基置换过程的调节机制,神经源性Agrin、肌特异性酪氨酸激酶Musk以及Rapsyn,已知在其中扮演核心角色[12]。NMJ作为神经突触的一种,其形成、发育和功能完善的过程均与神经肌肉运动功能有关。
关于运动训练是否会改变NMJ突触结构成分,不少学者持否定意见。Herscovich S等人[7]认为,10周耐力运动可以阻止胆碱乙酰转移酶(chloramphnicol acetyltransferase,CAT)及乙酰胆碱酯酶(acetylcholinesterase,ACh E)活性的增龄性降低,但却无法增加nAChR的数量。Lynch G. S.等人[10]也提出,运动后nAChR、AChE和CAT均无变化。也有观点认为,运动神经末梢和NMJ结构成分可以对运动发生适应性变化,耐力训练可以改变NMJ突触后膜nAChR的数目,提出运动对NMJ的影响主要表现在nAChR数量和AChE活性的变化[3],伴随于NMJ传递效率的电生理学变化。Desaulniers P[4]等则发现,16周耐力训练可以使不同肌肉运动终板nAChR的数目增加,从而改变nAChR的密度,这种作用是NMJ对长期耐力训练产生的适应性改变,同时受nAChR代谢状态的调节。另外,对NMJ超微结构的研究表明,小强度长时间的跑台训练可以使突触前活动区密度、突触后膜nAChR密度增加,从而使量子释放增加,并有足够的nAChR与之结合,在高突触功能水平上维持突触传递[16]。以上的研究主要针对运动对成年期NMJ的影响,以及通过NMJ染色和图像分析推断nAChR的变化情况,对于nAChR的分子水平变化以及对运动发育更为敏感的成长期关注较少。
本研究通过测定大鼠腓肠肌中nAChR部分亚基在出生后3~8周的表达情况,判断骨骼肌NMJ发生γ-ε亚基置换的时间,从分子水平研究成长期腓肠肌NMJ结构发育的特征以及运动对这一过程的作用,从而丰富早期发展运动能力的基础理论。
2 研究对象和方法
2.1 研究对象
出生18天的Sprague-Daw ley(SD)雄性大鼠130只,随机分为对照组(Control Group,C组)、运动组(Training Group,T组)和悬吊组(Hanging Group,H组),对照组常规饲养;运动组进行跑台运动,运动负荷从8 m/min、20 min/d开始,运动强度和运动时间交替增加,至8周时达42 m/min、60 min/d,每天运动1次,每周运动6天;悬吊组采用尾部悬吊造成后肢悬空去负荷。对照组在3周龄时取材一次,各组在达4周龄、5周龄、6周龄、8周龄分别取材,按0.3 m l/100 g体重剂量腹腔注射10%水合三氯乙醛溶液麻醉,迅速分离出腓肠肌于预冷的生理盐水中漂洗,滤纸吸干,置液氮速冻待测。
2.2 研究方法
主要仪器和试剂:Takara TP600型梯度PCR仪(日本); SLAN双通道荧光定量PCR仪(上海);JY-SP-B型电泳槽(北京);JY02S型凝胶紫外分析仪(北京);Mylab通用型RNA快速提取试剂盒(美莱博医学科技有限公司,中国); M-M uLV反转录酶:200 U/μl,Fermentas(MBI公司,美国);RNase Inhibitor:40 U/μl,Fermentas(MBI公司,美国)。
引物设计:所有引物均先由GNEBANK查询m RNA序列,用Primer软件进行引物设计,上海生工生物工程有限公司合成(表1)。
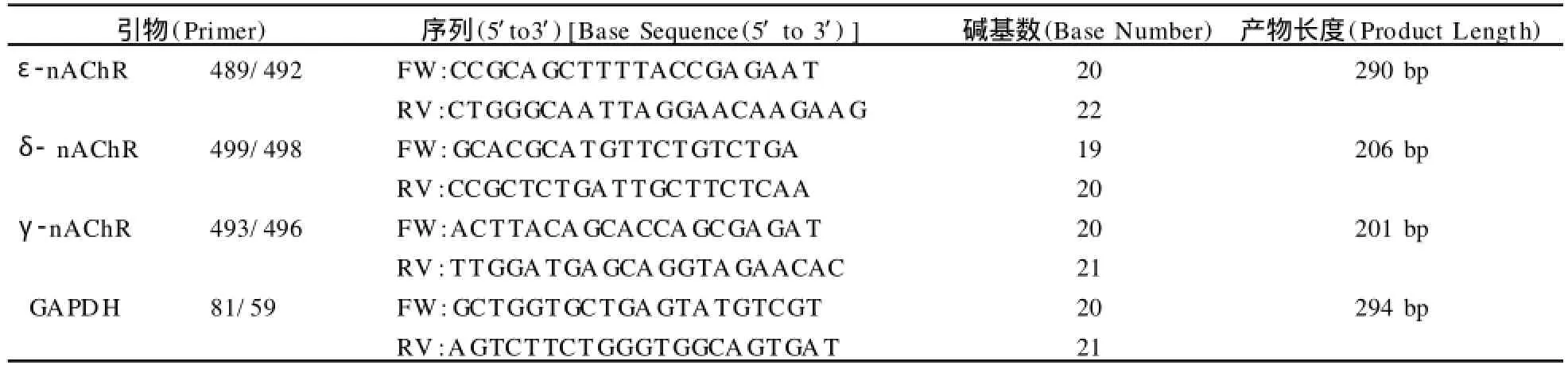
表1 ε-AChR、δ-AChR和γ-AChRm RNA引物序列一览表Table 1 Primers ofε-AChR,δ-AChRandγ-AChRm RNA
组织RNA的提取:从液氮中取出腓肠肌组织约0.1 g,迅速放入液氮预冷的研钵中,加入液氮充分研磨至粉末状;加入1 m l RNA提取液,充分混合,室温放置成溶液,最后加入50μl无RNase的水,溶解RNA。加氯仿0.25 m l,混匀,13 000 rpm离心5 min后,吸取上清750μl,加等体积异丙醇-20℃沉淀核酸,离心取沉淀,75%乙醇洗涤,加DEPC水溶解。用DNase I消化样品RNA中的DNA(表2)。
反转录(25μl反应体系):取RNA样品3μg,加Oligo Dt T18(50M)2.5μl,加DEPC水至25μl,混匀。70℃5 min,立即冰浴,然后在反应体系中依次加入5×buffer 8μl,dNTP(10 mM)4μl,RNase Inhibitor(MBI)1μl,混匀, 37℃5 min;再加入M-MuLV(MBI)2μl,于42℃60 min, 70℃10 min。

表2 DNA消化步骤一览表Table 2 Procedure of DNase digestion
PCR检测:向PCR管中分别加入不含染料2×qPCR TaqMix 12.5μl,10μM各基因正反向引物混合物0.5μl,对应的cDNA各1μl,其中一管不加模板用作阴性对照,各管补加水至25μl。混匀,置于SLAN荧光定量PCR仪中。95℃5 min,55℃15 s,65℃35 s(荧光检测),40循环,获得扩增曲线。
所得实验数据以均数±标准差(±SD)表示;用SPSS 13.0统计软件进行One-way ANOVA比较均值,显著性水平取P<0.05,非常显著性水平取P<0.01。
3 研究结果
图1、图3对γ-、ε-、δ-nAChR亚基在运动干预下同周龄不同组间的表达情况进行了横向比较。

表3 对照组3~8周龄大鼠腓肠肌γ-、ε-、δ-nAChR亚基m RNA相对表达量一览表(n=8)Table 3 Relative expressions ofγ-、ε-、δ-nAChRmRNA in gastrocnem ius of 3~8-week-old SD rats in control group

表4 运动组3~8周龄大鼠腓肠肌γ-、ε-、δ-nAChR亚基m RNA相对表达量一览表(n=8)Table 4 Relative expressions ofγ-、ε-、δ-nAChRmRNA in gastrocnem ius of 3~8-week-old SD rats in train ing group
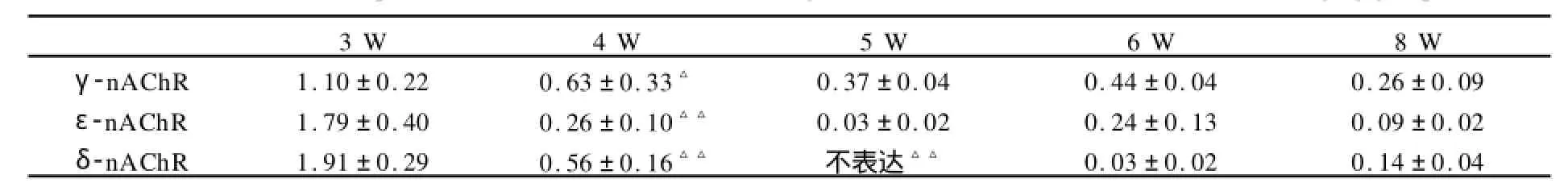
表5 悬吊组3~8周龄大鼠腓肠肌γ-、ε-、δ-nAChR亚基m RNA相对表达量一览表(n=8)Table 5 Relative expressions ofγ-、ε-、δ-nAChRm RNA in gastrocnemius of 3~8-week-old SD rats in hanging group

图1 正常发育及运动干预下3~8周龄大鼠腓肠肌γ-nAChRmRNA表达情况折线图(n=8)Figure 1 Expressions ofγ-nAChRm RNA in gastrocnemius of 3~8-week-old SD rats(compared between groups,n=8)
4 讨论
大鼠出生后3~8周龄处于从断乳到性成熟的成长期,本研究检测了大鼠骨骼肌γ-、ε-和δ-三种nAChR亚基在这一时期的转录水平,结果发现,骨骼肌nAChR在成长期的表达具有一定的规律:γ-、ε-和δ-nAChR亚基在出生后3周时均处于较高的表达水平,之后迅速下降(P< 0.01),至5周龄时基本不表达,三种亚基的下降趋势相似(图1~图3),这与现有的一些研究结果相似[5,9]。nAChR亚基的表达始于胚胎期,作为胚胎型NMJ的特征性成分, γ-亚基的表达开始最早,以保证胚胎期nAChR的大量广泛聚集[8],而ε-亚基则相反,其表达开始于神经支配形成之后,远远晚于其他亚基开始转录的时间,说明其表达是受神经支配诱导的[11]。出生后发生的γ-ε置换过程应该不是一种亚基消失之后另一种亚基才开始出现的替代形式,而是出生时各种亚基包括ε-和γ-都已经在NMJ上大量的表达,然后在NMJ各种调控因子和神经肌肉活动的诱导下,胚胎型γ-nAChR逐渐消失,或者含有γ-nAChR的NMJ发生突触消融,最后留下的是信号传递效率更高的含有成熟型ε-亚基的NMJ。本研究因研究时段的限制,未捕捉到nAChR各亚基表达高峰的出现时间,但推测ε-亚基表达高峰晚于γ-和δ-nAChR。

图2 正常发育及运动干预下3~8周龄大鼠腓肠肌ε-n AChRm RNA表达情况折线图(n=8)Figure 2 Expressions ofε-nAChRm RNA in gastrocnemius of 3~8-week-old SD rats(compared between groups,n=8)
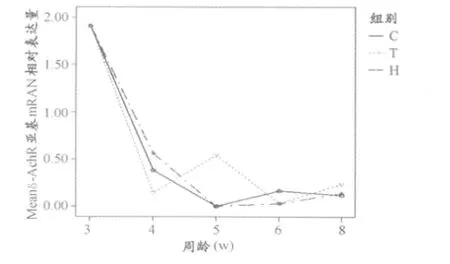
图3 正常发育及运动干预下3~8周龄大鼠腓肠肌δ-nAChR m RNA表达情况折线图(n=8)Figure 3 Expressions ofδ-nAChRm RNA in gastrocnemius of 3~8-week-old SD rats(compared between groups,n=8)
值得注意的是本研究发现γ-亚基表达减少的时间晚于其他两种亚基(P<0.01),发生在4周龄以后,而ε-和δ-亚基的表达在3周龄就开始大量减少,提示大鼠骨骼肌γ-ε亚基置换过程发生在出生后3~4周左右,即断乳之后很短的一段时间。Saito等曾检测大鼠咬肌nAChR的表达,发现该结构的成熟性转化发生在出生后的3周内,与咬肌运动功能开始的时间(大鼠断乳进食的时间为出生后3周左右)基本一致[13]。另一项研究小鼠舌肌和骨骼肌的结果显示,出生时舌肌已完成γ-ε亚基转换,但后肢骨骼肌尚未完成[18]。这可能是由不同部位肌肉的功能需要决定的,出生后即需要吸吮,而下肢运动功能在出生后10天左右才开始发育,骨骼肌的运动功能发育显然比负责吸吮的舌肌和负责咀嚼的咬肌复杂得多,其功能成熟的时间也应与结构完成成熟型转换的时间相一致,因此,可以推断出生后3~4周是大鼠骨骼肌NMJ功能成熟和运动能力进一步发育和完善的重要时期。本研究还发现,γ-nAChR在5周龄以后又出现小量的表达增加,推测可能与青春期发育有关,在此阶段,各种NMJ诱导因子的表达增多,神经肌肉活动增强,NMJ结构进一步发育完善,但这一过程并不会影响NMJ基本功能的稳定。
从运动干预对nAChR三种亚基表达的影响来看,悬吊组γ-亚基表达减少的时间推迟,5周龄时仍显著高于对照组(P<0.01),且在整个3~8周龄阶段持续地表达;运动组γ-亚基水平则显著低于对照组(P<0.01),提示运动可能是骨骼肌NMJ发生γ-ε亚基置换、促进胚胎型γ-亚基消退的诱导因素;这与一项对大鼠咬肌nAChR表达的研究结果相似,该研究发现咬肌运动减少(软食喂养)可以延缓γ-亚基消除的过程[2]。运动增强可能通过增强神经肌肉电活动促进了胚胎型γ-亚基的表达减少,从而加速γ-ε亚基置换的过程。在胚胎型nAChR缺乏的情况下,肌自发性肌电不能发生,提示γ-亚基与NMJ早期形成自发性电活动有关[15]。本研究未发现运动干预对ε-亚基和δnAChR表达具有显著影响。ε-亚基适应于成年的肌肉运动,Schwarz[14]等的研究也认为,ε-亚基的表达似乎对神经肌肉活动的变化不敏感,只受固有的发育程序影响,这种神经肌肉活动不敏感性是使nAChR成熟以后保持相对稳定的重要保障,有利于NMJ基本功能的维持和神经肌肉信号的稳定传递。肌肉失神经支配后nAChR各亚基的表达一般都会增加,γ-亚基可能重新在成年个体表达,ε-亚基则较为特殊,基本上不受失神经支配的影响[6],虽然失神经支配的过程比单纯的运动增强或减弱复杂,但其中也有神经肌肉活动减弱的过程,ε-亚基不受其影响也从另一个角度说明其对运动干预不敏感,从而使NMJ成熟后能够保持相对的稳定。
5 结论
1.出生后骨骼肌NMJ中γ-、ε-、δ-nAChR亚基的表达在一定时期内逐渐减少,γ-亚基的表达减少晚于其他两种亚基,γ-ε亚基置换的时间是运动功能发育的敏感期;青春期nAChR亚基再次表达增加。
2.运动干预可以在一定程度上影响骨骼肌NMJ的发育,运动训练可以促进骨骼肌胚胎型γ-亚基的消退和γ-ε置换,运动不足则延缓这个过程;但运动对成熟型ε-和δ-亚基的表达无显著影响,因此不会改变神经肌肉信号传递的基本稳定性。
[1]蔡文琴.发育神经生物学[M].北京:科学出版社,2007:240-241.
[2]马军利,刘洪臣,王冠超.软食喂养对发育中大鼠咬肌乙酰胆碱受体亚基mRNA表达的影响[J].临床口腔医学杂志,2008,24 (8):473-475.
[3]ALSHUA IBW B,FAH IM M A.Effectof exercise on physiological age-related change at mouse neuromuscular junctions[J]. Neurobiol Ageing,1990,11(5):555-561.
[4]DESAULN IERS P,LAVOIE P A,GARD INER P F.Endurance training increases acetylcholine receptor quantity at neuromuscular junctions of adult rat skeletal m uscle[J].Neu ro repo rt,1998, 9(16):3549-3552.
[5]FRA TERMAN S,KHURANA T S,RUBINSTEIN N A.Identification of acetylcholine receptor subunits differentially expressed in singly and multip ly innervated fibers of extraaocular muscles[J].Invest Ophthalmol Vis Sci,2006,47(9):3828-3834.
[6]GHED IN IP C,V IEL T A,HONDA L.Increased exp ression of acetylcholine receptors in the diaphragm muscle of MDX mice [J].M uscle Nerve,2008,38(6):1585-1594.
[7]HERSCOV ICH S,GERSHON D.Effects of aging and physical training on the neuromuscular junction of themouse[J].Gerontology,1987,33(1):7-13.
[8]H R BRENNER,S ROTZLER,W A KUES.Nerve-dependent induction of AChR epsilon-subunit gene exp ression in muscle is independent of state of differentiation[J].Dev Biol,1994,165: 527-536.
[9]L IU Y,PADGETT D,TA KA HASH IM.Essential roles of the acetylcholine receptor gamma-subunit in neuromuscular synaptic patterning[J].Development Epub,2008,5(11):1957-1967.
[10]L YNCH G S,RODGERSB J,W ILL IAMSD A.The effects of age and low-intensity endurance exercise on the contractile p roperties of single skinned fast-and slow-twich skeletalmuscle fibers[J].Grow th Dev Aging,1993,57(3):147-161.
[11]NORIH IRO YUMOTO,ShUJIWA KAUSU KI,A TSU KO SEHARA-FUJISAWA.The acetylcholine recep to r c-to-e sw itch occurs in individual endp lates[J].Biochemical Biophysical Res Communications,2005,331:1522-1527.
[12]RW ILL IMANN,C FUHRER.Neuromuscular synaptogenesis: clustering of acetylcholine recep tors revisited[J].Cell Mol Life Sci,2002,59:1296-1316.
[13]SA ITO T,OHNUKI Y,SAEKI Y.Postnatal changes in the nicotinic acetylcholine receptor subunits in ratmasseter muscle [J].A rch O ral Bio l,2002,47(5):417-421.
[14]SCHWARZ H,GIESE G,M ULLER H.Different functions of fetal and adult AchR subtypes for the formation and maintenance of neuromuscular synapses revealed inε-subunit deficient mice[J].Eur J Neurosci,2000,12(9):3107-3116.
[15]TA KAHASH I M,KUBO T,M IZOGUCH I A.Spontaneous muscle action potentials fail to develop without fetal-type acetylcholine recepto rs[J].EMBO Rep,2002,3(7):674-681.
[16]TOMASJ,SAN TAFEM,LANUZA M A.Physiologicalactivities-dependent ultratructural plasticity in normal adult rat neuromuscular junctions[J].Biol Cell,1997,89(1):19-28.
[17]W A KUES,B SAKMANN,V W ITZENMANN.Differential expression patternsof fiveacetylcholine receptor subunit genes in rat muscle during development[J].Eur J Neurosci,1995, (7):1376-1385.
[18]YAMANE A,OHNUKI Y,SAEKI Y.Developmental changes in the nico tinic acetylcholine recep to r in mouse tongue striated m uscle[J],J Dent Res,2001,80(9):1840-1844.
Effects of Activities on Expression of Nicotin ic Acetylcholine Receptor on Neuromuscular Junction of Skeletal M uscles in Growth Stage
AN Nan
Objective:To describe the development p rocess of skeletal neuromuscular junction (NMJ)in grow th stage as well as the effects of activities in this p rocess through the analysis of N acetylcho line recep to r(nAChR)subunits exp ressions.M ethods:3-week-o ld male SD rats have undergone activity increasing(treadmill training,group T)and activity decreasing(tail suspension,group H)intervention respectively.The exp ressions ofγ-,ε-,δ-nAchR m RNA w ere detected during 3~8 w eeks after birth.Results:1)γ-,ε-,δ-nAChR exp ressions in rat gastrocnemius decreased rapidly in 3~5 weeks after birth and only p resented trace amounts of exp ression at 5-w eek-old stage;2)the exp ression ofγ-nAChR was higher in group H at 5-w eek-old stage and low er in group T at 8-w eek-old stage compared w ith control group.There w ere no significant differences ofε-andδ-nAChR exp ressions between groups.Conclusion: The development of NMJ in postnatal stage has a typical sensitive period;exercise training can promote the recession of embryonicγ-nAChR,restriction on movements can significantly delay this p rocess instead.The NMJ has certain structural stability after getting mature.
NM J;grow th stage;development;nAChR subunit
G804.7
A
1000-677X(2011)11-0056-05
2011-09-19;
2011-10-25
国家体育总局体育科学研究所基本科研业务费课题(2006-17)的部分研究成果。
安楠(1974-),女,河北秦皇岛人,助理研究员,博士,研究方向为发展运动能力的理论与实践,Tel:(010) 87182522,E-mail:cfaannan@yahoo.com.cn。
国家体育总局体育科学研究所,北京100061 China Institute of Sport Science,Beijing 100061,China.
