锰对水产养殖动物生长、免疫与抗应激的影响
2010-10-23陈晓丹冼健安苟妮娜王安利
陈晓丹, 冼健安, 苟妮娜, 王安利
(华南师范大学 生命科学学院, 广东省水产健康安全养殖重点实验室, 广东省高等学校生态与环境科学重点实验室, 广东 广州 510631)
锰对水产养殖动物生长、免疫与抗应激的影响
Effects of Manganese on growth, immunity and anti-stress of aquatic animals
陈晓丹, 冼健安, 苟妮娜, 王安利
(华南师范大学 生命科学学院, 广东省水产健康安全养殖重点实验室, 广东省高等学校生态与环境科学重点实验室, 广东 广州 510631)
随着我国水产养殖业集约化和规模化的发展,高密度养殖中投喂频率增加, 投饲量增大, 消毒剂和药物滥用等带来了养殖环境的不断恶化, 致使养殖水产养殖动物应激因素增多, 病害滋生, 严重阻碍水产养殖业的发展[1,2]。因而寻求节约减排、安全高效、抗逆抗病抗应激的无公害健康养殖模式[3]已成为研究热点。
锰(manganese, Mn)是多种酶的必需组分和激活剂, 对水产养殖动物维持生命活动和生长发育有重要作用[4]。近年来一些研究证实, 锰还在提高免疫力、抗应激能力方面有特殊的作用。本文结合国内外研究成果, 对锰在水产养殖动物生长、免疫和抗应激能力的作用及相关机制进行阐述, 以期为促进锰生物学功能的深入研究, 将其纳入无公害健康养殖模式提供参考。
1 锰对水产养殖动物生长的影响
1.1 锰对物质代谢的影响
锰是精氨酸酶、脯氨酸肽酶、丙酮酸羧化酶、核糖核酸聚合酶等的组成成分, 也是体内多种酶的激活剂, 可激活特定酶(如糖基转移酶等)或非特定酶(如激酶、水解酶、脱羧酶等)[5~7]。锰通过这些酶参与蛋白质、糖类、脂肪、核酸的代谢及维生素B、C、E的合成, 如 1分子丙酮酸羧化酶在 4个 Mn2+参与组成下, 可催化丙酮酸生成草酰乙酸, 打通丙酮酸异生为葡萄糖的道路[8]; 又如法尼酰转移酶以锰作为激活剂, 催化 牛儿焦磷酸与异戊烯焦磷酸合成法尼焦磷酸, 进而合成胆固醇[8,9]。
锰对水产养殖动物的消化酶活性具有一定作用。在5×10-3mol/L浓度下, Mn2+可明显激活大菱鲆(Scophthalmus maximus)的肠蛋白酶[10]、三疣梭子蟹(Portunus trituberculatus)的蛋白酶、淀粉酶和纤维素酶, 并明显抑制三疣梭子蟹的羧肽酶 A、B, 而对日本对虾(Marsupenaeus japonicus)和三疣梭子蟹的胰蛋白酶活性则无显著性影响[10~12]; 在 1×10-3mol/L浓度下, Mn2+可明显激活三疣梭子蟹的淀粉酶、纤维素酶、蛋白酶、胰蛋白酶和胰凝乳蛋白酶, 并明显抑制其羧肽酶A、B和斑节对虾(Penaeus monodon)的胰凝乳蛋白酶[12,13]; 在5×10-4mol/L浓度下, Mn2+可明显激活三疣梭子蟹的淀粉酶、纤维素酶和羧肽酶 A,同时明显抑制羧肽酶 B的活性[12]。关于中国对虾(Penaeus chinensis)的实验也发现, 羧肽酶A的活性随饲料锰增加而上升, 在锰添加量为80 mg/kg时达最高值, 在高于80 mg/kg时则下降[14]。可见消化酶对锰含量具有一个适宜的浓度范围; 对于不同生物种类体内的同一种消化酶或同一生物体内的不同消化酶, Mn2+的作用效果并不完全一致, 且不同浓度Mn2+表现效果不同。
消化酶的活性与营养物质消化利用率、机体的生长发育相关, 但目前锰对水产养殖动物生长的研究多停留在饲料系数、增重率、体长增长率、肥胖度等宏观层面, 由于水产养殖动物的特殊性及残饵难以准确测定, 因而很难得出可靠的饲料系数。若将宏观指标和微观指标如一些消化酶的活性有机结合,可更全面、准确地反映水产养殖动物的生理状况。
1.2 锰对骨骼形成的影响
锰是骨正常形成所必需的元素。它可激活多糖聚合酶和半乳糖转移酶, 催化二磷酸尿苷-N-乙酰-半乳糖胺与二磷酸尿核苷-葡萄糖醛酸结合成多糖,对骨骼基质的硫酸软骨素的形成起重要作用。硫酸软骨素是骨有机基质黏多糖的组成成分, 其合成受阻将严重影响软骨的成骨作用, 甚至导致骨骼畸形[5,6,8]。虹鳟(Salmo gaidnerii)和鲤鱼(Cyrinus carpio)在饲料中缺锰或锰不足(4 mg/kg)时尾部畸形, 鱼体变短, 而在锰含量添加至 12~13 mg/L时虹鳟的畸形受遏止[15,16]。这说明锰对鱼类骨骼的正常发育有重要作用。
此外, 锰在鱼类骨骼中具较高含量[6,17]。这是由于在一定程度上, 脊椎骨相当于机体的锰元素库,可在饲料锰不足时部分释放锰供机体维持正常的生命活动[18]。鲤鱼、斑点叉尾(Ictalurus punctatus)、牙鲆(Paralichthys olivaceus)和黄颡鱼(Pelteobagrus fulvidraco)脊椎骨的锰含量在饲料缺锰时较低, 而随饲料锰的增加而不断上升, 斑点叉尾的甚至呈线性上升[6,18,19]。因此, 可将水产养殖动物骨骼中的锰含量作为评价饲料中锰是否充分的敏感指标。
1.3 锰对生长的影响
研究发现, 日本对虾仔虾在海水锰为40 µg/L时生长最好[20]; 虹鳟和鲤鱼在饲料中缺锰或锰不足(4 mg/kg)时生长缓慢; 锰含量添加至 12~13 mg/L时虹鳟的生长加速[15,16]。水产养殖动物锰的需求量因种类不同而异(表 1), 研究结果的差异与实验方法和实验条件, 特别是与饲料组成、虾体大小、水体物理和化学条件等因素的差异有关。提高锰生长需求量研究的准确性还有赖于生长指标、实验技术、作用机理等的不断完善。
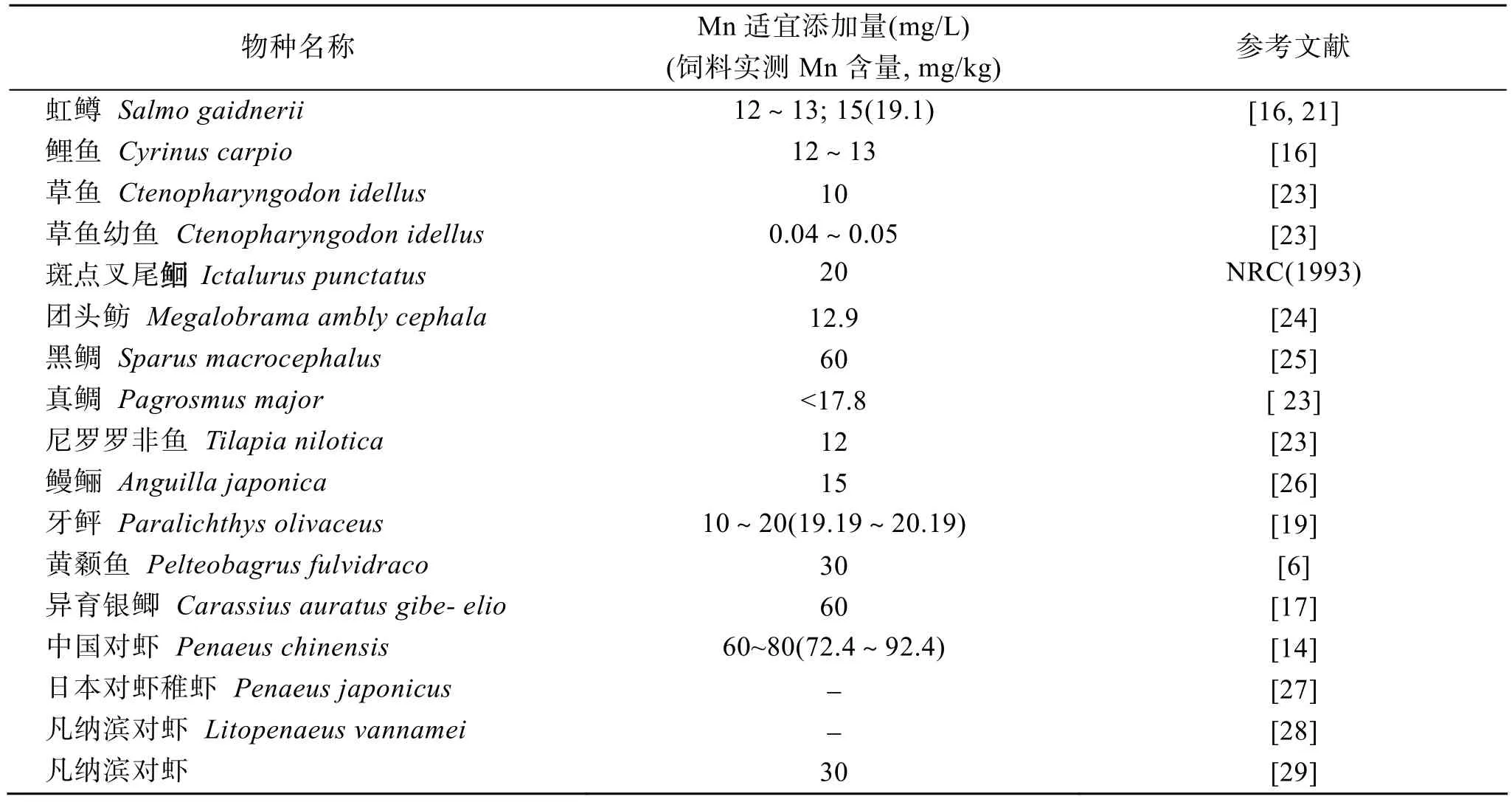
表1 水产养殖动物饲料锰的适宜添加量Tab. 1 Optimal Mn supplement in the diet of aquatic animals
2 锰对水产养殖动物免疫的影响
2.1 锰对细胞免疫的影响
鱼类的细胞免疫主要通过淋巴细胞(主要是T细胞)和吞噬细胞(单核细胞、巨噬细胞、粒细胞)两大类免疫细胞进行[30]。对于淋巴细胞, 锰一方面通过对碱性磷酸酶(alkaline phosphatase, AKP)活性的作用影响其自身代谢、吞噬和吞饮, 另一方面通过以下三种途径影响其增殖: 1)锰可激活鸟苷酸环化酶产生CGMP, 选择性促进T细胞增殖反应并协同增强其免疫功能; 2)Mn2+可与ATP的3个磷酸基结合生成Mn-ATP, Mn-ATP是腺苷酸环化酶(adenosine cyclophosphate cAMP)的底物, 而高浓度cAMP可抑制T淋巴细胞活化增殖; 3)适量Mn2+(1~10 µmol/L)时可满足淋巴细胞增殖反应的需求, 但浓度过高(100 µmol/L)时, Mn2+与 Ca2+竞争钙调蛋白(CaM)的结合位点, 使淋巴细胞内CaM-Ca2+含量下降, 干扰CaM的正常调节, 从而抑制T细胞的活化增殖[31,32]。对于吞噬细胞,锰盐抑制趋化性, 影响嗜中性白细胞对氨基酸的吸收; 增加锰可提高非特异性免疫中酶的活性和巨嗜细胞的杀伤力; 缺锰则会造成白细胞机能障碍和机体特定免疫力下降。此外, 锰与刀豆素 A有丝分裂原有相互作用, 低浓度锰可提高淋巴细胞对刀豆蛋白的反应性, 但作用机制尚不清楚[5,33]。
甲壳动物的细胞免疫主要是血细胞的免疫作用[34], 血细胞吞噬外来病原微生物的过程中,NADPH酶被激活, 糖酵解反应增强, 导致大量活性氧(如 O2-、H2O2等)产生[35~37]。尽管活性氧杀菌效果很强, 但其过量积累会损害细胞。甲壳动物主要利用锰-超氧化物歧化酶(manganese superoxide dismutase,Mn-SOD)、过氧化氢酶(catalase, CAT)和谷胱甘肽过氧化物酶(glutathione peroxidase, GPX)将活性氧转化为对机体无害的 H2O与O2[9,35~37]。锰对这三种酶具一定的激活作用[40,41]。可见, 锰在一定程度上有利于甲壳动物维持正常的细胞免疫。
2.2 锰对体液免疫的影响
2.2.1 特异性免疫因子
抗体是脊椎动物淋巴细胞产生的能与相应抗原结合的, 具免疫功能的一类球蛋白。目前研究表明鱼类中软骨鱼仅IgM和IgMG两种抗体, 而真骨鱼可能仅IgM一种抗体[30]。关于锰对鱼类抗体的影响研究较少, 但已有资料表明锰与抗体的合成有密切关系。锰可提高某些动物体内抗体的效价, 缺乏或过量都会抑制抗体产生[5,17], 但其作用机制尚未阐明。
2.2.2 非特异性免疫因子
2.2.2.1 超氧化物歧化酶
SOD 与金属元素(铜、锌、铁、锰)结合可催化O2-生成H2O2, 是机体内关键性的抗氧化酶, 可作为水产养殖动物机体的免疫性指标[38,42,43]。在SOD的多种形式中, 以 Mn-SOD的状态最稳定, 半衰期最长, 该酶主要位于鱼类线粒体和甲壳动物的细胞质中, 线粒体与细胞质中的 Mn-SOD由不同基因编码[44], 目前这两种基因已被克隆、定位和表达[42,43]。
Mn-SOD以锰为活性中心, 缺锰会导致其结构稳定性降低[41,45]。用锰不足的饲料喂养虹鳟, 其心肌、肝脏中的CuZn-SOD和Mn-SOD活力下降[23]; 据不同生长阶段添加适量的饲料锰可使凡纳滨对虾肌肉中 Mn-SOD的活性显著上升[46]; 在多个浓度饲料锰的刺激下, 日本沼虾(Macrobrachium nipponense)肌肉中SOD呈剂量效应的激活, 且在锰为150 µg/g时其活性达到最高[41]; 适量的饲料锰(260 mg/kg)在一定时间内对中华米虾(Caridina denticulata sinensis)体内 SOD的激活作用显著, 但锰超过一定浓度,SOD活性就会降低, 随着养殖时间的延长, 实验组虾体SOD的活性反而低于对照组, 说明饲料中添加锰表现出明显的剂量效应和时间效应[32]。此外, 适量的锰还可提高异育银鲫(Carassius auratus gibeelio)血清、黏液和肝胰脏中SOD的活性, 尤其是Mn对黏液T-SOD活性的影响甚至大于Cu、Zn、Fe[7]; 黄颡鱼的实验结果也与之相近[6]。其作用机制可能是少量锰进入体内, 使机体的脂质过氧化作用增强, 其代谢产物丙二醛(MDA)增加, 机体内的自稳机制会使体内抗氧化能力出现代偿性增强, 使 T-SOD活性代偿性增高[32]。
2.2.2.2 谷胱甘肽过氧化物酶
GPX是清除羟自由基的重要抗氧化酶, 它通过催化 H2O2分解为 H2O和 O2来防止 OH-产生, 并将脂氢过氧化物(OROH)还原成相应 ORH, 从而减轻细胞膜多不饱和脂肪酸的过氧化作用, 以及 OROH分解产物所引起的细胞损伤[9,41,47]。在多个浓度饲料锰的刺激下, 日本沼虾肌肉中GPX和SOD趋势相似,呈剂量效应的激活, 且均在锰浓度为 150 µg/g时活性最高[41]; 凡纳滨对虾实验也发现各生长阶段 SOD和GPX被显著激活的最适锰浓度也一致[46]。这说明GPX与SOD有协同作用, SOD活性的显著升高可能伴随GPX的升高来清除过量生成的H2O2[48]。
2.2.2.3 过氧化氢酶
CAT是初级抗氧化酶系统的主要成分, 与GPX一样能清除 H2O2。组织中的过氧化物主要由 GPX来催化, 而在H2O2浓度高的情况下, 则由CAT催化[35,49]。CAT与SOD也有协同作用, SOD活性的显著升高可能会伴随CAT的升高来清除高浓度H2O2。在多个浓度饲料锰刺激下, 日本沼虾肌肉中 CAT的活性也伴随肌肉中SOD被显著激活而提高并呈明显剂量效应的激活, 且二者在锰浓度为 150μg/g时活性均最高[41]。
2.2.2.4 碱性磷酸酶
AKP是一种非特异性磷酸水解酶, 能催化磷酸单脂的水解及磷酸基团的转移反应, 对水产养殖动物吸收水中钙质、形成磷酸钙、分泌及形成甲壳素具有重要作用[50]。据报道, 海水中添加不同浓度锰均能提高AKP酶的活性; 根据不同生长阶段添加适量的饲料锰对凡纳滨对虾的AKP活力均有显著的激活作用[30]; 中国对虾仔虾和日本对虾仔虾的AKP活性分别在海水中锰浓度为20 µg/L和40 µg/L时达到最高, 锰浓度太高则抑制其活性[53,54]。
2.2.2.5 溶菌酶
溶菌酶(lysozyme, LSZ)通过酶解病原体细胞壁的黏多糖将其杀死, 是抵抗细菌病毒感染的非特异性免疫调节因子, 其水平和活性直接关系到水产养殖动物的免疫力和健康[52,53]。据不同生长阶段添加适量的饲料锰可显著激活凡纳滨对虾的 LSZ活力[46]; 50mg/kg的锰能使黄颡鱼血清中LSZ的活力最高[6]; 锰为60mg/kg时异育银鲫黏液、血清中的LSZ活力最高, 且前者明显高于后者。这可能是由于黏液作为鱼体的第一道免疫屏障, 活力较高的黏液 LSZ对水体环境细菌具较强的杀灭和抵抗作用[17]。
3 锰对水产养殖动物抗应激的影响
水产养殖动物长期处于应激状态会使其免疫系统受抑制, 易受病原体侵染, 所以如何增强水产养殖动物的抗应激能力来抵抗日益恶化的养殖环境对水产养殖业的健康发展起重要作用[53,54]。抗氧化酶与水产养殖动物的抗应激能力密切相关。对虾感染白斑综合征病毒后, 体内CAT等多种抗氧化酶活性发生变化[55,56]; 给对虾饲喂免疫增强剂后, 其体内SOD的抗氧化活性上升, 且感染细菌病毒后的存活率升高[38,57]。这提示CAT、SOD可能参与对虾抵御病原的免疫反应, 可能是通过及时清除活性氧来保障免疫细胞内氧化还原反应的动态平衡、信号的正常传导及免疫相关基因的正常转录表达, 从而提高对虾对病原的抵抗能力[58]。
锰在应激条件下对抗氧化酶有一定作用。以不同锰添加量的饲料投喂日本沼虾 30d后低氧胁迫组肌肉中 SOD、GPX、CAT的活性均相对较高, 且均随锰浓度的增加而呈先升后降的变化趋势, 在锰为150 µg/g时达到最高值[46]; 在不同浓度多聚磷酸钠胁迫下, 饲料有锰(150 µg/g)组的中华米虾的SOD、活性也始终比无锰组的要高。这表明适量的饲料锰可在一定程度上激活抗氧化酶, 增强机体对环境胁迫的抵抗能力[59]。
4 问题与展望
综上所述, 锰是多种酶的必需组分和激活剂,需求量不多, 但对水产养殖动物的生长、免疫和抗应激有重要的影响。目前, 关于锰的系统研究较有限,部分生长免疫作用机理尚不十分清楚, 对抗应激影响的研究还处于起步阶段, 许多问题还有待进一步研究, 如锰对甲壳动物的壳是否具有影响、作用机制、锰与补体的关系、生长免疫的综合需求量以及在各种应激因素作用下锰的需求量等。随着分子生物学和生物化学技术的发展, 有关锰与基因表达与调控机理的研究应引起营养学工作者的广泛关注。
[1] 徐君卓. 水产养殖业面临的新挑战[J]. 现代渔业信息, 2008, 23(1): 5-8.
[2] 赵明军, 黄志斌. 无公害渔药研发面临的问题和急需研究的课题[J]. 中国渔业经济, 2008, 3: 52-60.
[3] 王安利, 廖绍安. 生态养殖与环保饲料[J]. 现代渔业信息, 2008, 23(4): 3-8.
[4] S Ahamad Ali. Copper, Manganese and Zinc Requirements in the Diet of Shrimp Penaeus indicus[J]. Asian Fisheries Science, 2000, 13: 201-207.
[5] Takeshi Watanabe, Viswanath Kiron, Shuichi Satoh.Trace minerals in fish nutrition[J]. Aquaculture, 1997,151: 185-207.
[6] 蒋蓉. 铜、铁、锰、锌对黄颡鱼生长和生理机能的影响[D]. 苏州: 苏州大学, 2006.
[7] 倪可德. 动物微量元素营养(一)[J]. 粮食与饲料工业,1996, 2: 28-32.
[8] 于晶锋. 无角多赛特肉羊与蒙古羊铜、锌、锰、SOD、羟自由基及细胞免疫动态的比较研究[D]. 呼和浩特:内蒙古农业大学, 2006.
[9] 王镜岩, 朱圣庚, 徐长发, 等. 生物化学 [M]. 第三版. 北京: 高等教育出版社, 2002. 102-103, 252 -253.
[10] 王海英, 薛长湖, 孙谧, 等. 大菱鲆肠道蛋白酶的分离纯化及性质的初步研究[J]. 水产学报, 2005, 29(5):624-629.
[11] Galgani F G, Benyamin Y, Van Wormhoud.. Purification,properties and immunoassay of trypsin from the shimp Penaeus japonicus[J]. Compare Biochemic Physiol,1985, 81B: 447-452.
[12] 胡毅, 潘鲁青. 10种金属离子对三疣梭子蟹中肠腺消化酶的初步研究[J]. 中国海洋大学学报(自然科学版),2006, 25(6): 52-57.
[13] Jiang S T, Michael W. Purification and characterization of Proteases from Digestive Tract of Grass Shrimp(Penaeus monodon)[J]. Journal of Food Scince,1991, 56(2): 322-326.
[14] 李荷芳, 郝斌, 刘发义, 等.饵科中添加锰对中国对虾的影响[J]. 海洋科学, 1993, 7(4): 48-54.
[15] 佐藤秀一. 关于鱼类对微量元用特性的研究[J]. 饲料研究, 1997, 4: 14-17.
[16] Ogino C, Yang G Y. Requirements of carp and rainbow trout foe dietary manganese and capper[J]. Nippon Suisan Gakkaishi, 1980, 46: 455-458.
[17] 郭建林, 叶元土, 蔡春芳, 等. 日粮中添加Fe、Cu、Mn、Zn对异育银鲫生长及其形体的影响[J]. 江苏农业学报, 2009, 25(1): 154-159.
[18] Gatlin D M Ⅲ, Wilson R P. Studies on the manganese requirement of fingerling channel catfish[J]. Aquaculture, 1984b, 41: 85-92.
[19] 魏万权, 李爱杰, 李德尚. 牙鲆幼鱼饲料中锰、钴适宜添加量的初步研究[J]. 浙江海洋学院学报, 2001,20(增刊): 83-87.
[20] 刘存岐, 王安利, 王维娜. 海水中Zn2+和Mn2+对日本对虾仔虾体内碱性磷酸酶活性的影响[J]. 水产科技情报, 2002, 29(5): 195-197.
[21] Satoh S, Takeuchi T, Watanabe T. Availability of manganese and magnesium contained in white fishmeal to rainbow trout Oncorhynchus mykiss[J]. Nippon Suisan Gakkaishi, 1991, 57: 99-104.
[22] 黄耀桐, 刘永坚. 草鱼种无机盐需要量之研究[J]. 水生生物学报, 1989, 13(2): 134-151.
[23] 李爱杰. 水产动物营养与饲料学[M]. 北京: 中国农业出版社, 1996. 56-65.
[24] 赵振伦, 高国富. 团头鲂鱼体矿物质成分的初步分析[J]. 南京农业大学学报, 1994, 17(4): 71-76.
[25] 李荷芳, 王辉亮, 梁海德, 等. 黑鲷对Ca, P, K, Mg和Mn的营养需求[A]. 中国科学院海洋研究所. 海洋科学集刊[C]. 1996. 135-141.
[26] 吴锐全, 肖学铮. 鳗鲡的营养需求与饲料配制[J]. 饲料研究, 1999, 7: 1-3.
[27] Kanazawa A, S Teshima, M Sasaki. Requirements of potassium, copper, manganese and iron[J]. Memoirs of Faculty of Fisheries Kagoshima University, 1984,33(1): 63-71.
[28] Davis D A, Lawrence A L, Gatlin D M Ⅲ. Evaluation of the dietary iron requirement of Penaeus vannamei[J].Journal of word Aquaculture Society, 1992a, 23(1):15-22.
[29] 董晓慧, 周歧存, 郑石轩, 等. 锰源和锰水平对南美白对虾生长性能和组织锰含量的影响[J]. 中国饲料,2005, 9: 29-31.
[30] 张艳秋, 詹勇, 许梓荣. 鱼类免疫机制及其影响因子[J]. 水产养殖, 2005, 26(3) : 1-5.
[31] 刘凤娟, 黄贤仪, 张月明, 等. 锰对细胞免疫功能的影响[J]. 环境与职业医学, 2002, 19(3): 139-142.
[32] 王宏伟, 郭立格, 赵建华, 等. 饲料中锰对中华米虾SOD 活性的影响[J]. 河北大学学报(自然科学版) ,2006, 26(6): 649-653.
[33] 熊正儒, 肖学梅, 郑云林. Mn对畜禽的作用[J]. 中国饲料, 1997, 14: 31-32.
[34] Erin J Burge, Daniel J Madigan, Louis E Burnett, et al.Lysozyme gene expression by hemocytes of Pacific white shrimp, Litopenaeus vannamei, after injection with Vibrio[J]. Fish Shellfish Immunol, 2007, 22:327-339.
[35] Holmblad T, Söderhäll K. Cell adhesion molecules and antioxidative enzymes in a crustacean, possible role in immunity[J]. Aquaculture, 1999, 172: 111-123.
[36] Xiang J H. Disease occurrence and control strategies of mariculture organisms[J]. Ocean Press, 2001, 1-4,78-83.
[37] Roch P. Defense mechanisms and disease prevention in farmed invertebrates[J]. Aquaculture, 1999, 172(1-2):125-145.
[38] Campa-Córdova A I, Hernández-Saaverdra N Y, De Philippis R, et al. Generation of superoxide anion and SOD activity in haemocytes and muscle of American white shrimp (Litopenaeus vannameii)as a response to β-glucan and sulphated polysaccharide[J]. Fish Shellfish Immunol, 2002, 12: 353-366.
[39] Pipe R, Porte C, Livingstone D. Antioxidant enzymes associated with the blood cells and haemolymph of the mussel Mytilus edulis[J]. Fish Shelfish Immunol,1993, 3: 221-233.
[40] 刘存歧, 王伟伟, 张亚娟. 水生生物超氧化物歧化酶的酶学研究进展[J]. 水产科学, 2005, 24(11): 49-52.
[41] 王宏伟, 曹向可, 钱庆增, 等. 饲料中锰对日本沼虾抗氧化酶活性的影响[J]. 河北大学学报(自然科学版),2008, 28(3): 300-304.
[42] Yao C L, A L Wang. Purification and partial characterization of Mn superoxide dismutase from muscle tissue of the shrimp Macrobrachium nipponense[J].Aquaculture, 2004, 241(1-4): 621-631.
[43] Zhang Q L F, H Li. The mitochondrial manganese superoxide dismutase gene in Chinese shrimp Fenneropenaeus chinensis: Cloning, distribution and expression[J]. Developmental and Comparative Immunology, 2007, 31(5): 429-440.
[44] Marius B, Hoexum Brouwer T, Grater W, et al. The paradigm that alloxygen-respiring eukaryotes have cytosolic CuZn-superoxidant dismutase and that Mn-superoxidant dismutase is loealized to the mitoehondria doesnot marinearthopods[J]. Bioehemistry,1997, 36(43): 133-138.
[45] 于晶锋. 无角多赛特肉羊与蒙古羊铜、锌、锰、SOD、羟自由基及细胞免疫动态的比较研究[D]. 呼和浩特:内蒙古农业大学, 2006. 15-32.
[46] 许安阳. 硒钴锰对南美白对虾不同生长阶段免疫机能的影响[D]. 广州:华南师范大学, 2005.
[47] 黄志坚, 林藩平, 邱承亮, 等. 富硒酵母对奶牛抗氧化能力和免疫功能的影响[J]. 营养学报, 2004, 26(1):27-30.
[48] Warner H R. Superoxide dismutase, aging and degenerative disease[J]. Free Redical Biol Med, 1994, 17:139-162.
[49] Yu B P. Cellular defenses against damage from reactive oxygen species[J]. Physiol Rev, 1994, 74(1): 139-162.
[50] 陈清西, 陈素丽, 朱凌翔, 等. 长毛对虾碱性磷酸酶功能基团的研究[J].. 厦门大学学报(自然科学版),1996, 35(4): 587-591.
[51] 刘存岐, 王安利, 王维娜, 等. 海水中几种金属离子对中国对虾幼体体中碱性磷酸酶和ATPase的影响[J].水产学报, 2001, 25(4): 298-303.
[52] Hikima S, Hikima J, Rojtinnakorn J, et al. Characterization and function of kuruma shrimp lysozyme possessing lytic activity against Vibrio species[J]. Gene,2003, 316: 187-195.
[53] Chrousos G P, Gold P W. The concepts of stress and stress system disorders. Overvi- ew of physical and behavioural homeostasis[J]. JAMA, 1992, 267(9):1 244-1 252.
[54] Laurence Mercier, Elena Palacios, Ángel I.Campa-Córdova, et al. Metabolic and immune responses in Pacific whiteleg shrimp Litopenaeus vannamei exposed to a repeated handling stress[J].Aquaculture, 1996, 258: 633-640.
[55] Chang C F, Su M S, Chen H Y, Liao I C, et al. Dietary beta-1, 3-glucan effectively improves immunity and survival of Penaeus monodon challenged with white spot syndrome virus[J]. Fish Shellfish Immunol, 2003,15(4): 297-310.
[56] Campa-Córdova A I, Hernandez-Saavedra N Y, De Philippis R. Ascencio F. Superoxide dismutase as modulator of immune function in American white shrimp(Litopenaeus vannamei)[J]. Comp Biochem Physiol C Toxicol Pharmacol, 2002, 133(4): 557-565.[57] Mohankumar K, Ramasamy P. White spot syndrome virus infection decreases the activity of antioxidant enzymes in Fenneropenaeus indicus[J]. Virus Res,2006a, 115(1): 69-75.
[58] Mathew S, Ashok Kumar K, Anandan R, et al. Changes in tissue defence system in white spot syndrome virus(WSSV)infected Penaeus monodon[J]. Comp Biochem Physiol C Toxicol Pharmacol, 2007, 145(3): 315-320.
[59] 王宏伟, 王建芳, 马志刚, 等. 多聚磷酸钠胁迫下锰对中华米虾 SOD活性的影响[J]. 四川动物, 2008,27(5): 773-776.
S963.1
A
1000-3096(2010)10-0108-06
2010-03-02;
2010-06-01
国家自然科学基金资助项目(30671628); 广东省教育部产学研结合资助项目(2006D90204005); 国家科技支撑计划项目(2007BAD29B06); 广东省科技计划项目(2007B020708013);广东省重大科技专项(20090204); 广州市科技支撑计划项目(2009Z1-E681)
陈晓丹(1986-), 女, 广东汕头人, 硕士研究生, 从事水生动物营养免疫学与环境营养学研究, E-mail: youxincxd@126.com; 王安利,
, E-mail: wanganl@scnu.edu.cn
(本文编辑: 康亦兼)
