染料木黄酮生物活性及抗病机理的研究进展
2010-11-02曲妍妍张永忠
曲妍妍,张永忠,2,*
(1.东北农业大学理学院应用化学系,黑龙江哈尔滨150030; 2.教育部大豆生物学重点实验室,黑龙江哈尔滨150030)
染料木黄酮生物活性及抗病机理的研究进展
曲妍妍1,张永忠1,2,*
(1.东北农业大学理学院应用化学系,黑龙江哈尔滨150030; 2.教育部大豆生物学重点实验室,黑龙江哈尔滨150030)
染料木黄酮是苷元形式的大豆异黄酮,具有多种生物学活性。动物实验、临床研究及流行病学调查表明,染料木黄酮对白血病、病毒感染、肿瘤等具有防治作用,并具有防辐射及预防骨质疏松的功能。它的抑制细胞免疫和体液免疫以及调节细胞凋亡等作用也日益受到重视。本文简述了染料木黄酮的生物活性及抗白血病、抗病毒感染、抗肿瘤、抗辐射及预防骨质疏松的机理。
染料木黄酮,白血病,病毒感染,抗肿瘤,抗辐射,骨质疏松
大豆异黄酮是大豆生长过程中形成的一类次生代谢产物。大豆中天然存在的异黄酮共有12种,可以分为3类,即黄豆苷类(Daidzin groups)、染料木苷类(Genistingroups)、黄豆黄素苷类 (Glycitin groups),每类以游离型(苷元)、葡萄糖苷型、乙酰基葡萄糖苷型、丙二酰基葡萄糖苷型等4种形式存在[1]。目前国内在大豆异黄酮研究中所用名词术语比较混乱,不够规范和严肃。表现为一个英文名词(如genistein)被译成多个中文名词,而有时又将两个英文名词译成同一个中文名词。本论文关于大豆异黄酮的名词术语依据于文献[2],如将genistein译成染料木黄酮[2]。这是因为genistein是大豆异黄酮苷元,从化学结构上看是黄酮类化合物,化学名称为5,7,4'-三羟基异黄酮。因最初发现存在于染料木属(Genista)植物中,所以最初译成染料木因(科学出版社1961年出版的《英汉化学化工词汇》)或染料木素。金雀花属(Cytisus scoparius)植物中也发现了genistein,所以有人将genistein译成了金雀异黄素。科学出版社2000年出版的《英汉化学化工词汇》(第4版)以全国科学技术名词审定委员会已公布的化学化工标准名词为依据,将genistein校译成染料木黄酮。各种大豆中异黄酮总含量约为1200~4216μg/g,以异黄酮葡萄糖苷和丙二酰基异黄酮葡萄糖苷为主要形式,共占异黄酮总量的95%~98%,而游离型异黄酮(苷元)的含量很低。染料木黄酮是一种主要存在于豆科植物中生物合成的抗微生物毒素的前体,是大豆异黄酮复合物中最简单的一种,但染料木黄酮是12种大豆异黄酮中的一种主要活性因子,是大豆异黄酮产品中最有效的功能成分,具有多种生理功能。它是一种有效的抗氧化剂,一种有效的蛋白酪氨酸激活酶抑制剂,一种植物雌激素。国外在研究大豆异黄酮的生理活性时,应用的原料不是大豆异黄酮粉(染料木黄酮含量极少的一种混合物),而是采用具有明确化学结构的染料木黄酮。染料木黄酮在动物细胞中具有广泛药理学功效,它的多种生物活性已在过去10年中被3600余篇文献所报道,大多数的研究都集中在染料木黄酮作为一种酪氨酸蛋白激酶(TPK)抑制剂的药理学活性上,包括化学防癌、预防白血病的活性,以及抗辐射和抗病毒感染的活性。通过流行病学和动物模型研究发现,膳食中摄入的染料木黄酮具有许多对人体健康有益作用的生物活性成分,可以预防甚至治疗许多疾病,包括乳腺癌、前列腺癌、白血病和病毒感染等[3]。本文对有关染料木黄酮的生物活性进行综述,并简述染料木黄酮治疗各种疾病的作用机制。
1 染料木黄酮的化学结构、抗氧化活性及其代谢产物
任何物质的宏观性质主要是由其内在的结构决定的。同样,染料木黄酮和黄豆苷元之所以被称为植物雌激素,是因为其结构与哺乳动物内源雌激素——雌二醇结构相似,具有活性基团——二酚羟基,其中7,4'两个羟基间的距离几乎等于17β-雌二醇上3,17位羟基间的距离。正是由于这种特殊的结构,能够与雌激素靶细胞结合而呈现出一定的雌激素功能。染料木黄酮和17β-雌二醇化学结构如下:
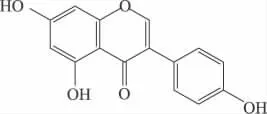
图1 染料木黄酮(genistein)
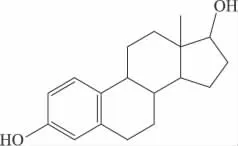
图2 17 β-雌二醇(Estradiol)
Lamartiniere C A报道,与17 β-雌二醇相比,异黄酮类化合物的双环结构及每个芳环上至少有一个羟基,这对于发挥活性是必不可少的,尤其是4'-OH直接与受体结合,结构特异性高,决定着与雌激素受体亲和力的大小[4]。Song TT等人以大鼠子宫增重法(uterine weight method)对雌性B6D2F老鼠进行实验,发现已烯雌酚、黄豆黄素、染料木黄酮和黄豆苷元的相对雌激素作用强度为105、3、1和0.26。17β-雌二醇,乙烯雌酚,染料木黄酮、黄豆苷元和大豆黄素对子宫雌激素受体的相对结合强度为100%、95%、0.49%、0.027%和0.028%[5]。由此可以发现在植物雌激素中,羟基的位置与个数决定了其活性的高低。Adams NR也指出植物雌激素必须具有能够与此激素受体相结合的结构,而双羟基结构正是与雌激素受体结合的主要部位,对于其发挥雌激素作用是非常重要的[6]。
不同结构的异黄酮抗氧化能力也是有差异的[7-8]。Corinna E.R等人对染料木黄酮、黄豆苷元、鸡豆黄素和芒柄花黄素及其代谢产物体外抗氧化研究发现,染料木黄酮的抗氧化能力最强,而芒柄花黄素最低。并总结分析得出,染料木黄酮的活性之所以比黄豆黄素和樱黄素(prunetin)高是由于它的A环上有5,7-两个羟基,黄豆苷元少一个5位羟基,樱黄素的7位是甲氧基。染料木黄酮和大豆黄素(daidzein)代谢产物在A环或B环有双羟基结构,且抗氧化能力均比原物质高,因此可以确定这种特殊结构可以增强物质的抗氧化能力。通过比较大豆黄素和6-OH大豆黄素,染料木黄酮和鸡豆黄素,芒柄花黄素和黄豆苷元,樱黄素和染料木黄酮的抗氧化能力,发现前者的能力均高于后者,因此可以确定是由于它们分别由一个相应碳原子上的羟基被甲氧基取代所造成的。由于鸡豆黄素比芒柄花黄素多一个5-OH,所以鸡豆黄素比芒柄花黄素的抗氧化能力略高[9]。
大豆异黄酮多以其各种葡萄糖苷的形式存在,这些葡萄糖苷只有在体内经水解作用形成苷元的形式,才具有较强的生理作用。Toru Izumi等[10]研究表明:在人体中,大豆异黄酮苷元的吸收速度与吸收量都比其糖苷形式大得多。大豆异黄酮的苷元包括染料木黄酮(genistein)、黄豆苷元(daidzein)和黄豆黄素(glycitein,主要存在于大豆胚芽中),其中染料木黄酮的活性最高。在未加工的植物体中,异黄酮主要以糖苷形式存在,被人和动物摄入体内后,在肠道细菌、微生物的β-2-葡萄糖苷酶或β-2-半乳糖苷酶的作用下,转化为具有生物活性的苷元,即转变成游离形式。这些分解产物一部分进入肠壁组织,另一部分尚未分解的异黄酮苷随粪便一起排出。被吸收的异黄酮通过血管被运送到肝脏中,并且在肝脏中迅速合成,合成的主要是葡萄糖醛酸。在肠道不含上述细菌的动物体内,其血液和胆汁中均不存在大豆异黄酮。因此,如果使用抗生素,则会使身体中的异黄酮含量缺乏。例如:染料木黄酮的代谢半衰期是8.4 h,因此,每天食用2~3次,能使异黄酮的血液浓度维持稳定。
膳食大豆异黄酮的主要来源是大豆产品。1g大豆粉含近800μg大豆黄素和超过500μg的染料木黄酮(主要是糖苷类);1g大豆蛋白含近150μg的大豆黄素和250μg的染料木黄酮。精制的大豆产品,如枸橼酸铋和酱油所含染料木黄酮的水平比豆腐的含量低,豆腐是亚洲膳食中大豆异黄酮的主要来源。大豆异黄酮的摄入水平可以通过尿液排泄物来测定,那些常吃富含大豆食品的成年人,他们的尿酚含量比那些很少吃大豆产品的多近100倍[11]。
2 染料木黄酮的抗白血病活性[12]
Raynal等研究发现,染料木黄酮能够抑制人类和鼠类白血病细胞株生长。他们采用的是克隆原分析法,这是一种最主要的评估体外抗肿瘤制剂活性的方法。染料木黄酮降低白血病细胞株的集落生成能力在评估其抗肿瘤活性方面是非常重要的。
该研究还发现,染料木黄酮诱导了骨髓和淋巴白血病细胞株对时间和剂量依从性效应。在1~20μmol·L-1时不生成白血病细胞集落,这个浓度在富含大豆的饮食所提供的染料木黄酮血浆浓度范围内。有趣的是,低浓度(2μmol·L-1)染料木黄酮处理24h后也能引起集落生成率降低10%。已有报道说染料木黄酮对肿瘤细胞来说是细胞毒性制剂而对正常细胞则不是。另外,染料木黄酮对病人没有遗传毒性。
他们还发现染料木黄酮对肿瘤抑制基因的复活作用,该基因被白血病细胞株中的启动子超甲基化作用所抑制。染料木黄酮(1~20μmol·L-1)诱导了沉寂的肿瘤抑制基因(人细胞中p57KIP2和鼠细胞中p15CDKN2B)的再表达。在经染料木黄酮处理后,这些基因的可变cDNA的扩增可能是由异黄酮在不同靶物质上的多效性作用引起的。染料木黄酮作用于食管鳞状细胞癌细胞,使几个肿瘤抑制基因重新表达,这些基因之前均被变体DNA甲基化所抑制。染料木黄酮对肿瘤抑制基因的复活和脱甲基作用说明了这些制剂的部分抗白血病活性与它后生的作用有关。它对靶物质的作用,就像抑制酪氨酸激酶的活性,可能也对其抗白血病功效有贡献。例如,酪氨酸激酶抑制剂有抗肿瘤活性并被临床验证。
3 染料木黄酮抑制沙粒病毒感染[13]
沙粒病毒属有包膜病毒家族,能够引起人类出血热综合症。染料木黄酮是一种酪氨酸激酶抑制剂,能够抑制一些病毒,例如猿猴病毒40(SV40)的侵染[14-16]。在细胞被病毒感染前用染料木黄酮处理1h后,观察到PICV(伯钦特沙粒病毒)核酸及其糖蛋白表达量相对于未处理细胞是减少的,这也许会对病毒感染起到抑制作用或者减少病毒的复制。染料木黄酮处理细胞不能改变Vero细胞的TfR(铁传递蛋白受体)的水平。噬菌斑测定分析显示,用染料木黄酮处理过的PICV感染的Vero细胞中的病毒浓度比未处理的或用DMSO处理的降低90%。辛德比斯病毒的浓度并不受染料木黄酮处理的影响[17]。此外,用台盼蓝染色法未观察到用100μmol·L-1染料木黄酮处理的Vero细胞具有毒性。
接下来研究了在Vero细胞受感染后注入染料木黄酮是否有抑制感染的作用。观察到当细胞刚开始受到药物处理与感染后接受药物处理比较,病毒浓度就下降了66.5%,当细胞受感染前用染料木黄酮处理30min后病毒浓度下降了56.9%,当细胞受感染前用染料木黄酮处理8h后病毒浓度下降了33.6%,当感染48h后在培养基中加入染料木黄酮时,病毒产率下降了73%。这些数据表明在被病毒感染后用染料木黄酮处理细胞可以抑制病毒的侵入。总而言之,不管是之前或之后用染料木黄酮处理细胞均可以减少PICV病毒感染的发生。
为了进一步证明染料木黄酮的处理对PICV感染的细胞的影响,研究了染料木黄酮对两种转录因子的影响,其中一个牵涉到PICV的感染过程,另一个作用于整体的酪氨酸使其磷酸化。分析受感染的Vero细胞全细胞提取物,染料木黄酮处理过的受感染细胞和未处理过的受感染细胞相比,磷酸酪氨酸的总体水平也减少了。有趣的是,磷酸化的细胞外信号调节激酶(p42和p44MAPK)没有被染料木黄酮处理所抑制;相反还出现了增长的现象。
为了证明染料木黄酮是否抑制了LASV(Old World沙粒病毒)的侵入,利用了复制缺陷型LASVMLV逆转录病毒的假模标本系统。标记基因在假模标本转导的Vero细胞中的表达受到染料木黄酮剂量不同的抑制。然而,LASV-MLV的转导却不因0.1% DMSO(染料木黄酮的载体)的处理而受到影响,这说明抑制作用是由药物引起的而不是载体。总之,以上结果显示LASV的侵入需要酪氨酸激酶活性。
PICV侵染培养的细胞诱导ATF-2磷酸化作用对染料木黄酮是敏感的。PICV在CREB(cAMP效应元件连接)过程中的侵染结果为转录因子CREB在DNA上与cAMP反应元件结合。这一蛋白存在于由PICV侵染诱导的细胞信号网络中[18]。染料木黄酮处理后丝氨酸磷酸化作用的减弱可能与上游活性物质减少或染料木黄酮介导的上游信号媒介物质的酪氨酸磷酸化作用受到抑制有关。ERK(细胞外信号调节激酶)1处理和ERK2磷酸化用是随着染料木黄酮的处理而增强的,而侵染是其它染料木黄酮介导的激活ERK路径效应的结果。然而,ERK磷酸化作用的增强证明了由染料木黄酮介导的激酶抑制作用有相当大的特异性。
4 染料木黄酮的抗肿瘤作用[19-20]
对癌细胞周期进展的控制是控制肿瘤生长的有效策略,就像对人类癌细胞的分子分析揭示了细胞周期调节器在多数正常的恶性肿瘤中是非常容易发生突变的。体外研究结果显示了用染料木黄酮处理雄激素敏感的(LNCaP)细胞可以让其在细胞周期的G1期停止,这说明染料木黄酮可能抑制细胞周期循环。在染料木黄酮处理后的LNCaP细胞中未找到任何显著的编程性细胞死亡诱导作用。染料木黄酮处理后细胞周期调节蛋白表达量的减少预示着这些细胞失控的细胞周期被破坏。染料木黄酮诱导的G1期停止是通过p21/CIP1表达的上调介导的,已报道染料木黄酮能使鼠的成纤维细胞、黑色素瘤细胞[21]、前列腺癌(LNCaP)细胞[22]在细胞周期的G0-G1停止。
用染料木黄酮处理雄激素不敏感(DuPro)细胞导致G2-M期细胞周期停止,并且在染料木黄酮浓度为25μmol·L-1时凋亡细胞数量显著增加。染料木黄酮能够诱导乳腺细胞、胃细胞、人类黑素瘤细胞[23]、PC3前列腺癌细胞和肺癌细胞在细胞周期的G2-M期停止。染料木黄酮处理雄性激素不敏感的PC3细胞使其在G2-M期停止并改变了两种细胞周期调节蛋白的表达,即细胞周期调节蛋白B1表达量的显著减少伴随着p21WAF1表达量的增加。细胞周期调节蛋白B1/CDK1复合物对细胞有丝分裂是必需的,所以,B1表达量的减少能够导致G2-M期的停止。
染料木黄酮能够上调某些基因mRNA的表达,例如,乳腺癌中的BRCA1基因,食管鳞状上皮细胞癌中的p16INK4a、RARb和MGMT28。剂量式染料木黄酮处理能够增加前列腺癌细胞株中肿瘤抑制基因p16和p21的mRNA和蛋白质的表达。在分析p16和p21启动子的CpG岛甲基化状态后,研究了这一通过后生途径的诱导机制。DNA甲基化分析发现两种细胞株中未经过任何染料木黄酮处理时,p21启动子也出现了缺失。然而,p16在染料木黄酮处理后,在DuPro细胞中完全甲基化。p21WAF1启动子在多数人类癌细胞中均未被甲基化[24]。在人类肺癌细胞、结肠癌细胞和用deacitabine处理后的急性髓细胞样白血病细胞株中均发现了未甲基化的 p21 WAF1启动子诱导的p21WAF1的表达[25-26]。此外,Shin22报道了p21基因在胃癌细胞中未被甲基化,这证实了甲基化不是其钝化的机制。已发现p16基因启动子在LNCaP细胞中未被甲基化而在DuPro细胞中完全甲基化。p21(LNCaP和 DuPro)和 p16 (LNCaP)CpG岛甲基化的缺失说明甲基化不是这些细胞株中染料木黄酮诱导表达的机制。芯片分析染色质重塑途径为前列腺癌中染料木黄酮的作用机制。剂量式染料木黄酮处理LNCaP、DuPro和RWPE细胞导致其转录起始位点组蛋白 H3、H4和H3dimethK4乙酰化的增加。在任何细胞株中均未发现p16和p21启动子组蛋白修饰的抑制。诱导p16和p21基因表达的是活性染色质修饰的强化而不是抑制组蛋白修饰的缺失。
为了进一步证明芯片数据的结果,用荧光定量PCR检测了HAT、p300、CREBBP、PCAF和HAT1的表达。结果证明了染料木黄酮对所有的HAT的表达有上调作用,这样也许能促进催化转录激活作用的组蛋白的乙酰化。据此观点,组蛋白超乙酰化和超乙酰化在致癌作用中似乎都是重要的,组蛋白超乙酰化能够诱导基因表达也许是高纤维饮食具有抗癌作用的机理。核小体组蛋白乙酰化的稳定状态已经由HAT和HDAC竞争的动态平衡建立了。大体来说,组蛋白赖氨酸残基的超乙酰化能促进转录激活作用,而脱乙酰基作用则使转录沉没[27-28]。近期的研究发现,许多转录激活因子有HAT活性,这支持了组蛋白乙酰化与转录激活作用有关这一观点。
举例来说,染料木黄酮能够抑制人类前列腺癌细胞运动性。Endoglin是一种重要的前列腺癌细胞运动性抑制基因,在前列腺癌发病时其表达是缺失的。因此确定endoglin的缺失是否对染料木黄酮的功效有影响是很重要的,Clarissa S.Craft首次发现了染料木黄酮通过激活endoglin伴随信号途径补偿了endoglin缺陷。实际上,endoglin激活了Smad1转录子的活性。这依次表现出需要ALK2,即激酶感受态ALK2。同时也发现endoglin抑制前列腺癌细胞活力通过一种机制,其包含类型 I TGFβ超家族受体ALK2和Smad1[29]。因此,目前鉴定ALK2-Smad1依赖性激活作用是正确的,该活性属于Smad1转录子活性,是通过染料木黄酮补偿的缺陷型endoglin结合的机制。除此之外,染料木黄酮在某些途径上的作用具有一定的特异性。特别是,染料木黄酮降低的是前列腺癌细胞的侵入而不是细胞的生存能力[30]。虽然人为改变endoglin表达能够影响细胞活力,但却不能影响其繁殖能力。最后,目前的研究表明,细胞繁殖能力受到密切关注,与其它相关条件相比,繁殖能力在实验条件下不会变化。概括来说,染料木黄酮能够诱导低endoglin前列腺癌细胞变异成低活力的高endoglin表型。这是由于染料木黄酮介导的Smad1激活作用,该作用依靠了具有激酶活性的ALK2。因为endoglin在前列腺癌发病过程中是缺失的,贡献给它的迁移表型,当前的研究支持一种观点,即拥有低endoglin前列腺癌细胞表达的个体可能从染料木黄酮治疗上获得更大的益处。这些发现有助于解释正在进行中的第二阶段的染料木黄酮在前列腺癌和其它癌症中的分子功效的研究。
5 染料木黄酮抗辐射损伤的作用
实验结果表明,染料木黄酮对UV所致损伤有防护作用。Moore[31]等研究了染料木黄酮对急性UVB照射所致人造皮肤损伤的防护作用。在照射前1h,分别给予人造皮肤10、20、50mmol·L-3染料木黄酮处理,然后分别给予20MJ·cm-2或60MJ·cm-2的照射,免疫组织化学分析法检测照射后6h和12h皮肤内增殖细胞核抗原和嘧啶二聚体的含量。结果显示,与空白对照组相比较,无论是给予20MJ·cm-2还是60MJ·cm-2剂量的照射,染料木黄酮处理组的嘧啶二聚体含量比对照组都明显降低,并且随着浓度的增加,嘧啶二聚体的含量降低越多;而增殖细胞核抗原的数量则随着染料木黄酮浓度的增加而增加,在空白对照组只观察到极少的增殖细胞核抗原。提示染料木黄酮可以减轻辐射所致的DNA损伤,促进受损细胞的增殖修复。
8-羟基脱氧鸟嘌呤(8-OH-dG)是鸟嘌呤的氧化产物,可导致DNA难以修复的GC和TA易位,是辐射诱导癌变和突变的重要基础。研究表明,UVB照射可使裸鼠皮肤和内脏的 H2O2、脂质过氧化物(MDA)和8-OH-dG的水平较未照射组升高了2~3倍,给予100mmol·L-1的染料木黄酮溶液(乙醇与丙烯乙二醇混合的体积分数为1∶1)涂抹,可降低H2O2和8-OH-dG的含量约50%,降低MDA的含量30%~50%[32]。Kruk等[33]的实验也观察到培养液中加入0.5mmol·L-1的染料木黄酮可有效地抑制羟自由基所致的脱氧核糖的降解,与对照组相比,可减少20%~60%的脱氧核糖降解。环氧化酶-2 (cyclooxygenase,Cox-2)是炎症过程中前列腺素(PG)合成的限速酶,是“快速反应基因”的表达产物,在细胞受到刺激后大量表达。近年来大量研究表明,Cox-2的表达与肿瘤的发生发展密切相关,一些抑制Cox-2表达的药物可以预防肿瘤的发生。染料木黄酮可以下调受照细胞Cox-2的mRNA异常表达,体外培养角质生成细胞的Cox-2的表达量在受照后较对照组升高3.3倍,加入染料木黄酮共培养后,其表达明显降低,仅为对照组的2.4倍。Wei等[34]的研究表明,染料木黄酮可以有效地阻止人体和裸鼠皮肤因UV损伤而诱发的光损伤和分子变异,对UV照射小鼠引起的皮肤癌和皮肤老化有直接的抑制作用,尤其在慢性实验中(每周2次,每次0.3kJ·m-2),局部应用染料木黄酮涂抹可以降低光致肿瘤的发生率近90%。
染料木黄酮还可以明显抑制UV照射诱发的皮肤炎症反应。小鼠涂抹20mmol·L-1的染料木黄酮,可降低其受照射后的皮肤炎症反应强度和减弱辐射引起的迟发型超敏反应(CHS)抑制,小鼠受照射后48h,染料木黄酮组的背脊中央皮褶厚度增加0.071~0.126mm,而对照组则平均增加了0.91mm;对照组CHS反应降低64%,染料木黄酮组仅降低了13%。染料木黄酮对UV和其他因素协同诱导的损伤也具有防护作用。临床上常用补骨脂素加UVA(PUVA)治疗白癜风等皮肤病,长期使用则会增加患鳞状细胞癌及恶性黑色素瘤的风险。Shyong[35]观察发现,染料木黄酮可显著降低PUVA诱发的小鼠炎症反应,减少皮肤红斑面积和溃疡的形成;组织学检查显示,局部涂抹染料木黄酮可使照后72h增殖细胞核抗原的数量由空白对照组的29%回升到40.7%,基本达到正常组的水平,同时还可完全抑制PUVA引起的细胞内多ADP聚合酶(PARP)降解产物及半胱天冬酶(caspase-3)活性的升高。
6 染料木黄酮对骨质疏松的预防作用
雌激素是女性的重要生殖激素,绝经期或绝经期后妇女体内雌激素水平降低,对成骨细胞的刺激减弱,使骨形成和骨吸收的平衡失调,骨吸收大于骨平衡,骨质逐渐丢失,骨密度减小,这是老年妇女易发生骨质疏松的重要病因。妇女绝经后适当补充雌激素,可预防骨质疏松的发生。大豆异黄酮是一类典型的植物雌激素,主要存在于大豆及其制品中。有报道证实亚洲骨质疏松相关骨病的发病率低于大多数西方人群,许多学者认为这与亚洲人群摄入豆制品多于西方有关[36-37]。
关于GEN对于成骨细胞的作用机制目前有多种解释,其中比较公认的说法是类雌激素作用。因为GEN的结构与雌激素非常相似,可以与成骨细胞上雌激素受体结合而发挥功效,雌激素是影响骨代谢的重要物质,对成骨细胞增殖、编码骨基质蛋白、激素受体、转录因子基因的表达基细胞因子等有直接作用。
ALP是成骨细胞主要功能活性酶,富含于胞浆中,在分化早期,ALP的活性反映了成骨细胞的成骨活性,ALP可水解有机磷酸释放出无机磷,相对增加局部无机磷浓度,用于羟磷灰石的形成,促进矿化过程,是骨形成的特异性酶。成骨细胞加药培养72h后测定其胞浆内的ALP活性,与对照组比较,各剂量组GEN都可增高成骨细胞的ALP活性,结果具有统计学意义(P<0.05)。同时随着GEN浓度的升高,ALP的数值也越高,具有一定的相关趋势(P<0.05)。说明10-8mol·L-1至10-5mol·L-1浓度的GEN可促进成骨细胞分化,并与GEN剂量相关。
[1]井乐刚,张永忠.微生物发酵制备大豆异黄酮的研究进展[J].微生物学通报,2003,30(2):86-88.
[2]张永忠,孙艳梅.大豆异黄酮研究中的名词术语[J].中国粮油学报,2004(4):46-52.
[3]Armin Littermann,Jujia Geppert,et al.Short term effect of high soy supplementation on sex hermones,bone markers and lipid parameters in young females adults[J].Eur J Nutr,2004,43:100-108.
[4]Lamartiniere C A,Moore J,Holland M,et al.Neonatal Genistein Chemoprevents Mammary Cancer[J].Proc Soc Exp Biol Med,1995,208(1):120-123.
[5]Song TT,Hendrich S,Murphy PA.Estrogenic Activity of Glycitein,a Soy Isoflavone[J].J Agric Food Chem,1999,47,1607-1610.
[6]Adams NR.Detection of the Effedts of Phytoestrogens on Sheep and Cattle[J].J Anim Sci,1995,73:1509-1515.
[7]陈学颖,张永忠.红三叶草异黄酮及其代谢研究进展[J].中草药,2007,38(11):1741-1744.
[8]张永忠,陈学颖,孙艳梅.四种异黄酮抗氧化活性的比较研究[J].食品科学,2008,29(2):383-386.
[9]Corinna ER,Sabine EK.Antioxidant Activity of Isoflavones and their Major Metabolites Using Different in Vitro Assays[J].J Agric Food Chem,2006,54:2926-2931.
[10]Schreiber M D,Liu J H,Subbiah M T,et al.Dietary inclusion of whole soy foods result s in significant reductions in clinical risk factors for osteoporosis and ardiovascular disease in normal postmenopausal women[J].Menopause,2001(8): 384-392.
[11]H P L EE,L Gourl EY,S W Duefy,et al.Dietary effect son breast cancer risk in Singapore[J].Lancet,1991,337:1197-1200.
[12]Noёl J-M Raynal,Louise Momparler.Antileukemic Activity of Genistein,a Major Isoflavone Present in Soy Products[J].J Nat Prod,2008,71:3-7.
[13]Eric M Vela,Gavin C,Bowick.Genistein treatment of cells inhibits arenavirus infection[J].Antiviral Research,2008,77: 153-156.
[14]Akiyama T,Ishida J,Nakagawa S,et al.Genistein,a specific inhibitor of tyrosinespecific protein kinases[J].J Biol Chem,1987,262:5592-5595.
[15]Damm E,Pelkmans L,Kartenbeck J,et al.Clathrin-and caveolin-1-independent endocytosis:entry of simain virus 40 into cells devoid of caveolae[J].J Cell Biol,2005,168:477-488.
[16] PelkmansL,PuntenerD,HeleniusA.Localactin polymerization and dynamin recruitment in SV40-induced internalization of caveolae[J].Science,2002,296:535-539.
[17]Lecot S,Belouzard S,Dubuisson J,et al.Bovine viral diarrhea virus entry is dependenton clathrin-mediated endocytosis[J].J Virol,2005,79:10826-10829.
[18]Bowick G,Fennewald S,Elsom B,et al.Differential signaling networks induced by mild and lethal hemorrhagic fever virus infections[J].J Virol,2006,80:10248-10252.
[19]Shahana Majid,Nobuyuki Kikuno.Genistein Induces the p21WAF1/CIP1 and p16INK4a Tumor Suppressor Genes in Prostate Cancer Cells by Epigenetic Mechanisms Involving Active Chromatin Modification[J].Cancer Res,2008,68:(8):2736-2744.
[20]Clarissa S.Craft,Li Xu,Diana Romero.Genistein Induces Phenotypic Reversion of Endoglin Deficiency in Human Prostate Cancer Cells[J].Molecular Pharmacology,2008,73:235-242.
[21]Kuzumaki T,Kobayashi T,Ishikawa K.Genistein induces p21(Cip1/WAF1)expression and blocks the G1 to S phase transition in mouse fibroblast and melanoma cells[J].Biochem Biophys Res Commun,1998,251:291-295.
[22]Shen JC,Klein RD,Wei Q,et al.Low dose gensitein induces cyclin dependent kinase inhibitors and G1 cell cycle arrest in human prostate cancer cells[J].Mol Carcinog,2000,29:92-102.
[23]Casagrande F,Darbon JM.p21CIP1 is dispensable for the G2arrest caused by genistein in human melanoma cells[J].Exp Cell Res,2000,258:101-108.
[24]Ying J,Srivastava G,Gao Z,et al.Promoter hypermethylation of the cyclin-dependentkinase inhibitor(CDKI)gene p21WAF1/CIP1/SDI1 is rare in various lymphomas and carcinomas[J].Blood,2004,103:743-746.
[25]Zhu WG,Srinivasan K,Dai Z,et al.Methylation of adjacent CpG sites affects Sp1/Sp3 binding and activity in the p21(Cip) promoter[J].Mol Cell Biol,2003,23:4056-4065.
[26]Schmelz K,Wagner M,Dorken B,Tamm I.5-aza-2-deoxycitidine induces p21(WAF)expression by demethylation of p73 leading to p53-independent apoptosis in myeloid leukemia[J].Int J Cancer,2005,114:683-695.
[27]Grozinger CM,Schreiber SL.Deacetylase enzymes:biological functions and the use of small molecule inhibitors[J].Chem Biol,2002,9:3-16.
[28] KristeleitR,StimsonL,WorkmanP,etal.Histone modification enzymes:novel targets for cancer drugs[J].Expert Opin Emerg Drugs,2004,9(1):135-154.
[29]Craft CS,Romero D,Vary CPH,et al.Endoglin inhibits prostate cancer motility via activation of the ALK2-Smad1 pathway[J].Oncogene,2007,26:7240-7250.
[30]Huang X,Chen S,Xu L,et al.Genistein inhibits p38MAP kinaseactivation,MMP-2 and cell in vasion in human prostate epithelial cells[J].Cancer Res,2005,65:3470-3478.
[31]Moore JO,Wang Y,Stebbins WG,et al.Photoprotective effect of isoflavone genistein on ultraviolet B induced pyrimidine dimer formation and PCNA expression in human reconstituted skin and its implications in dermatology and prevention of cutaneous carcinogenesis[J].Carcinogenesis,2006,27(6):367.
[32]Wei H,Zhang X,Wang Y,et al.Inhibition of UV lightinduced oxidative events in the skin and internal organs of hairless mice by isoflavone Genistein[J].Cancer Lett,2002,185(1):21.
[33]Kruk I,Aboul-Enein HY,Michalska T,et al.Scavenging of reactive oxygen species by the plant phenols genistein and oleuropein[J].The JournalofBiologicaland Chemical Luminescence,2005,20(2):81.
[34]Wei HC,Saladi R,Lu YH,et al.Isoflavone genistein: photoprotection and clinical implications in dermatology[J].Nutr,2003,133(11):3811.
[35]Shyong EQ,Lu Y,Lazinsky A,et al.Effects of the isoflavone 4,5,7-trihydroxy-isoflavone(Genistein)on peoralen plus ultraviolet A radiateon(PUVA)-induced photodramage[J]. Carcinogenesis,2002,23(2):317.
[36]Ikeda Y,Iki M,Morita A,et al.Intake of fermented soybeans natto is associated with reduced bone loss in postmenopausal women Japanese population-based osteoporosis study(JPOS)[J].J Nutr,2006,136:1323-1328.
[37]常红,黄国伟.染料木黄酮对成骨细胞I型胶原表达及转化生长因子 β1的影响[J].营养学报,2008,30(2): 194-198.
Research progress on biological activity and prevention disease mechanism of genistein
QU Yan-yan1,ZHANG Yong-zhong1,2,*
(1.College of Science,Northeast Agricultural University,Haerbin 150030,China; 2.Key Laboratory of Soy biological of Ministry of Education,Haerbin,150030,China)
Genistein is an isoflavone in the aglucone form.It has varieties of biological activity.The animal experiments,clinical researches and epidemiological investigations indicated that genistein has the effect for prevention and treatment of leukemia,viral infection,tumor,radiation protection and bone rarefaction precaution. Genistein is able to inhibit cellular and humoral immunity and regulate cell apoptosis.The biological activity of genistein and the mechanism of leukemia,viral infection,tumor and radiation prevention were reported.
genistein;leukemia;viral infection;tumor prevention;radiation protection;bone rarefaction
TS201.2
A
1002-0306(2010)08-0364-06
2009-03-12 *通讯联系人
曲妍妍(1984-),女,硕士研究生,研究方向:食品化学。
国家863课题(2008AA10Z331)。
