半干旱区四种典型豆科牧草群落生理生态的分异研究*
2010-12-21左胜鹏王会梅李凤民山仑
左胜鹏,王会梅,李凤民,山仑
(1.安徽师范大学环境科学与工程学院,安徽芜湖241000;2.中国科学院 水利部水土保持研究所黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌712100)
黄土高原半干旱地区是一个农、林、牧综合发展的区域。长期以小麦等粮食作物为主,产业结构单一,生产力水平低下是当地农业存在的现实问题[1]。实施草田轮作,增加豆科牧草的种植比例,通过豆科牧草的生物固氮,增加土壤含氮量和有机质含量,改善土壤质量,有利于提高作物生产力和改善系统稳定性[2]。苜蓿(Medicago sativa)是我国人工栽培最早,分布面积最大的牧草之一,在我国黄土高原已有千年的种植历史,已成为我国黄土高原新产业带一个重要组成组分[3]。沙打旺(Astragalus adsurgens)作为一种优良的豆科牧草和绿肥植物,在我国黄河故道地区栽培也已有近百年的历史。由于它具有适应性强,抗旱、耐寒、耐盐碱、抗风沙的特性,20世纪70年代初先后在我国华北、西北、东北等地引种成功后,人们对其进行了广泛的研究,取得了不少有效的结果[4]。
红豆草(Onobrychisviciaef olia)是一种多年生豆科植物,邓振镛等对红豆草的生长特性作了系统分析,指出盛花期生产力最高[5]。胡枝子(Lespedeza dahurica)是本地天然群落中的主要伴生种之一,分布广泛,属豆科半灌木饲草,其生态适应性强,生长发育快,营养价值高,适口性好,是各种牲畜喜食的饲草之一,并具有良好的水土保持功效[6]。豆科牧草在黄土高原半干旱地区土地利用与可持续发展中将会发挥越来越重要的作用[7]。在不同生态环境和不同生产条件下如何选择与配置不同的豆科牧草,对建立合理的土地利用结构和产业结构具有重要意义。本文通过研究比较了4种豆科饲草植物水分利用与生产力形成的种群生态条件,探讨了其中的网络耦联机制,试图为合理利用豆科牧草,建立适宜的草田轮作系统和退耕还草中适宜的草种选择提供科学依据。
1 样地概况
试验在中国科学院安塞试验站进行。安塞站地处黄土高原中部,区内沟壑纵横,坡陡沟深,属典型的梁峁状丘陵沟壑区。海拔高度1 080 m,年平均温度8.8℃,最冷月为1月,月平均温度-6.9℃,最热月为7月,月平均温度22.6℃,全年≥10.0℃的积温为3 113.9℃,无霜期为159 d。年平均降雨量541.2 mm,其中7-9月占全年降水量的60%~80%,且多暴雨。平均水面蒸发量1 800~2 200 mm,为降水量的3.7倍,气候干燥度1.14,属暖温带半干旱区,森林草原地带,土壤类型主要为黄绵土[8]。
2 材料和方法
本研究的4种牧草样地均于1995年7月播种,川地、旱作、无灌溉和地表来水。每样地4×10 m2,行距70 cm,海拔高度1 070 m。测定于2004年8月下旬到9月初牧草抽穗期进行。
在4个样地中分别用目测法估算草群盖度,并随机选10株测定植株高度;然后在每个样地中设置1 m×1 m样方各3个,在各样方内齐地刈割植物地上部,把4种豆科植物和其它杂草及枯落物分开,称其鲜重,然后带回室内,在105℃下杀青10 min后,放入70℃恒温下经24 h烘干,称重。
地下部生物量测定选用直径为9 cm土钻,在每个1 m2样地中取地上生物量的地方,进行“W”形取5个样点打钻,分五层取样,即 0-20 cm、20-40 cm 、40-60 cm 、60-80 cm 、80-100 cm,所取根样用水冲洗干净,装入纸袋,放入烘箱中105℃杀青10 min,70℃恒温干燥24 h直到恒重,称取干重,最终转化为1 m2样地内的地下生物量干物质重。
称重法测定土壤水分。每 10 cm一层,深至150 cm,3次重复。
数据统计均采用 SAS 8.1软件进行分析和5%、1%水平上的显著性检验以及相关性分析。
3 结果与分析
3.1 土壤水分的比较
4种草地在60 cm土层以上土壤含水量均较高,差异性较小,而在60 cm以下土壤含水量迅速下降,不同草地差异显著增大,但4类草地总体随土层深度土壤水分呈逐渐下降趋势(图1)。其中胡枝子和红豆草样地土壤水分相对比较相近,80 cm以上土壤含水量保持稳定,这两草地含水量基本没有差异,80 cm以下胡枝子草地土壤水分仍然保持相对稳定,而红豆草草地在120 cm以下含水量迅速下降,显著低于胡枝子草地。沙打旺和苜蓿草地土壤水分接近,在40 cm以下就显著低于红豆草和胡枝子草地,在80 cm以下土壤含水量则迅速下降,在130 cm以下苜蓿草地土壤水分显著低于沙打旺样地。总的来看,除胡枝子样地外,其它3种草地土壤表层(0-60 cm)水分要明显多于下层(60-150 cm)。在下层土壤水分之间存在明显的分异,土壤水分从高到底的顺序为:胡枝子>红豆草>沙打旺>苜蓿。苜蓿、沙打旺和红豆草均表现出明显的下层土壤干旱、上层土壤湿润的典型下伏干旱特征。样地整个土壤剖面平均湿度苜蓿为12.3%,沙打旺为12.5%,红豆草为15.3%,胡枝子为16.8%。
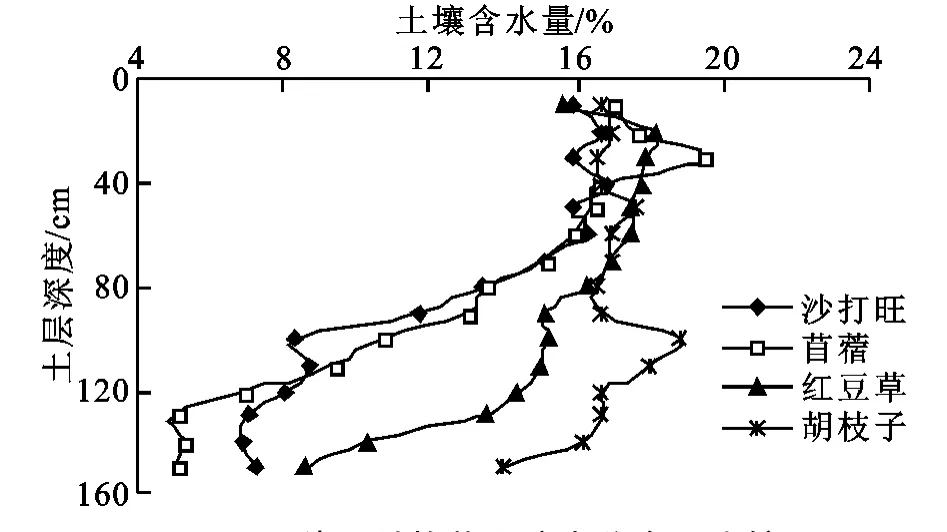
图1 4种豆科牧草土壤水分含量比较
3.2 地上部生长特征
从表1可看出4种豆科牧草地上部生长存在明显差异。苜蓿和胡枝子样地覆盖度无显著差异,但显著高于红豆草和沙打旺样地(p<0.01)。而红豆草覆盖度显著高于沙打旺样地(p<0.05)。苜蓿和红豆草株高无显著差异,二者显著低于沙打旺和胡枝子,沙打旺植株最高,并显著高于其它3种样地。沙打旺和苜蓿两豆科牧草地上生物量差异不显著,但它们显著高于红豆草和胡枝子,前者为后者的两倍多,净生产力比较为:沙打旺>苜蓿>胡枝子>红豆草。从单位面积(1 m2)取样的总生物量看,4类豆科牧草总生物量之间差异不显著,总体比较与净生产力相似,只是红豆草稍高于胡枝子。4类牧草种群中均存在伴生杂草,并且其生物量在种群间差异显著。杂草生物量在红豆草样地中最大,其次为胡枝子和沙打旺,最小的是苜蓿,其中红豆草与沙打旺和苜蓿之间杂草差异达到显著水平。豆科牧草种群均存在显著的更新效应,枯落物之间差异显著,从更新率看,红豆草(0.56)>胡枝子(0.45)>沙打旺(0.32)>苜蓿(0.26)。
在调查的4类种群中杂草生物量占总地上生物量的比值为9.0%~60%,其中苜蓿样地最低,为8.9%,其次是沙打旺草地,占10.5%,而胡枝子和红豆草 2样地杂草占总生物量的比值分别高达32.7%和58.9%。同时发现,伴生杂草与牧草地上生物量存在消长关系,牧草地上生物量高的其伴生杂草相对比较少。伴生杂草与牧草地上生物量的比值发现:红豆草(2.2)>胡枝子(0.7)>沙打旺(0.2)>苜蓿(0.1)。相关分析发现,这4类豆科牧草地上生物量与伴生杂草量、地上生物量与枯落物量、杂草量与枯落物量以及覆盖度与地上生物量均存在明显的相关性,相关系数分别为0.94(p<0.001)、0.73(p<0.01),0.66(p<0.05)和0.50(0.05<p<0.01)。
3.3 地下部生物量与根冠比
4类豆科牧草草地地下生物量调查发现其垂直分布存在明显差异,但均呈现随土壤深度生物量下降趋势(图2)。苜蓿草地0-100 cm各层地下生物量均显著高于其它样地,随土壤深度的增加几乎线性减少,0-40 cm土层地下生物量占0-100 cm土层总根量的69.5%。而沙打旺、红豆草和胡枝子草地0-40 cm土层地下生物量分别为0-100 cm地下生物量的80.2%、80.5%和73.9%。0-40 cm土层的根量从高到低的顺序为:苜蓿>红豆草>沙打旺>胡枝子,后3类样地在40 cm以下根生物量变化很小。这4类样地在0-100 cm土层中的根生物量从高到低分别为:苜蓿样地2 000 g/m2,红豆草805.15 g/m2,沙打旺480.1 g/m2和胡枝子382.84 g/m2。对沙打旺、苜蓿、胡枝子和红豆草4样地的根量与土层深度进行二次曲线回归发现,各样地根系生长与深度有明显的相关性(p<0.05),相关系数分别为0.961,0.999,0.948,0.957。苜蓿样地根冠比最大,可达2.59,红豆草的根冠比略大于1(1.09),胡枝子和沙大旺根冠比分别为0.62和0.53。

表1 不同草地地上部的生长特征
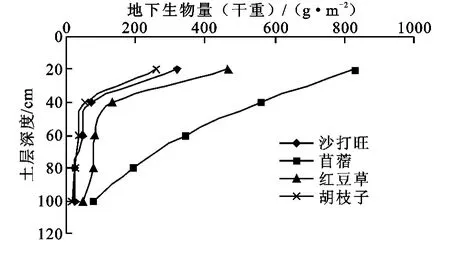
图2 不同牧草样地根系生物量
3.4 地上生物量与土壤水分的关系
地上生物量与90-100 cm土层水分含量呈显著正相关(p<0.05),与l0-80 cm土层水分和110-150 cm土层水分相关性不显著(p>0.05)。土层0-40 cm相关系数明显小于40 cm以下的相关系数。而地下生物量也与土壤水分显示出显著的的曲线相关性(图3,p<0.05),沙打旺、苜蓿和红豆草草地地下生物量与土壤水分含量曲线为U型,胡枝子草地曲线为∩型,在调查的土壤水分范围内,前三类牧草根系发育与土壤水分基本为显著正相关,而且根系生长的土壤水分阈值为12.5%、13.2%和16.3%,而胡枝子根系发育基本稳定在16.6%~18.9%的土壤水分。沙打旺草地中水分含量为9.37%~12.89%时,沙打旺的根系生长受到抑制;土壤含水量为11.87%时,苜蓿草地的地下生物量最低;土壤含水量为15.78%~16.59%时,同样红豆草的根系生长被严重抑制;胡枝子根系在土壤含水量17.68%时生长量最大,低于或高于此值都不利于根系生长。
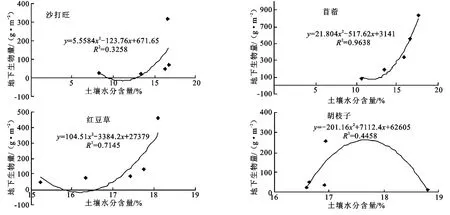
图3 地下生物量与土壤含水量的关系
4 讨论
4种豆科牧草种群生态中的土壤水分环境均表现明显区域滞性,从表层到深层土壤含水量有递减趋势,与Liu等研究人工柠条林土壤水分含量和垂直分布规律基本一致[9],但深度上的变化动态不尽重合,可能四物种对水分资源利用与保持有关系,如根系的发育深度与获取水分能力显著相关,还有根系形态与物化特性对水土保持影响较大[10]。本研究中发现,除胡枝子外,试验区不同样地各层土壤水分变化均近似稳态随机过程,土壤水分表现为胡枝子>红豆草>沙打旺>苜蓿,说明了苜蓿和沙打旺水分利用率较高,适宜在干旱半干旱地区生长,但容易形成土壤干层,与Xu等研究结果基本一致[11],而胡枝子和红豆草由于较小的蒸腾作用和叶表面气孔较小,水分散失也少,适合改良土壤水分环境,可能的解释为全球变化的情况下,当二氧化碳浓度升高时,导致气孔开启的程度变小,以减少植物水分的损失,从而使更多的水留在地表[12]。

表2 单位面积生产力与各层土壤水分含量的相关性
基于地上部生物量的调查可以看出四类豆科牧草的生产力差异显著,这不仅与它们水分利用状况有关,还可能与牧草之间的固氮或氮沉积的氮汇与氮源之间的转化调节有关[13]。从4种豆科牧草的覆盖度和株高来看,叶面积指数和冠层的差异性导致不同物种对光能的利用程度也不一样,从而导致这些物种的净初级生产力的差异[14],如调查中发现沙打旺的覆盖度和株高较其它三者大,可能间接地解释了它的生产力较高。沙打旺与苜蓿的高生产力导致两者的牧草更新也快,产生的枯落物量也大。在这四类牧草的伴生杂草看,杂草与牧草存在消长关系,沙打旺与苜蓿地上生物量大,而杂草比例就相对小;而红豆草与胡枝子生产力低,其伴生杂草相对比较多,可能是牧草与杂草两者的种间竞争所决定,或牧草活体根系释放和相应枯落物腐解产生的化感物质导致对杂草的化感效应等机制所驱动种群格局变化[15]。
沙打旺、苜蓿、胡枝子和红豆草四种牧草在黄土高原丘陵区根系生物量的地下分配具有明显的垂直结构,呈倒金字塔形,上宽下窄,均表现为从土壤表层到深层有逐渐下降的趋势,而且与水分分布特点紧密相关,与Xu等研究干旱气候下白羊草群落地下部生长结果基本一致[16],说明水分生态因素对牧草地下生物量的形成有重要影响。在0-100 cm深度下,苜蓿地下生物量最大达到约2 000 g/m2,分别是沙打旺(480.11 g/m2)、红豆草(805.15 g/m2)和胡枝子(382.84 g/m2)的4.16倍、2.48倍和5.22倍,说明苜蓿对土壤浅层的水分利用较大,容易短期内形成高的根系生物量,立地生长适应性效果显著。一般土壤表层0-40 cm地下生物量基本决定了总地下生物量,可能豆科牧草的根表层结瘤导致地下生物量的表层聚集和根系沉积[17]。在相关分析基础上,发现土壤水分、土壤深度和根系量存在显著相关性,如苜蓿植株生物量表现最高,可能较高生产力与其经过多年生长具有庞大的植株根系有关,这有利于吸收土壤中的深层水分[18],这样地上部分和地下部分形成一种相互促进关系[11],从而在干旱条件下建立一种新的适应性生存对策[19]。
一般植物的地上部和地下部作为植物体最基本的组成部分,共同有机地完成植物体的整体功能,特别在水分亏缺条件下植物显示出明显的整体抗旱性[20]。研究认为,植物的根部在遭受土壤干旱时,地上部会受到伤害,在伤害之前,植物会做出一些适应性反应,追逐有限的供水,最终影响到干物质的积累与在植物不同部位的分配[21]。本研究得出4种牧草的根冠比苜蓿>红豆草>胡枝子>沙打旺,说明这类豆科牧草存在对干旱胁迫根冠均不敏感,根敏感到冠敏感的种间变异差异,如苜蓿为冠部敏感型,沙打旺和胡枝子为根敏感型,而红豆草为根冠均不敏感型。这其中可能原因苜蓿物种在土壤水分减少的情况下,根系到处延伸,追逐水源,根冠竞争碳水化合物,而为了避免水分胁迫,同化物向根系分配较多,促进根系生长,使得根冠比增大[22]。但根冠关系对环境条件的响应是以遗传特性为基础的,不同类型植物由于遗传特性不同,其对环境条件变化的响应也不同。在田间,土壤条件对根系的影响要比遗传特性大得多,水分条件变化也常常是导致根冠生长差异的主要原因[23]。在农业生产中表现为通过耕作栽培措施等调整植物根冠关系以增强植物的抗旱适应能力。在冬小麦进行返青期根冠分配调节后发现,小麦的表层根量和根呼吸消耗降低,对产量没有显著影响,土壤水分消耗相对减少,提高了水分利用效率,达到了节水增效和水分高效利用的目的[24]。
黄土高原土壤水分条件的优劣是反映植物生产力的重要标志。在黄土高原地区分布的草原带的梁、峁、坡等水分较差的地段,特别容易形成“小老树”。这主要违背了“适地适树(草)”原则,造成植物生长需水与土壤供水之间的矛盾加剧,以至于植物生长受阻。从生态适应性的角度来看,这可能由于植株生长力弱,蒸腾失水量不大,以至于土壤水分亏缺程度不致于加剧,从而达到了一种土壤水分稳态[25]。本研究也发现在黄土高原半干旱地区植被生产力的高低与土壤水分状况直接相关(表2)。从4种豆科牧草土壤含水量与地上生物量相关性分析发现,90-100 cm土层土壤储水量与生物量关系显著,可能是由于在该层 CaCO3淀积而形成淀积层[26],而且40 cm以下土层相关系数大于40 cm以上土层相关系数,从生物量与各土层含水量相关性分析,土层越深,生物量与土壤水分含量关系越密切[27],其原因可能是50 cm以上土层的土壤水分受到地面强烈蒸发和土壤无效水的影响。在水分和生产力的关系中植物根系生长对水分的响应意义重大。根的生长决定着水分的供应,间接影响到植物的生长,同时根系也做为光合作用的一个巨大的库而存在。因此对植物生产力与水分关系的研究与根冠整体有关[28]。
在干旱、半干旱地区,不同类型植物生产力在整体上是随着年降雨量的减少而降低,水分不足是限制不同植物生产力发挥的主要限制因子[29]。通过4种豆科植物群落类型中生态分析表明由于豆科牧草的生产力稳定性、物种覆盖度以及土壤水分含量高均显示其植物优势地位优越,说明采取退耕恢复天然植被对增加区域物种多样性和维持系统生产力稳定性是可能的[30]。在半干旱地区,复杂的地形地貌加之降雨的时空分布不均导致不同立地条件微环境多种多样,同时草灌植物为多年生植物,各年的气候环境条件也不同[31]。因此,本研究只能根据调查大致反映8月份陕北地区同一地域条件下不同豆科植物生产力和土壤水分状况。对于4种豆科牧草的长期适应性和持续性还难以定论,特别是针对全球变化下温度和水分的不均一,不同立地条件下4种豆科植物的生长和生产力状况需要准确评价,并且建设长期定位或半定位研究点进行跟踪测定,为该地区大范围植被建设提供依据和参考。
[1] Lu C H,van Ittersum M K,Rabbinge R.Quantitative assessment of resource-use efficient cropping systems:a casestudy for Ansai in the Loess Plateau of China[J].European Journal of Agronomy,2003,19(2):311-326.
[2] Ashraf M,Mahmood T,Azam F,et al.Comparative effects of applying leguminous and non-leguminous green manures and inorganic N on biomass yield and nitrogen uptake in flooded rice(Oryza sativa L.)[J].Biology and Fertility of Soils,2004,40(2):147-152.
[3] Liu W Z,Hu M J,Li F M,et al.Ecological characteristics of soil water and its relations to landform andvegetation in a small semiarid watershed in aloess hilly area of China[J].The International Journal of Sustainable Development and World Ecology,2005,12(3):326-333.
[4] Kita F,Hongo A,Iwashita F.Cytology of wild and cultivated forms and inter-regional hybrids of Astragalus adsurgens Pall.f rom China and Japan[J].SABRAO Journal of Breeding and Genetics,1998,30(1):35-44.
[5] Tufenkc S,Erman M,Sonmez F.Effect of phosphorus and nitrogen application and Rhizobium inoculation onyield and nutrient uptakein sainfoin(Onobrychissativa)[J].Indian Journal of Agricultural Sciences,2005,75(5):261-264.
[6] Du F,Shao H B,Shan L,et al.Secondary succession and its effects on soil moisture and nutrition in abandoned old-fields of hilly region of Loess Plateau,China[J].Colloids and Surfaces B,Biointerfaces,2007,58(2):278-285.
[7] Shangguan Z P,Zheng S X.Ecological properties of soil water and effects on forest vegetation in the Loess Plateau[J].The International Journal of Sustainable Development and World Ecology,2006,13(4):307-314.
[8] Yang M Y,Tian J L,Liu PL.Investigating the spatial distribution of soil erosion and deposition in asmall catchment on the Loess Plateau of China,using137Cs[J].Soil&Tillage Research,2006,87(2):186-193.
[9] Liu W Z,Hu M J,Li F M,et al.Ecological characteristics of soil water and its relations to landform and vegetation in a small semiarid watershed in aloess hilly areaof China[J].The International Journal of Sustainable Development and World Ecology,2005,12(3):326-333.
[10] Moreno F,Pelegrin F,Fernandez J E,et al.Soil physical properties,water depletion and crop development undertraditional and conservation tillage in southern Spain[J].Soil&Tillage Research,1997,41(1/2):25-42.
[11] Xu B C,Gichuki P,Shan L,et al.Aboveground biomass production and soil water dynamics of four leguminous forages in semiarid region,northwest China[J].South African Journal of Botany,2006,72(4):507-516.
[12] Betts R A,Boucher O,Collins M,et al.Projected increase in continental runoff due to plant responses to increasing carbon dioxide[J].Nature,2007,448(7157):1037-1041.
[13] Reich P B,Knops J,Tilman D,et al.Plant diversity enhances ecosystem responses to elevated CO2and nitrogen deposition[J].Nature,2001,410(6830):809-812.
[14] Hubbell S P,Foster R B,O′Brien ST,et al.Lightgap disturbances,recruitment limitation,and tree diversity in aneotropical forest[J].Science,1999,283(5401):554-557.
[15] Suding K N,LeJeune K D,Seastedt T R.Competitive impacts and responses of an invasive weed:dependencies onnitrogen and phosphorus availability[J].Oecologia,2004,141(3):526-535.
[16] Xu B C,Li F M,Shan L,et al.Gas exchange,biomass partition,and water relationships of three grass seedlings under water stress[J].Weed Biology and Management,2006,6(2):79-88.
[17] Thomas G V,Biddappa CC,Prabhu S R.Evaluation of N2-fixing cover legumes as green manures for N substitutionin coconut(Cocos nucifera Linn.)palm[J].Tropical Agriculture,2001,78(1):13-18.
[18] Wang W Y,Wang Q J,Wang H C.The effect of land management on plant community composition,species diversity,and productivity of alpine Kobersia steppe meadow[J].Ecological Research,2006,21(2):181-187.
[19] NI J,Wang G H.Northeast China Transect(NECT):Ten-Year Synthesis and Future Challenges[J].Acta Botanica Sinica,2004,46(4):379-390.
[20] Escos J,Pugnaire F I,Puigdefabregas J,et al.Stress resistance strategy in an arid land shrub:interactions between developmental instability and f ractal dimension[J].Journal of Arid Environments,2000,45(4):325-336.
[21] Maggio A,Pascale S de,Ruggiero C,et al.Physiological response of field-grown cabbage to salinity and drought stress[J].European Journal of Agronomy,2005,23(1):57-67.
[22] Bingham I J.Soil-root-canopy interactions[J].Annals of Applied Biology,2001,138(2):243-251.
[23] Shinozaki K,Yamaguchi S,Kazuko S,et al.Gene networks involved in drought stress responseand tolerance[J].Journal of Experimental Botany,2007,58(2):221-227.
[24] Li F M,Liu X L,Li S Q.Effects of early soil water distribution on thedry matter partition between roots and shoots of winter wheat[J].Agricultural Water Management,2001,49(3):163-171.
[25] Shi H,Shao M G.Soil and water loss f rom the Loess Plateau in China[J].Journal of Arid Environments,2000,45(1):9-20.
[26] Aiman E A.Modeling of alkaline scale formation in falling film horizontal-tube multiple-effect distillers[J].Desalination,2007,205(1):124-139.
[27] Shangguan Z P.Soil desiccation occurrence and its impact on forest vegetation in the Loess Plateau of China[J].The International Journal of Sustainable Development and World Ecology,2007,14(3):299-306.
[28] Irvine J,Law B E,Kurpius M R.Coupling of canopy gas exchange with root and rhizosphere respiration in a semi-arid forest[J].Biogeochemistry,2005,73(1):271-282.
[29] Laundre JW.Relationships between water availability,carbon isotope discrimination and plant productivity in two semi-arid grass and shrub species[J].Journal of Arid Environments,1999,41(1):49-60.
[30] James A,Andre F C,James N,et al.Ecological restoration:A new frontier for nature conservation and economics[J].Journal for Nature Conservation,2006,14(3/4):135-139.
[31] Castillo V M,Martinez M M,Albaladejo J.Runoff and soil loss response to vegetation removal in a semiarid environment[J].Soil Science Society of America Journal,1997,61(4):1116-1121.
