不同沙生植被土壤酶活性分异特征研究*
2010-12-21刘济明戴全厚
余 娜,刘济明,张 超,戴全厚
(1.贵州大学林学院,贵阳 550025;2.西北农林科技大学,陕西 杨陵 712100)
沙漠化是当今全球最严重的环境与社会经济问题之一,我国是世界上荒漠化危害严重的国家之一,尤其是我国北方的沙漠化以其面积广大和发展迅速而引人关注。沙漠化过程使土壤的风蚀作用加剧,破坏了土壤的理化性质,蓄水量减少且易蒸发失水,严重降低了土地生产潜力。通过筛选合适的植被类型构建植被群落体系,可以有效地减少沙漠化土壤的风蚀作用,固定流沙,促进土壤有机质和黏粒积累,形成地表结皮,进而改善土壤质量[1-2]。土壤酶是高分子有机物催化分解的一类具有蛋白质性质的生物催化剂,主要来源于土壤微生物的活动、植物根系分泌物和腐解的动植物残体[3],参与土壤中各种有机质的分解、合成与转化,以及无机物质的氧化与还原等过程,是土壤生态系统代谢的一类重要动力,在很大程度上反映土壤物质循环与转化的强度,常被用来反映土壤生态系统变化的预警和敏感指标[4-5],特别是在极其脆弱的荒漠化生态系统中土壤酶的生态作用就显得更加重要[6]。
目前针对沙漠化土壤质量的研究主要集中于土壤理化性质和沙生植物的固沙机理上[7-10],而有关土壤生物学性质虽有个别报道[6],但仍较为薄弱。因此本文选取毛乌素沙地南缘陕西省榆林沙生植物园中9种不同人工固沙植被土壤为研究对象,分析了土壤养分和酶活性的变化,旨在从土壤酶学角度筛选出适合沙地生长的植物类型,从而为制订合理可行的防风固沙林地的建设与管理方案提供参考,为人类研究退化生态系统的恢复和重建提供理论依据和实践指导。
1 材料与方法
1.1 研究区自然概况
该研究区位于毛乌素沙地南缘陕西省榆林沙生植物园(109°41′75″E,38°22′38″N),地处毛乌素沙漠与陕北黄土高原的过渡地带,属于典型的生态脆弱区,平均海拔1 300 m,年均气温10℃,年均降水量400 mm。该区域长期来由于不合理的土地利用,导致严重的土地荒漠化,1958年开始在流动沙丘上进行各项治沙技术和固沙林试验研究,经过多年的工作,流动沙地全部改造为固定沙地,植被盖度达80%以上。
在研究区内选择改造年限、相近坡位坡向一致的9种植被类型为研究对象,它们分别为刺槐(Robinia pseudoacacia,RP),沙蒿(Artemisia desertorum,AD),踏郎(Hedysarum mongolicum,HM),花棒(Hedysarum scoparium,HS),白柠条(Caragana korshinkii,CA),小叶杨(Simon poplar,SP),樟子松(Pinus sylvestris var.mongolica,PS),油松(Pinus tabulaef ormis,PT),臭柏(Sabina vulgaris,SV),并选择一个沙裸地为对照样地(CK),具体样点描述如表1。
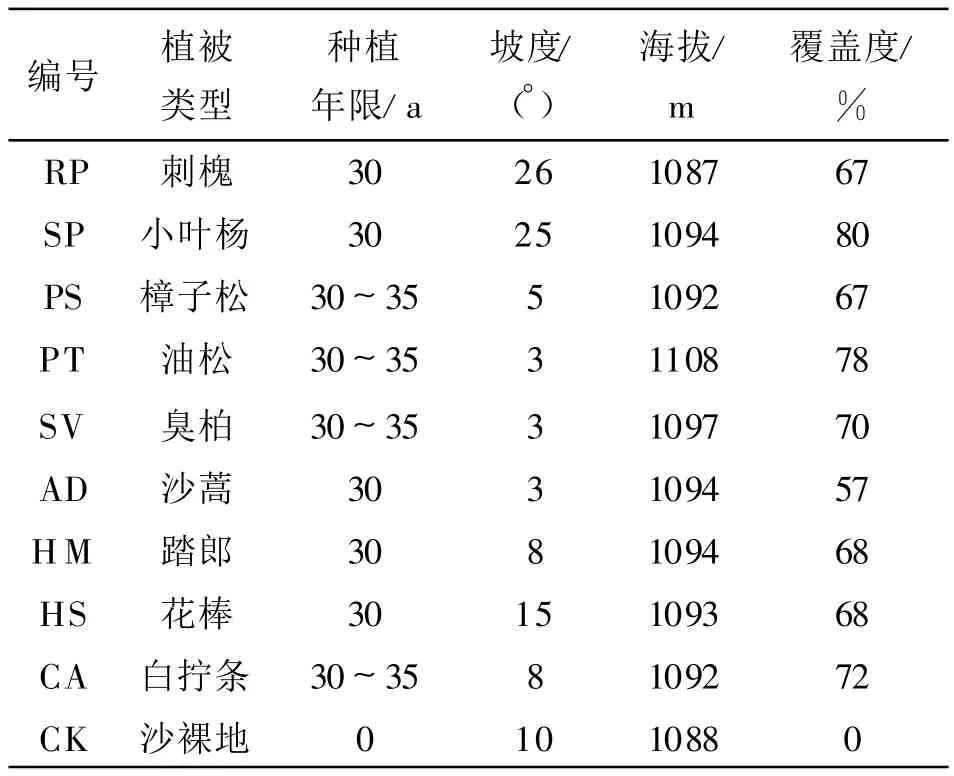
表1 样地基本特征
1.2 样品采集及分析
在试验地各处理小区按S型选取9种沙生植物土壤:用土钻法取0-20 cm混合土样,重复3次。土壤p H值、全氮、全磷、有机质、碱解氮、速效磷及速效钾含量采用常规方法测定[11]。蔗糖酶测定:3,5-二硝基水杨酸比色法,以24 h后1 g土壤中含有的葡萄糖毫克数表示;脲酶测定:靛酚比色法,活性以24 h后1 g土壤中NH 3-N的毫克数表示;磷酸酶测定:磷酸苯二钠比色法,以1 g土壤中24 h后苯酚的毫克数表示;过氧化氢酶测定:滴定法(0.1 mol/L的标准KMnO4液滴定),活性以1 g土壤20 min后消耗0.1 mol/L KMnO4毫升数表示;多酚氧化酶测定:碘量滴定法,酶活性用滴定相当于1 g土壤虑液的0.01 mol/L I2的毫升数[3]。土壤酶评价指数方法参见参考文献[12]。
1.3 数据统计分析
数据分析采用SPSS 15.0统计软件,显著水平为 P<0.05和P<0.01。
2 结果分析与讨论
2.1 不同沙生植物林地土壤化学性质
土壤作为一种独立的自然体,对水、肥、气、热及根系生长空间具有调节功能,同时又受各种环境因素的影响,土地利用方式的变化,可使土壤性质发生显著改变。退化的流动沙地经过植被恢复后,不仅固定了流沙,而且显著改善了土壤化学性质(表1),总体表现为有机质、全氮、碱解氮、全磷、速效磷含量显著增加,分别较沙裸地增加了53.4%~322.1%、49.6%~261.6%、32.5%~167.8%、501.1%~1415%、28.1%~2627%、17.1%~304.8%,p H 值除沙蒿和油松外均呈现降低趋势。在不同化学性质中各种植被对全磷和速效磷的改善作用最强,而不同植被类型对土壤化学性质的作用不一,总体来说臭柏、柠条、花棒、刺槐和樟子松对有机质和氮素的改善作用最强,踏郎、柠条和花棒对速效磷的改善作用最强,小叶杨对速效钾的增加作用最强。
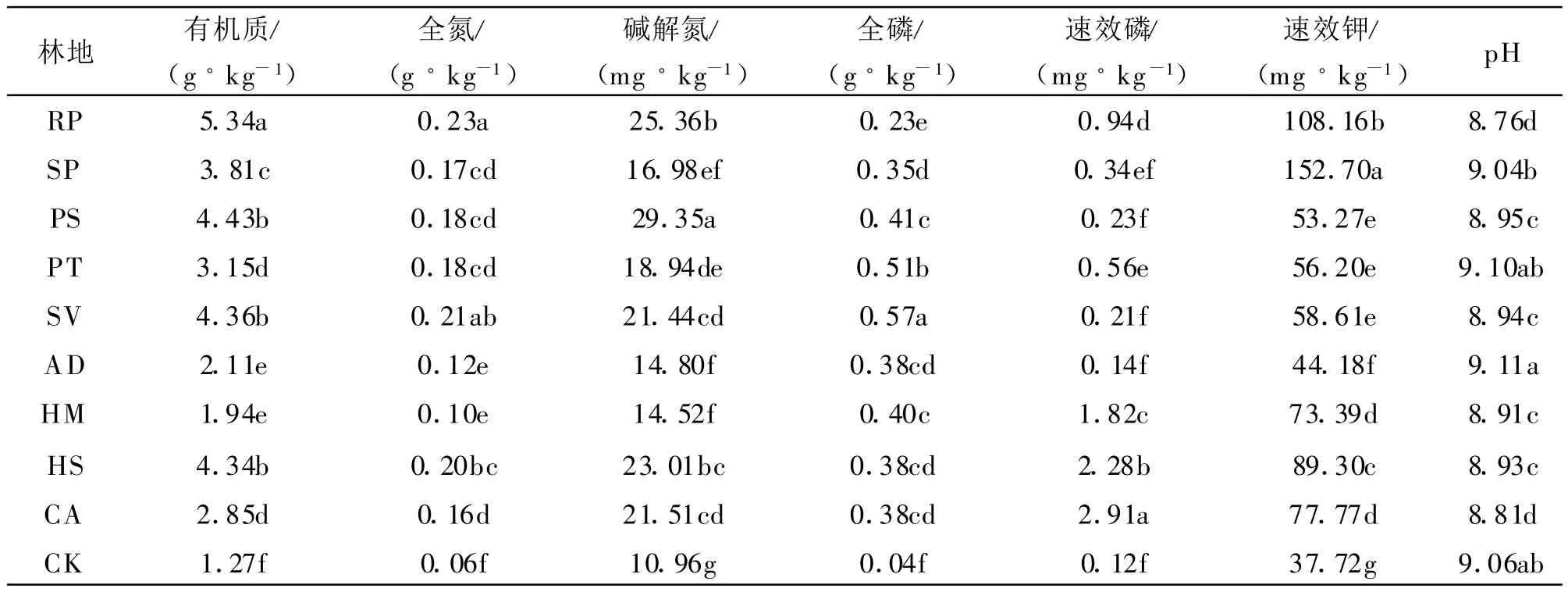
表2 不同沙生植被土壤养分
2.2 不同沙生植物林地土壤酶活性分异特征
2.2.1 蔗糖酶 蔗糖酶直接参与土壤有机质的代谢过程,对增加土壤中易溶性营养物质起着重要的作用,其活性可以作为评价土壤熟化程度和土壤肥力水平的一个指标[3,13-14]。一般情况下,土壤有机质含量越高,蔗糖酶活性越强。沙裸地种植植被后,土壤蔗糖酶活性除沙蒿外均呈现显著增加趋势(图1),分别较沙裸地增加69%~1 107%,其中刺槐和樟子松增幅最大,踏郎增幅最低,不同植被类型增加幅度差异较大。表明沙裸地种植植被后,归还到土壤中的枯枝落叶增多,促进了微生物代谢,蔗糖酶活性明显提高,碳素转化作用增强。
2.2.2 磷酸酶 土壤磷酸酶是生物磷代谢的重要酶类,在土壤磷素循环中起重要作用,可以加速有机磷的脱磷速度。磷酸酶活性与土壤中有机磷的含量呈正相关,活性越高,土壤中有机磷的矿化速度越大,因此其活性可以被用来评价土壤磷素生物转化方向与强度指标和作为生态系统对外界环境因素响应的指示物[15-16]。流沙地土壤磷酸酶活性较低,营造植被后除沙蒿和踏郎外均可显著增加磷酸酶活性(图3),增幅分别达83%~229%,其中樟子松和刺槐增幅最高,小叶杨增幅最低。磷是植物生长的主要营养元素之一,土壤中95%的磷是以迟效性状态存在,当全磷含量低于0.8~1.0 g/kg时,土壤常出现磷供应不足。沙裸地由于没有植被生长,长期处于侵蚀状态,导致土壤磷素贫瘠,随着种植植被,磷酸酶活性明显增强,促进了有机磷向无机磷转化,从而为植物提供了更多的可利用的无机磷酸盐,提高了磷素的有效性。
2.2.3 脲酶 土壤脲酶是存在于土壤中由简单蛋白质构成的生物催化剂,其可将土壤中的有机化合物尿素水解为氨态氮,使植物所需的养分转化为有效态,对提高氮素的利用率和促进土壤氮素循环具有重要意义[3]。沙裸地种植植被后,脲酶活性变化显著(图2),沙裸地上营造其它植被类型后,除小叶杨和踏郎没有显著增高外,其余7种植被类型较沙裸地均有显著增加,增幅达19.9%~152.3%,花棒和白柠条增幅最高,其次为樟子松和沙蒿,最后增幅最小的刺槐、油松和臭柏。安韶山[14]等研究表明植被恢复可以提高土壤脲酶活性,并依赖于有机质的存在,当有机质含量增加时,酶积极参与其转化分解过程,活性提高。本研究也表明植被生长可以增加土壤有机质含量,提高了土壤脲酶转化底物的含量,脲酶活性增加,促进了氮素的循环。
2.2.4 过氧化氢酶 过氧化氢酶能酶促水解过氧化氢分解为水和氧的反应,解除过氧化氢对植物的毒害作用,可以用来表征土壤的生化活性[17],与土壤有机质转化速度密切相关,在土壤物质和能量转化中占有重要地位[18]。如图4所示,种植植被后除踏郎外过氧化氢酶活性均显著高于沙裸地。增幅达54%~271%,其中油松和樟子松增幅最大,柠条增幅最低。以上说明随着沙裸地种植植被有效地增加了土壤中营养物质,促进了土壤生化代谢能力增强,同时产生了更多的过氧化氢等有毒物质,为了缓解生物氧化作用对土壤和生物体的破坏能力,提高过氧化氢酶活性是一个主要的途径。
2.2.5 多酚氧化酶 多酚氧化酶主要起着促进土壤腐殖质组分中芳香族化合物的转化作用,从而形成胡敏酸分子,完成土壤芳香族化合物循环[19-22],常用来反映土壤腐殖质状况。相对于其它几种酶沙裸地种植植被后,多酚氧化酶活性总体变幅不大,仅刺槐、油松和臭柏显著降低,其余6种植被并未有显著变化(图5)。
目前针对植被恢复后土壤多酚氧化酶活性的变化规律还没有一致的结论[23-25],这主要是由于凋落物的分解途径较为复杂,我们的研究表明沙裸地种植不同植被后,虽然凋落物的种类差异较大,但是并未显著影响多酚氧化酶活性,另一方面凋落物可能通过直接降解再缩合形成腐殖质等途径分解,从而造成多酚氧化酶活性的降低。
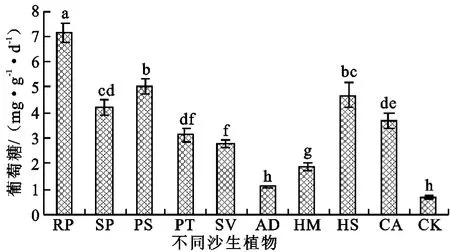
图1 不同沙生植物蔗糖酶活性变化

图2 不同沙生植物脲酶活性变化

图3 不同沙生植物磷酸酶活性变化

图4 不同沙生植物过氧化氢酶活性变化
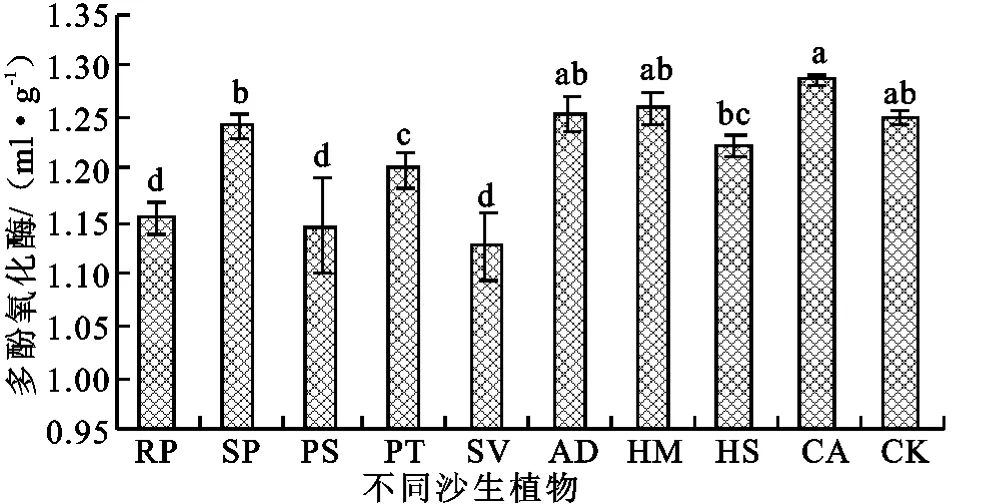
图5 不同沙生植物多酚氧化酶活性变化
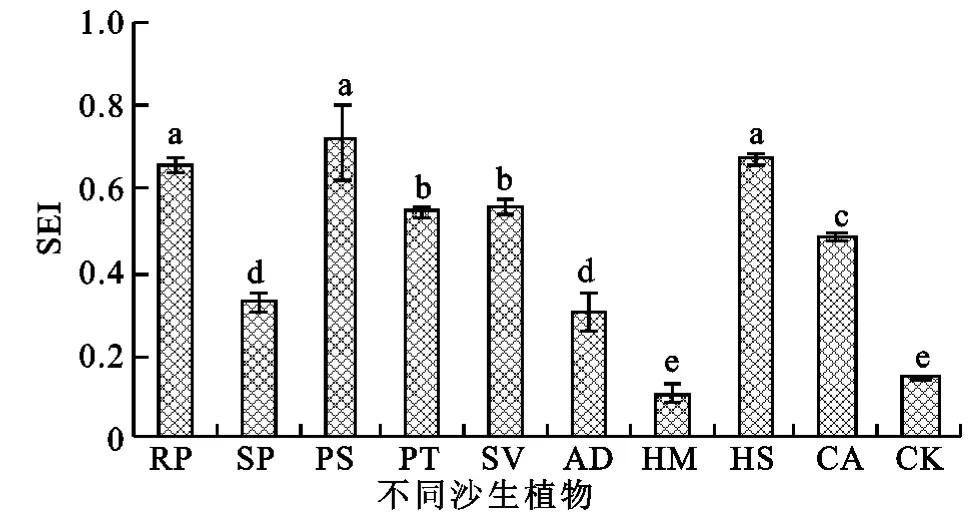
图6 不同沙生植物土壤酶活性变化
2.3 土壤酶活性与养分之间的相关关系
相关性分析表明(表 3),蔗糖酶、磷酸酶、过氧化氢酶、多酚氧化酶活性之间具有显著或极显著的正相关或负相关(P<0.05,P<0.01),且它们还和有机质、全氮和碱解氮具有显著或极显著的相关性,而与全磷、速效磷、速效钾和p H值的相关性相对较弱;脲酶较其它酶活性相关性较低,仅和磷酸酶、碱解氮、速效磷具有较强的相关性。以上结果说明,不同种类的酶活性由于其在土壤中参与的生化反应作用不同,造成和土壤不同养分因子之间的相关性具有一定差异性,但是总体上和土壤养分等具有较强的相关性,可以用来指示治沙过程中土壤质量的演变过程。
2.4 土壤酶指数
从前面的分析可以看出,不同的植被类型对不同的酶种类影响差异都较大,为了克服不同酶活性在反映治理沙裸地的过程中土壤酶属性演变过程的片面性,采用土壤酶指数(SEI)作为酶因子的综合作用表征,从而客观、全面地反映土壤酶活性的变化过程。
研究表明(图6),SEI在沙裸地种植植被后变化显著,增幅从大到小依次为刺槐林和樟子松、花棒、油松、臭柏、白柠条、小叶杨、沙蒿,踏郎和沙裸地没有显著差异。沙裸地由于没有植被生长,因此很难有枯落物等物质归还到土壤中,造成土壤养分低下,加之水分亏缺,严重地制约了微生物的生长与繁殖,因此土壤酶活性相对较低,种植植被后,归还到土壤中的枯落物显著增加,促进了微生物的生长与繁殖,酶活性增加。不同植被类型由于其枯落物种类、数量和根系分泌物等多种因素的差异,导致微生物种群数量发生异质性,土壤酶活性产生分化。
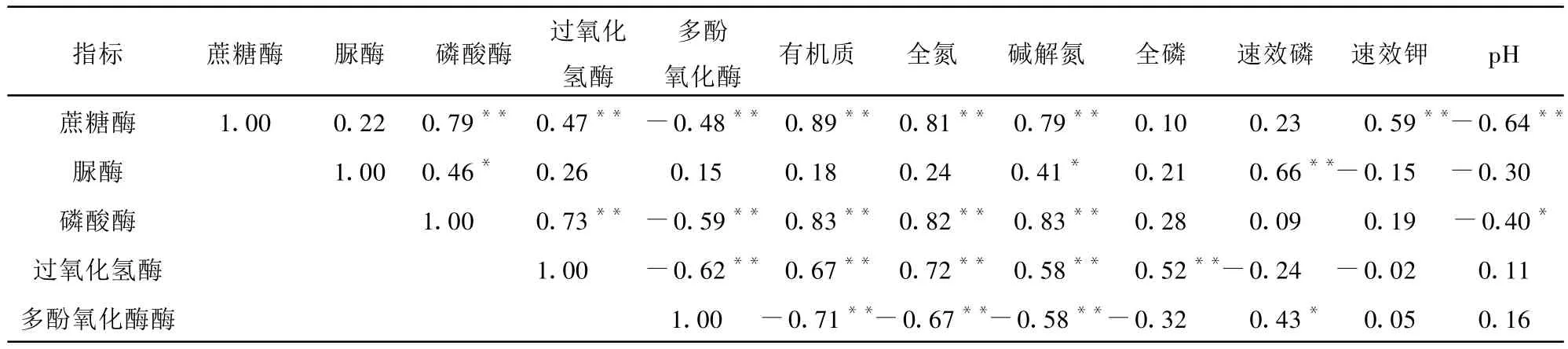
表3 土壤酶活性与养分之间的相关分析
3 结论
(1)沙裸地种植植被后,土壤养分得到改善,总体表现为有机质、全氮、碱解氮、全磷、速效磷含量显著增加,p H 值降低,臭柏、柠条、花棒、刺槐和樟子松对有机质和氮素的改善作用最强,踏郎、柠条和花棒对速效磷的改善作用最强,小叶杨对速效钾的增加作用最强。
(2)土壤酶随沙裸地种植植被变化明显,蔗糖酶、磷酸酶、脲酶和过氧化氢酶活性总体呈升高趋势,多酚氧化酶有所降低;不同植被类型对不同酶活性的改善作用表现出较强的异质性;相关性分析表明土壤蔗糖酶、磷酸酶、过氧化氢酶、多酚氧化酶活性之间具有较强的相关性,并和有机质、全氮和碱解氮相关性较高;SEI在沙裸地种植植被后显著增加,增幅从大到小依次为刺槐林和樟子松、花棒、油松、臭柏、白柠条、小叶杨、沙蒿,踏郎和沙裸地没有显著差异。
[1] 蒋德明,宗文君,李雪华,等.科尔沁西部地区荒漠化土地植被恢复技术研究[J].2006,25(3):243-248.
[2] 赫晓慧,常庆瑞,温仲明,等.农牧交错带不同人工植被下荒漠化土壤肥力的变化[J].中国沙漠,2006,26(6):915-919.
[3] 关松荫.土壤酶及其研究法[M].北京:农业出版社,1986.
[4] 周礼恺.土壤酶学[M].北京:科学出版社,1987:118-159.
[5] 南京土壤所.土壤微生物研究法[M].北京:科学出版社,1985.
[6] 徐恒,廖超英,李晓明,等.榆林沙区人工固沙林土壤微生物生态分布特征及酶活性研究[J].西北农林科技大学学报:自然科学版,2008,36(12):135-141.
[7] 曹成有,蒋德明,全贵静,等.科尔沁沙地小叶锦鸡儿人工固沙区土壤理化性质的变化[J].水土保持学报,2004,18(6):108-111.
[8] 吴祥云,刘广,韩辉.不同类型樟子松人工固沙林土壤质量的研究[J].防护林科技,2001(4):15-17.
[9] 吴祥云,姜风岐.樟子松人工固沙林衰退的规律和原因[J].应用生态学报,2004,15(12):2225-2228.
[10] 贺山峰,蒋得明,阿拉木萨,等.科尔沁沙地小叶锦鸡儿灌木林固沙效应的研究[J].水土保持学报,2007,21(1):84-87.
[11] 中国生态系统研究网络观测与分析标准方法:土壤理化分析与剖面描述[M].北京:中国标准出版社,1996.
[12] 王兵,刘国彬,薛萐,等.黄土丘陵区撂荒对土壤酶活性的影响[J].草地学报,2009,17(3):282-287.
[13] 叶存旺,翟巧绒,郭梓娟,等.沙棘-侧柏混交林土壤养分、微生物与酶活性的研究[J].西北林学院学报,2007,22(5):1-6.
[14] 张旭辉,潘根兴.连续多年栽培叶用银杏下土壤养分及酶活性变化[J].土壤通报,2004,35(1):21-25.
[15] 于群英.土壤磷酸酶活性及其影响因素研究[J].安徽技术师范学院学报,2001,15(4):5-8.
[16] 沈菊培,陈振华,陈利军,等.草甸棕壤水稻田磷酸酶活性及对施肥措施的响应[J].应用生态学报,2005,16(3):583-585.
[17] 薛立,邝立刚,陈红跃,等.不同林分土壤养分、微生物与酶活性的研究[J].土壤学报,2003,40(2):280-285.
[18] 杨万勤,王开运.土壤酶研究动态与展望[J].应用与环境生物学报,2002,8(5):564-570.
[19] Durin N,Esposito E.Potential applications of oxidative enzymes and phenol-oxidase-like compounds in wastewater and soil treatment:a review[J].Applied Catalysis B:Environmental,2000,28:83-99.
[20] Insam.Developments in soil microbiology since the mid 1960s[J].Geoderma,2001,100:389-402.
[21] Diamantidis G,Eosse,Potier P,Rene.Purification and characterization of thefirst bacteria1laecase in the rhizospheric bacterium Azospirillum lipoferum[J].Soil Biology&Biochemistry,2000,32:919-927.
[22] Toseano G,Maria L,Greco G.Oxidative polymerisation of phenols by a phenol-oxidase from green olives[J].Enzyme and Microbial Technology,2003,33:47-54.
[23] 郑华,欧阳志云,易自力.红壤侵蚀区恢复森林群落物种多样性对土壤生物学特性的影响[J].2004,18(4):137-141.
[24] 李传荣,许景伟,宋海燕,等.黄河三角洲滩地不同造林模式的土壤酶活性[J].植物生态学报,2006,30(5):802-805.
[25] ZHANG Yong-mei,ZHOU Guo-yi,WU Ning,et al.Soil Enzyme Activity Changes in Different-Aged Spruce Forests of the Eastern Qinghai-Tibetan Plateau[J].Pedosphere,2004,14(3):305-312.
