三峡水库蓄水前后春季长江口鱼类浮游生物群落结构特征
2010-09-05刘淑德线薇微
刘淑德,线薇微
(1.中国科学院海洋研究所海洋生态与环境重点实验室,山东青岛 266071;2.中国科学院研究生院,北京 100049)
三峡水库蓄水前后春季长江口鱼类浮游生物群落结构特征
刘淑德1,2,线薇微1
(1.中国科学院海洋研究所海洋生态与环境重点实验室,山东青岛 266071;2.中国科学院研究生院,北京 100049)
根据1999年、2001年、2004年和2007年春季长江口及其领近海域40个站位鱼类浮游生物拖网调查资料,采用相似性分析,探讨了长江口及其邻近海域春季鱼类浮游生物的组成、群落结构及生物多样性。分析结果表明:三峡水库蓄水后的2004年长江口春季鱼类浮游生物丰度迅速下降,仅为1999年的13.9%和2001年的4.3%,2007年略有回升;鱼类浮游生物群落结构年度间差异显著,2004年鱼类浮游生物群落结构与1999年和2001年相异性最高,?和凤鲚种群数量变动对其群落变异的贡献率最大,群落相似性以2004和2007年最高,达71.17%;与蓄水前相比,蓄水后的长江口鱼类浮游生物群落多样性也显著下降。
长江口;鱼类浮游生物;群落结构;生物多样性
1 概 述
长江口区是我国最大的河口渔场。它西靠大陆,有长江入海,北接吕泗渔场,南临著名的舟山渔场,水产资源丰富。长江口水域既是大黄鱼、小黄鱼、带鱼和银鲳等经济种类的重要产卵场和育幼场,也是夏秋季银鲳、刀鲚、凤鲚、带鱼、石首鱼类以及鲐、等中上层鱼类的重要索饵场;又是名贵鱼类鲥、松江鲈、中华鲟溯河或降海洄游的必经水道[1]。这些注定了长江河口区鱼类浮游生物种类组成的复杂性和多样性,也注定了长江口在鱼类浮游生物生态学研究中的重要地位。
近年来,随着长江流域经济发展和人口增加,人类对流域资源与环境的利用愈来愈多,环境污染的加剧最终给长江口及邻近海域的环境造成巨大压力。同时,长江流域的筑库建坝、截流引水,改变了入海径流和泥沙输运原有的季节性与年际变化的格局。河口环境因子的改变,势必影响到生态系统中生物群落的分布格局。其中,鱼类浮游生物(包括鱼卵和仔稚鱼)资源作为鱼类的补充资源,其群落结构和数量变动直接影响了该水域鱼类资源的生存与延续。
三峡水利枢纽工程于2003年6月完成一期工程蓄水(坝前达135 m),蓄水过程中拦截了长江上游的大量来水来沙,对河口及其邻近海域的生态与环境带来一定程度的影响[1]。本文根据1999年5月、2001年5月、2004年5月和2007年5月三峡水库蓄水前后4个春季航次鱼类浮游生物拖网调查资料,探讨长江口及其邻近海域春季鱼类浮游生物的种类组成、群落结构及生物多样性等,以期为长江口的海洋生物资源保护、合理利用以及科学评价三峡工程的生态环境效应提供重要依据。
2 材料和方法
2.1 调查方法
调查船为底层双拖网渔船(300HP),于1999年5月、2001年5月、2004年5月和2007年5月按照《海洋调查规范》(GB12763.1-7-91),采用大型浮游生物拖网(网长280 cm,网口内径80 cm,网口面积0.5 m2)在30°45′至32°N之间、123°20′E以西长江口水域40个站点进行水平和垂直取样(图1)。水平拖网10 min,垂直拖网由底层到表层。所得样品保存于5%的福尔马林的溶液中。在实验室内采用生物解剖镜,对各站鱼类浮游生物样品进行种类鉴定、个体计数和发育阶段的判别。
2.2 分析方法
首先对种类-丰度调查数据进行筛选,将调查数据中,仅在一个站位出现且丰度小于2%的物种去除,因为稀有物种没有可辨别的分布也没有明确的丰度类型,并且它们的存在对群落结构只提供很少的信息[2]。不同年份间鱼类群落结构组成的差异采用Bray-Curtis相似性指数[3]:
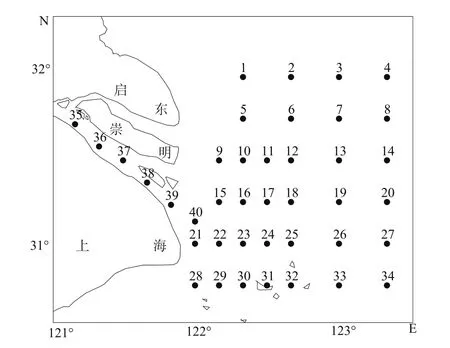
图1 长江口浮游植物生物群落调查站位图Fig.1 Location of survey stations in the study area for ichthyoplankton in the Changjiang River estuary
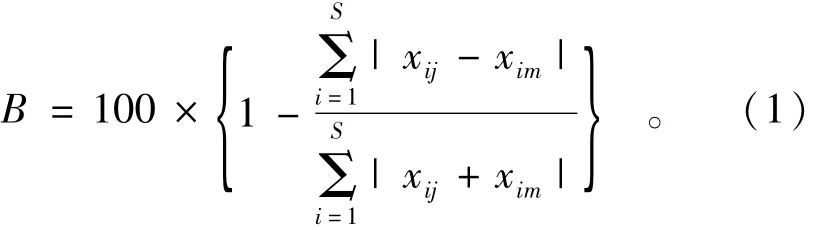
式中:S为4次调查中的总种类数;xij和xim分别为第i种类在j年份和m年份的平均单位网次捕获量,xij和xim需经4次方根转换,这样能够消除一些物种极端丰度的优势。
采用百分比相似性分析(SIMPER),计算引起年际间群落差异种类的贡献率。相似性分析检验(ANOSIM)用来检验年际间的相似性程度,R值越接近0,相似性程度越高;R值越接近1,差异越大。
采用相对重要性指数(IRI)计算鱼类群落优势种成分。

式中:N%为水平与垂直网中的个体数量比例之和;F%为水平和垂直网中频次比例之均值。本文选取IRI值大于100的种类为优势种,IRI值大于10的种类为常见种,小于10的为稀有种。
Margalef种类丰富度指数
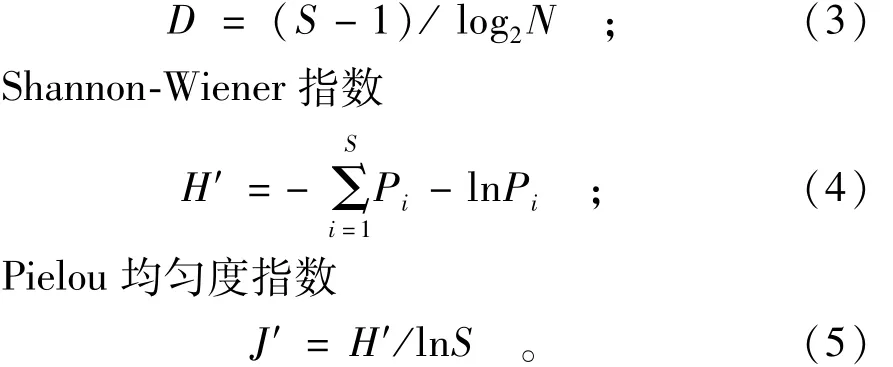
式(3)~(5)中:S为种类数;N为总尾数;Pi为i种鱼占总渔获量的比例。
全部数据的统计分析用SPSS 12.0软件完成,平面分布图用SURFER 8.0软件绘制,群落结构分析使用PRIMER 5.0软件完成。
3 结 果
3.1 种类组成的变化
4个航次共获得鱼卵1 883粒,仔稚鱼10 726尾,隶属11目21科39种,其中以鲈形目和鲱形目种类最多。4个年度春季调查中,1999年共采集到鱼卵439粒,仔稚鱼2 293尾,隶属7目12科13种;2001年共采集到鱼卵1 341粒,仔稚鱼7 467尾,隶属10目16科29种;2004年共采集到鱼卵52粒,仔稚鱼328尾,隶属7目10科16种;2007年春季共采集到鱼卵51粒,仔稚鱼638尾,隶属8目12科16种。
3.2 鱼类浮游生物丰度及其空间分布
1999年至2007年间,2001年长江口春季鱼类浮游生物丰度高于1999年,2004年迅速下降,仅为1999年的13.9%和2001年的4.3%;与2004年相比,2007年长江口鱼类生物丰度略有回升,分别为1999年的25.2%和2001年的7.8%。
如图2所示,1999年鱼类浮游生物在长江口南支、调查水域的南部和东北部均有高分布点出现。2001年主要集中分布在长江口口门内和长江口南部近岸水域。2004年和2007年,仅有零星分布点丰度超过250个/站,其中2004年主要分布在南部近岸水域,2007年增加了长江口南支河道水域。
3.3 优势种组成及空间分布
依据IRI指数确定的优势种及其组成(见表1)。1999年优势度较高的种类有、凤鲚、六丝矛尾虎鱼和白氏银汉鱼;2001年,白氏银汉鱼失去其优势地位,优势种组成中增加了小黄鱼、矛尾复虎鱼和松江鲈;2004年和2007年的优势种包括凤鲚、白氏银汉鱼、和松江鲈,但2个年度优势种的排序不同:2007年白氏银汉鱼取代凤鲚,成为优势度最高的种类。从表1可以看出:1999年至2007年,凤鲚和始终在长江口鱼类浮游生物中占据优势地位;白氏银汉鱼除在2001年出现过波动外,在其它年度也处于优势地位;松江鲈的优势度从1999年的34.33上升到2007年的226.45,在长江口鱼类浮游生物群落中的重要性逐步提升。
从1999年至2007年,长江口鱼类浮游生物优势种的丰度发生了显著变化。与2001年相比,2004年、凤鲚和松江鲈丰度分别减少了93.3%,98.2%和90.2%,在2007年仅略有回升。

图2 长江口鱼类浮游生物丰度空间分布(个/站)Fig.2 The spatial distribution of ichthyop lankton in the Changjiang River estuary

表1 春季鱼类浮游生物优势种类组成Table 1 The dom inant ichthyop lank ton com position in spring
3.4 群落结构
4个不同年份调查中鱼类浮游生物群落间相似性指数见表2。以最近的2007年为例,与2001年的相似性指数较低,与2004年的相似性指数最高,高达71.17%。2004年与2001年的相似性最低,为46.65%。可以看出,与2001年相比,蓄水后的长江口鱼类浮游生物群落格局出现了一定程度的变异。
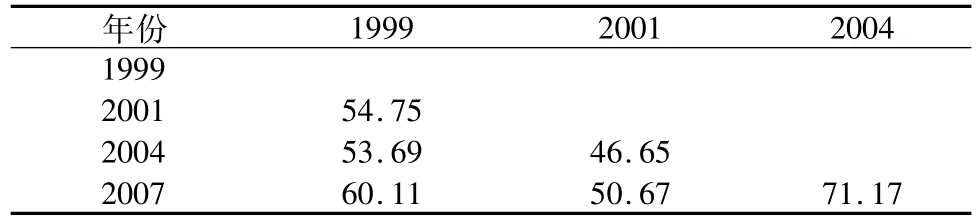
表2 不同年份间种类群落组成的相似性指数Table 2 The sim ilarity indexes of species composition between different years%
ANOSIM分析表明:4个年度长江口及其邻近海域鱼类浮游生物群落结构总体差异显著(R=0.112,P=0.2)。其中2004年和2007年的相似性程度最高(R=0.075,P=7.4),其次是1999年和2007年(R=0.083,P=8.2);蓄水后的2004年与1999年和2001年的长江口鱼类浮游生物群落结构差异显著(P<0.05),与2007年的差异不显著(P>0.05)。
利用SIMPER分析,可得到影响长江口鱼类浮游生物群落组成差异的物种贡献率。分析结果显示,?和凤鲚是对2004年与1999年、2001年间鱼类浮游生物群落结构差异贡献最大的种类,贡献率分别为11.93%和11.81%;同时,凤鲚对2007年与1999年、2001年间的群落结构差异贡献最大,分别为13.35%和11.36%。可以看出,长江口鱼类浮游生物优势种凤鲚和?丰度的时空变异造成了4次春季调查年度间群落结构变异。
3.5 群落多样性
在4个调查年份中出现的鱼卵和仔稚鱼的种类分别为13,29,16和16种,呈现出先升高,随后又下降的趋势。在研究区域内,种类丰富度指数也呈现类似的变化趋势。但均匀度和多样性指数则有不同的变化,均匀度指数除了2001年有一定的波动外,总体成上升的趋势,多样性指数则刚好相反,呈下降的趋势(见表3)。对4个调查年份中的3个生物多样性指数的ANOVA分析发现,丰富度和均匀度指数年度间差异不显著(P>0.05),而Shannon-Wie-ner指数则有极显著差异(P<0.01),其中1999年和2007年(P<0.05)、2001年和2004年(P<0.05)差异显著,2001年和2007年(P<0.01)差异极显著。可以看出,蓄水后的两个年度的生物多样性显著低于蓄水前水平。
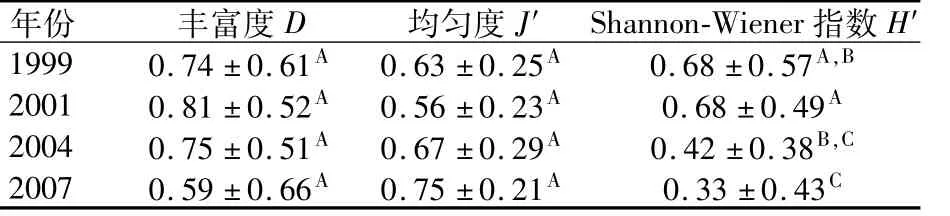
表3 长江口及其邻近海域鱼类浮游生物多样性指数变化Table 3 Changes of ichthyop lankton biodiversity in the Yangtze River estuary and adjacent coastal sea area
4 讨 论
4.1 种类组成和数量的变化
河口的环境特点和物质基础决定了栖息于河口的鱼类浮游生物的种类和数目。拥有众多的稀有种和高丰度的少数种是河口鱼类浮游生物群落普遍的特征[4],长江口也不例外。在4个调查年份中,共记录鱼类浮游生物39种,其中优势种集中在5~6种,其丰度占总丰度的90%以上。4个航次调查中,出现年度间种类演替:除、凤鲚、松江鲈、白氏银汉鱼、六丝矛尾虎鱼、小黄鱼和七星底灯鱼等种类外,部分种类在三峡水库蓄水前后出现或消失。例如和鲐在2004年和2007年没有捕获,银鲳的捕获量也明显下降;细条天竺鱼以前没有捕获,在2004年和2007年则有出现。另外,还有部分物种只在个别年份有捕获。例如,1999年出现的鲷科一种,2001年的褐菖、焦氏舌鳎和短吻三线舌鳎,2004年的寡鳞飘鱼、棘头梅童鱼,2007年的日本鬷、有明银鱼等。旧物种的消失和新物种的出现皆是因为其原生境条件发生改变。河口地区受干流水利工程和捕捞、污染等人为因素的影响使环境条件波动较大,造成了一部分种类消失,一部分种类新出现的现象,这种现象也曾在Ria de Aveiro河口的年际变化中出现[5]。
从种类盐度适应类型上看,半咸水型和沿岸型鱼类浮游生物始终是长江口及其邻近海域的主要种类;近海型的和七星底灯鱼也一直在长江口及其邻近海域的鱼类浮游生物群落中占有重要的地位,其它近海型种类多为稀有种且年际波动大。淡水型种类只分布于低盐度水域,受冲淡水的影响较大,调查区域内淡水型出现不多。2004年出现了银飘鱼和寡鳞飘鱼2种较之以前有增加的趋势,可能与1-5月三峡水库因需腾出防洪库容而放水,下泄流量较天然流量有所增加,长江口冲淡水范围增加有关。
从鱼类浮游生物丰度变化可以看出,在三峡水库蓄水前后鱼卵和仔稚鱼在绝对数量上有明显的差别,蓄水后已成明显的下降之势。这其中有三峡水库蓄水影响径流分布改变长江口及其邻近海域的理化和生物因子的原因,也有近年来持续高强度的捕捞和河口水域的污染影响。另外,2004年春季数量的严重减少还与生物因素相关。2004年春季大型水母爆发,其渔获量占到了98.44%[6],水母不仅是鱼类浮游生物的捕食者,而且可以分泌大量的毒素,直接引起鱼类浮游生物的死亡,是影响长江口及其邻近海域鱼类浮游生物的数量下降及鱼卵种类的单一和数量极少的一个重要因素。
从空间分布上,长江口及其邻近海域的鱼类浮游生物分布更集中于长江口附近,调查海域其它站位的数量在蓄水后有相应减少。除了生殖习性等生物因素外,盐度、温度、混浊度、溶解氧、径流量、栖息地结构属性、深度、水动力条件以及到河口的距离等非生物因素决定了河口鱼类的分布区域[7],长江口鱼类浮游生物部分优势种分布区域的变动是长江口及其邻近海域生态系统结构变化的显著信号。Drinkwater和Frank[8]曾提出河口径流的变动引起地形的改变、温度和盐度变动和浊度和溶解氧的变化,这些效应最终都影响到河口生物的丰度和分布类型。至于哪些环境条件的改变导致了长江口鱼类浮游生物数量变动,还需深入的研究论证。
4.2 优势种的变化
4.3 群落结构的相似性和生物多样性的变化
鱼类浮游生物是鱼类的补充资源,鱼类浮游生物群落结构的稳定性对鱼类资源的补充,延续和维持生态平衡具有重要的意义。通过Bray-Curtis相似性指数的比较发现,长江口鱼类浮游生物群落结构在4个调查年份中总体差异显著。由SIMPER分析的结果可知,长江口及其邻近海域鱼类浮游生物群落的年际差异主要源于部分优势种丰度变化。
群落中物种多样性指数是群落中种的多少和种间个体数量的均衡性的反映。对长江口及其邻近海域的各个调查站位3项指数研究表明,物种丰富度和均匀度指数在4个调查年份并无明显的差异,而Shannon-Wiener指数则存在较大的差异,并呈下降的趋势,这种不均衡性是由优势种丰度上的减少和出现频率变小,且数量少的稀有种相对减少引起的。Connell[10]提出中度干扰理论,即中等程度的干扰能维持高的多样性,而过度的干扰会降低多样性水平。Sanders[11]也指出低物种多样性与增加的扰动和不断变化的非生物环境条件有关。从上世纪70年代后期开始,长江口及其邻近海域经受了滩涂围垦、水域污染和过度捕捞等诸多因素的干扰,2003年的三峡工程建设也带来了长江口一系列环境条件的直接和间接变化,环境干扰的最终结果带来了长江口鱼类浮游生物多样性的降低。
[1] 罗秉征,沈焕庭.三峡水库与河口生态环境[M].北京:科学出版社,1994.(LUO Bing-zheng,SHEN Huan-ting.The Three Gorges Project and the Ecological Environment of Yangtze Estuary[M].Beijing:Sciecnce Press,1994.(in Chinense))
[2] SEDBERRY R G,CARTER J.The fish community of a shallow tropical lagoon in Belize,Central America[J].Estuaries,1993,6:198-215.
[3] BRAY JR,CURTIS JT.An ordination of the upland for-est communities of southern Wisconsin[J].Ecological Monographs,1957,27:325-349.
[4] WHITfIELD A K.Ichthyofaunal assemblages in estuar-ies:a South African case study[J].Fish Biology and Fisheries,1999,9:151-186.
[5] BAUCHOTM L,PRASA.Guia de Los Peces de Mar de Espafia y de Europa[M].Bracelona:Ediciones Omega,1987.
[6] XIANW W,KANG B,LIU R Y.Jellyfish blooms in the Yangtze estuary[J].Science,2005,307:41.
[7] AKIN S,B E,WINEMILLER K O,YIMAZ H.Fish as-semblage structure of Koycegiz Lagoon-Estuary,Turkey:Spatial and temporal distribution patterns in relation to environmental variation[J].Estuarine Coastal and Shelf Science,2005,64:671-684.
[8] DRINKWATER K F,FRANK K T.Effects of river regu-lation and diversion onmarine fish and invertebrates[J].Aquatic Conservation:Freshwater and Marine.Ecosys-tems,1994,4:135-151.
[9] STRYDOM N A,WHITFIELD A K,WOOLDRIDGE T H.The role of estuarine type in characterizing early stage?sh assemblages in warm temperate estuaries[J].South Africa.African Zoology,2003,(38):29-43.
[10]CONNELL JH.Diversity in tropical rainforests and coral reefs[J].Science,1978,199:1302-1310.
[11]SANDERSH L.Marine benthic diversity:a comparative study[J].American Naturalist,1968,102: 243-282.
(编辑:罗玉兰)
Ichthyoplankton Community Structure Characteristics during Spring in Yangtze River Estuary before and after Im poundment of Three Gorges Reservoir
LIU Shu-de1,2,XIANWei-wei1
(1.Key Laboratory of Marine Ecology and Environment Sciences,Institute of Oceanology,Chinese Academy of Sci-ence,Qingdao 266071,China;2.Graduate University of the Chinese Academy of Sciences,Beijing 100049,China)
Spatial and temporal variation of ichthyoplankton community structure before and after impoundment of Three Gorges Reservoir(TGR)in the Yangtze River estuary and adjacent sea area(30°45′-32°00′N,121°00′-123°20′E)was analyzed based on the samples at40 stations during spring of1999,2001,2004 and 2007.Through the study,39 species,consisting of freshwater,brackish,coastal and marine varieties,were collected in trawl nets.The ichthyoplankton abundance degree in 2004 rapidly was decreased with 13.9%of 1999 and 4.3%of 2001,respectively,and appreciably increased in 2007.The dominant species comprised Engraulis japonicus,Coil-iamystus,Allanetta bleekeri,Trachidermus fasciatu and Chaeturichthys hexanema.The dominance of Engraulis ja-ponicus and Coiliamystuswas reduced in 2004.Therewas significant dissimilarity within four year’s ichthyoplank-ton community structure,and the dissimilarity in 2004 was higher,comparied with 1999 and 2001.Similarity of percentage analysis showed that Engraulis japonicus and Coilia mystus represented the highest contribution in the community structure variation.Moreover,the similiarity index of ichthyoplankton was highest between 2004 and 2007 with the value 71.17%;Ichthyoplankton community diversity before and after impoundment of TGR was sig-nificantly different in the Yangtze Rive estuary,and the diversity after impoundment evidently decreased.A series of environmental disturbances,which happened in the Yangtze River estuary and adjacent water during 1999-2007,directly or indirectly caused the variation of ichthyoplankton community structure.
Yangtze River estuary;ichthyoplankton;community structure; diversity
Q915.862
A
1001-5485(2010)10-0082-06
2010-08-23
中国科学院知识创新工程重大项目(KZCX1-YW-08-01)
刘淑德(1985-),男,山东烟台人,硕士研究生,主要从事鱼类资源与生态学研究,(电话)0532-82898594(电子信箱)shudliu@126.com。
