P2X7受体介导异丙酚对缺氧海马突触前膜谷氨酸Ca2+依赖性释放的抑制作用
2010-11-29刘红亮戴体俊
刘红亮,戴体俊
谷氨酸兴奋毒性是脑缺血/氧损伤的重要机制,细胞外液的谷氨酸含量主要由突触前膜释放和胶质细胞重摄取决定。以往研究发现[1],异丙酚(propofol,P)可促进重摄取功能降低谷氨酸含量而产生脑保护作用。生理状态下,突触前膜释放谷氨酸通过两种机制:Ca2+依赖性释放(囊泡释放)和Ca2+非依赖性释放(谷氨酸转运体反向转运)[2]。而缺血/氧时,异丙酚对谷氨酸的两种释放机制产生何种作用目前尚不清楚。中枢神经系统内,ATP是一种重要神经递质,其离子型受体P2X受体有7种亚型,其中P2X7受体在突触前膜表达丰富而且在调节谷氨酸释放中发挥关键作用[3]。缺血/氧时,谷氨酸释放是否亦受P2X7受体调控以及异丙酚是否通过作用于突触前膜P2X7受体而影响谷氨酸释放均值得研究。因此,本实验采用海马突触前膜缺氧模型,观察P2X7受体在谷氨酸Ca2+依赖性释放和Ca2+非依赖性释放中的作用,异丙酚对谷氨酸释放的影响及P2X7受体在其中的作用。
1 材料
1.1 实验药品与仪器 异丙酚为AstraZeneca公司产品(批号:CFF755);谷氨酸标准品、dihydrokainic acid(DHK,谷氨酸转运体抑制剂)、P2X7受体特异性拮抗剂亮蓝G(BBG)及特异性激动剂BZATP为Sigma公司产品;其他试剂均为国产市售分析纯。GL20C高速冷冻离心机(中科院武汉科学仪器厂);HPLC荧光检测器(美国 Varian公司);SinochromODS 4.6 mm ×250 mm 5 μm C18色谱分析柱(大连依利特科学仪器有限公司)。
1.2 实验动物 Sprague-Dawley♂大鼠,150~200 g,由徐州医学院实验动物中心提供。
2 方法
2.1 人工脑脊液(aCSF)的制备 aCSF成分为(mmol·L-1):NaCl 126.5,NaHCO327.5,KCl 2.4,KH2PO40.5,CaCl21.1,MgCl20.83,Na2SO40.5,Glucose 11.8,DHK 0.05,将 pH 值调至 7.3,用 O2/CO2(95%/5%)充分饱和。用于缺氧缺糖实验的无糖aCSF,将glucose置换为同等浓度的sucrose,并持续充以95%N2/5%CO2。用于谷氨酸Ca2+非依赖性释放试验时,将Ca2+、DHK从aCSF中去除。
2.2 突触体的制备 大鼠击昏断头、取脑置于冰面上,分离海马,按1/10(W/V)加入到0.32 mol·L-1的冷蔗糖溶液中进行匀浆,以1 000×g离心5 min,取上清液再以12 000×g离心20 min,所得沉淀为突触体,以上步骤在0~4℃操作完成[4]。
2.3 突触体缺氧缺糖实验 将制备的突触体先应用aCSF于37℃下孵育20 min,然后移入无糖aCSF中,于37℃下进行缺氧实验。观察P2X7受体对谷氨酸释放的影响时,将突触体分为4组,假性处理组(Sham,不缺氧缺糖)、对照组(Control,缺氧缺糖)、BBG 组(终浓度 1 μmol·L-1)、BZATP 组(终浓度100 μmol·L-1)。每组分别于缺氧前(基础值)、缺氧20 min及40 min时收集孵育液测定谷氨酸浓度,谷氨酸Ca2+非依赖性释放试验中,将DHK从aCSF中去除。根据结果确定下一步实验。
另将制备的突触体预先于37℃下应用aCSF孵育20 min,然后移入无糖aCSF中进行缺氧40 min。观察异丙酚对谷氨酸释放的影响时,将突触体随机分为 6 组,P 0 μmol·L-1组、P 1 μmol·L-1组、P 3 μmol·L-1组、P 10 μmol·L-1组、P 30 μmol·L-1组和P 100 μmol·L-1组(孵育液中分别含异丙酚浓度为 0、1、3、10、30、100 μmol·L-1)。各组在实验结束时收集孵育液进行谷氨酸浓度测定并计算异丙酚的半效抑制浓度(IC50)。
观察P2X7受体在异丙酚抑制效应中的作用时,将突触体分为4组,对照组(Control,缺氧缺糖)、Propofol组(孵育液中含2倍IC50水平的异丙酚)、Propofol+BBG组(孵育液中含2倍IC50水平的异丙酚和 BBG 1 μmol·L-1)及 Propofol+BZATP 组(孵育液中含2倍IC50水平的异丙酚和BZATP 100 μmol·L-1)。各组于缺氧40 min后收集孵育液进行谷氨酸浓度测定。以上实验组每组含6份突触体,每份突触体中蛋白含量为0.26 g·L-1,蛋白含量应用考马斯亮蓝法测定。
2.4 谷氨酸浓度测定 应用反相高效液相色谱法(RP-HPLC)测定各实验组突触体孵育液中的谷氨酸浓度[5]。具体步骤如下:配制衍生剂:邻苯二甲醛 27 mg、2-巯基乙醇 40 μl溶于 0.5 ml甲醇中,加入硼酸钠溶液并调节pH值为7.0。取样本50 μl,衍生剂25 μl,振荡反应1 min 后进样20 μl。A 流动相为0.5 mol·L-1乙酸钠(内含体积分数为0.05的四氢呋喃),B流动相为甲醇。洗脱程序为(T,%):(0,32);(7,62);(9,32),其中 T 为洗脱时间(min),%为B相在流动相中的浓度。流速0.9 ml·min-1,柱温38℃。荧光检测器 λex为360 nm,λem为450 nm。应用Prostar工作站进行梯度控制和色谱数据处理。应用外标法计算谷氨酸浓度。谷氨酸突触体释放量以每克蛋白(g protein)含有谷氨酸的毫克数来表示,单位:mg·g-1Pro。
3 结果
3.1 P2X7受体拮抗剂和激动剂对缺氧海马突触前膜谷氨酸释放的影响 各组于缺氧前的基础释放水平差异无显著性。对照组中,缺氧20 min和40 min时,谷氨酸Ca2+依赖性释放增高(P<0.01)。而BBG组中,缺氧不同时点的谷氨酸Ca2+依赖性释放水平与Sham组比较差异无显著性(P>0.05),与对照组比较则降低(P<0.01)。BZATP组中,缺氧不同时点谷氨酸Ca2+依赖性释放水平高于对照组(P<0.01)。而缺氧不同时点,各组中谷氨酸Ca2+非依赖性释放无变化,组间差异亦无显著性(P>0.05)。见 Tab 1。
3.2 异丙酚对谷氨酸Ca2+依赖性释放的影响 异丙酚1~100 μmol·L-1可浓度依赖性的抑制缺氧海马突触前膜谷氨酸Ca2+依赖性释放。根据Hill公式,异丙酚对谷氨酸Ca2+依赖性释放的IC50为(27.4 ±5.2)μmol·L-1(Tab 2)。
Tab 1Effect of P2X7 receptor antagonist and agonist on glutamate release(mg·g-1Pro,±s,n=6)

Tab 1Effect of P2X7 receptor antagonist and agonist on glutamate release(mg·g-1Pro,±s,n=6)
**P<0.01 vs sham;##P<0.01 vs control
Group Ca2+-dependent release Baseline 20 min 40 min Ca2+-independent release Baseline 20 min 40 min Sham 1.20±0.20 1.15±0.28 1.20±0.23 0.51±0.12 0.48±0.13 0.55±0.14 Control 1.15±0.20 2.82±0.27** 4.07±0.41** 0.46±0.15 0.50±0.13 0.50±0.14 BBG 1.15±0.25 1.23±0.21## 1.12±0.20## 0.50±0.15 0.50±0.13 0.46±0.15 BZATP 1.16±0.27 5.23±0.42**## 5.10±0.48**##0.47±0.15 0.48±0.15 0.50±0.11
3.3 P2X7受体在异丙酚抑制谷氨酸释放中的作用 55 μmol·L-1(2倍IC50)异丙酚可抑制缺氧海马突触前膜谷氨酸Ca2+依赖性释放(P<0.01),在异丙酚和BBG或异丙酚和BZATP的共同作用下,谷氨酸释放水平低于对照组(P<0.01),但与异丙酚单独作用比较,差异无显著性(P>0.05),见Tab 3。
Tab 2 Effect of propofol on glutamate release(mg·g-1Pro±s,n=6)

Tab 2 Effect of propofol on glutamate release(mg·g-1Pro±s,n=6)
Group Glutamate release P 0 μmol·L-14.36±.51 P 1 μmol·L-1 4.22 ±0.43 P 3 μmol·L-1 3.86 ±0.40 P 10 μmol·L-1 2.46 ±0.23 P 30 μmol·L-1 1.30 ±0.20 P 100 μmol·L-11.20±0.19
Tab 3 Role of P2X7 receptor in the inhibitory effect of propofol±s,n=6)

Tab 3 Role of P2X7 receptor in the inhibitory effect of propofol±s,n=6)
**P<0.01 vs control
Group Glutamate release/mg·g-1Pro Control 4.21±0.60 Propofol 1.29±0.19**Propofol+BBG 1.24±0.20**Propofol+BZATP 1.28±0.23**
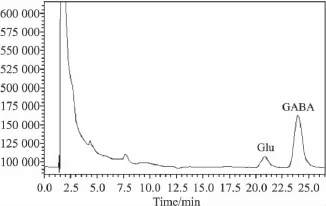
Fig 1 Standard chromatograph of Glu(10 mg·L-1)and GABA(10 mg·L-1)in RP-HPLC
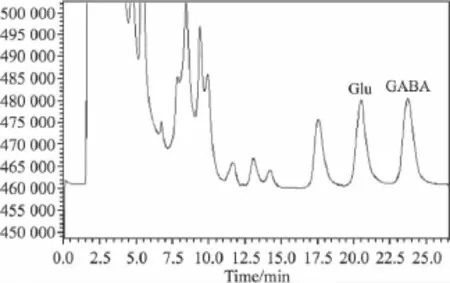
Fig 2 Chromatograph of Glu and GABA in sample in RP-HPLC
4 讨论
突触前膜释放谷氨酸有两种机制:Ca2+依赖性释放即量子式囊泡释放和Ca2+非依赖性释放即由突触前膜的谷氨酸转运体反向转运而释放[2]。本实验结果表明,在缺氧条件下,谷氨酸的释放为Ca2+依赖性,而且这种释放由突触前膜P2X7受体介导。P2X7受体在中枢神经系统内分布广泛,尤其在突触前膜表达丰富。P2X7受体被其内源性配体ATP激活后,离子通道开放而使得Ca2+、Na2+内流,引起突触前膜去极化[7]。ATP作为中枢神经系统内一种重要神经递质,生理状态下通过突触前膜P2X7受体调节其他神经递质的释放和功能[8]。BZATP是人工合成的P2X7受体特异性激动剂,在研究P2X7受体功能时是一种重要的工具药。本实验中,P2X7受体特异性拮抗剂BBG可完全拮抗缺氧时谷氨酸的释放,而特异性激动剂BZATP可进一步使谷氨酸释放增加,表明缺氧条件下,激活P2X7受体可促使谷氨酸释放。
静脉麻醉药异丙酚对脑缺血损伤具有保护作用[9],但亦有研究发现异丙酚对大鼠皮层和海马脑片谷氨酸引起的损伤无影响[10],本实验发现异丙酚可浓度依赖性的抑制缺氧环境下突触前膜谷氨酸Ca2+依赖性释放。在进行谷氨酸Ca2+依赖性释放时,在aCSF中加入谷氨酸转运体抑制剂DHK,以排除Ca2+非依赖性释放和重摄取的影响,使得结果更具有可靠性。本实验中异丙酚抑制谷氨酸释放的IC50为 27.4 μmol·L-1,而异丙酚临床麻醉浓度的EC50为 2 μmol·L-1(游离浓度)[11],表明抑制缺氧环境下突触前膜谷氨酸Ca2+依赖性释放的浓度远高于临床麻醉所需浓度。而临床相关浓度的异丙酚是否可通过其他机制产生脑保护作用尚需进一步研究。
在2倍IC50异丙酚作用下,突触前膜P2X7受体可被完全抑制,使得加入BBG或BZATP后P2X7受体效应无变化。由于突触前膜体积小,难以应用电生理学测定异丙酚对P2X7电流的影响。但以往研究发现[12],异丙酚可抑制巨噬细胞膜P2X7受体电流,且IC50与本实验相近。鉴于突触前膜P2X7受体在谷氨酸Ca2+依赖性释放中的介导作用,可以认为异丙酚抑制缺氧环境下谷氨酸的Ca2+依赖性释放是通过抑制突触前膜P2X7受体来实现的。
总之,突触前膜P2X7受体介导了缺氧环境下谷氨酸的Ca2+依赖性释放,异丙酚可浓度依赖性的抑制谷氨酸Ca2+依赖性释放且这种效应由P2X7受体介导。
[1]Valley J,Guillet B A,Masmejean F M,et al.Neuroprotective effects of propofol in a model of ischemic cortical cell cultures[J].Anesthesiology,2003,99(2):368-75.
[2]杨雄里,译.神经生物学·从神经元到脑[M].北京:科学出版社,2003:81-2.
[2]Yang X L.Neurobiology·from neuron to brain[M].Beijing:Science Press,2003:81-2.
[3]Marcoli M,Cervetto C,Paluzzi P,et al.P2X7 pre-synaptic receptors in adult rat cerebrocortical nerve terminals:a role in ATP-induced glutamate release[J].J Neurochem,2008,105(6):2330-42.
[4]吴春福,汪范生.离体脑组织神经递质释放研究方法[M]//徐叔云,卞如濂,陈修主编.药理实验方法学,第3版.北京:人民卫生出版社,2001:773.
[4]Wu C F,Wang F S.Methods of neurotransmitter release from brain tissue in vitro[M].//Xu S Y,Bian R L,Chen X,chief editor.Methodology of pharmacological experiments.3rd ed,Beijing:People′s Health Press,2001:773.
[5]刘红亮,戴体俊,姚尚龙.硫喷妥钠对大鼠前额叶皮层突触体谷氨酸和GABA释放的影响[J].中国药理学通报,2004,20(11):1235-8.
[5]Liu H L,Dai T J,Yao S L.Effect of thiopental sodium on the release of glutamate and GABA from rat prefrontal cortical synaptosomes[J].Chin Pharmacol Bull,2004,20(11):1253-8.
[6]Joo D T,Xiong Z,MacDonald J F,et al.Blockade of glutamate receptors and barbiturate anesthesia:increased sensitivity to pentobarbital-induced anesthesia despite reduced inhibition of AMPA receptors in GluR2 null mutant mice[J].Anesthesiology,1999,91(5):1329-41.
[7]North R A.Molecular physiology of P2X receptors[J].Physiol Rev,2002,82(4):1013-67.
[8]Khakh B S,Gittermann D,Cockayne D A,et al.ATP modulation of excitatory synapse onto interneurons[J].J Neurosci,2003,23(19):7426-37.
[9]Gelb A W,Bayona N A,Wilson J X,et al.Propofol anesthesia compared to awake reduces infart size in rats[J].Anesthesiology,2002,96(5):1183-90.
[10]于布为,薛庆生,夏 梦,等.丙泊酚对大鼠脑片不同性质损伤的影响[J].中华医学杂志,2003,83(13):1176-9.
[10]Yu B W,Xue Q S,Xia M,et al.The influences of propofol on different kinds of brain injuries in rat brain slices[J].Chin Med J,2003,83(13):1176-9.
[11]Smith C,McEwan A,Jhaveri R,et al.The interaction of fentanyl on the Cp50 of propofol for loss of consciousness and skin incision[J].Anesthesiology,1994,81(4):820-8.
[12]刘红亮,戴体俊.利多卡因在异丙酚抑制巨噬细胞膜P2X7受体电流中的双向调节作用[J].中国药理学通报,2009,25(7):911-4.
[12]Liu H L,Dai T J.Impact of lidocaine on the inhibitory effect of propofol on P2X7-gated currents[J].Chin Pharmacol Bull,2009,25(7):911-4.
