钙对镉胁迫下生菜幼苗生长和生理的影响
2010-07-31石贵玉李佳枚廖文雪
石贵玉,李佳枚,韦 颖,廖文雪,叶 静
(1.广西师范大学 生命科学学院,广西 桂林 541004;2.广西环境工程与保护评价重点实验室,广西 桂林 541004)
随着矿产资源大量开发利用,各种化学产品、农药、化肥的广泛使用以及城市污泥、污水的农业利用,重金属对土壤、水体的污染也越来越严重[1-2]。在主要的重金属污染物中,镉 (Cd) 有比较高的生物毒性,Cd对高等植物的毒害作用主要表现为抑制植物对水分的吸收和运输;抑制光合作用和呼吸作用;抑制其它新陈代谢等[3]。
很多研究表明,钙 (Ca)在植物抗逆境过程中具有重要作用,外源Ca可增强植物对许多非生物逆境的适应性,减轻逆境对植物所造成的伤害[4-5],营养液及土壤中添加Ca对Cd的毒害具有缓解作用[6]。
生菜 (Lactuca sativa)别名莴苣,属菊科莴苣属植物,生菜营养价值较高,主要以生食为主,是凉拌色拉、汉堡包不可缺少的蔬菜。在南方普遍栽培,是人们喜食的叶菜类之一。因生菜多在城市郊区种植,易受Cd污染。因此,研究生菜Cd积累及毒害生理机制,培育食用部位含Cd量低的生菜品种具有重要的理论和实际意义。
本试验以生菜为材料,研究Ca对不同 Cd水平胁迫下的生菜植株生长生理的影响,探讨Ca对不同Cd水平下生菜抗氧化酶活性的变化以及影响植株生长的生理机制,以求为发展绿色食品和无公害蔬菜提供科学依据。
1 材料与方法
1.1 材料和处理
试验用生菜秧苗受赠于桂林市穿山乡,品种为生菜王 (Lettuce)。
秧苗先用自来水冲洗根上的泥土,挑选生长一致的幼苗在营养液中进行预培养,每盆5株,营养液配方采用华南农业大学叶菜类配方[7],预培养1周后,进行胁迫处理。
试验设置如下:1号不加 Cd对照 (CK);2号Ca 20 mg·L-1(以CaCl2加入,下同);3号Cd 10 mg·L-1(以 CdSO4加入,下同);4号 Cd 20 mg·L-1;5 号 Ca 20 mg·L-1+Cd 10 mg·L-1;6号 Ca 20 mg·L-1+Cd 20 mg·L-1。重复 3 次。分别于处理后10 d取样测定过氧化物酶 (POD)、过氧化氢酶 (CAT)、超氧化物歧化酶 (SOD)的活性和丙二醛 (MDA)、还原性糖含量。第20天测定叶绿素含量和植株生长量。
1.2 测定方法
POD活性的测定:用愈伤木酚氧化法[8],以470 nm波长下每1 min每1 g材料的光密度变化表示酶活性大小。
CAT活性的测定:用紫外吸收法测定[9],以1 min内D240减少0.1为一个酶活性单位。
SOD活性的测定:按Giannopolitis和Ries的方法,以每单位时间内抑制光化还原50%的氮蓝四唑 (NBT) 为一个酶活性单位[10]。
MDA含量的测定:采用硫代巴比妥酸比色法测定 MDA 含量[8]。
还原性糖含量的测定:用蒽酮法[8]。
叶绿素含量的测定:采用分光光度法,用Arnon 法计算叶绿素含量[8]。
生长量测定:采用尺寸测量法和天平法。
数据采用Excel软件处理,采用 SPSS 13.0软件用单因素方差分析对以上数据进行样品的差异显著性检验,采用LSD方法进行多重比较检验。
2 结果与分析
2.1 对生菜幼苗生长的影响
从表1可见,培养20 d时,2号处理株高和鲜重与对照基本相同,3号、4号处理则株高和鲜重均比对照低,且浓度愈高作用愈明显,但5号和6号有Ca存在时,株高和鲜重则分别比3、4号高,说明Ca有缓解Cd毒害的作用。
2.2 对生菜幼苗叶绿素含量和组分的影响
叶绿素是植物进行光合作用的色素,其含量高低可以在一定程度上反映光合作用水平[11]。叶绿素含量降低,光合作用减弱,会导致植物生长受抑制,生物量下降。从表1可以看出,随着Cd污染浓度的升高,叶绿素总量呈下降的趋势,同时叶绿素a/b比值随着Cd污染浓度的增高而降低,说明Cd毒害使生菜叶片叶绿素含量降低,且对叶绿素a的影响小于对叶绿素b的影响。从表1中还可以看出,Ca+Cd,处理幼苗叶绿素含量和组成均高于Cd处理,说明Ca有缓解Cd毒害,保持较高水平叶绿素含量的作用。
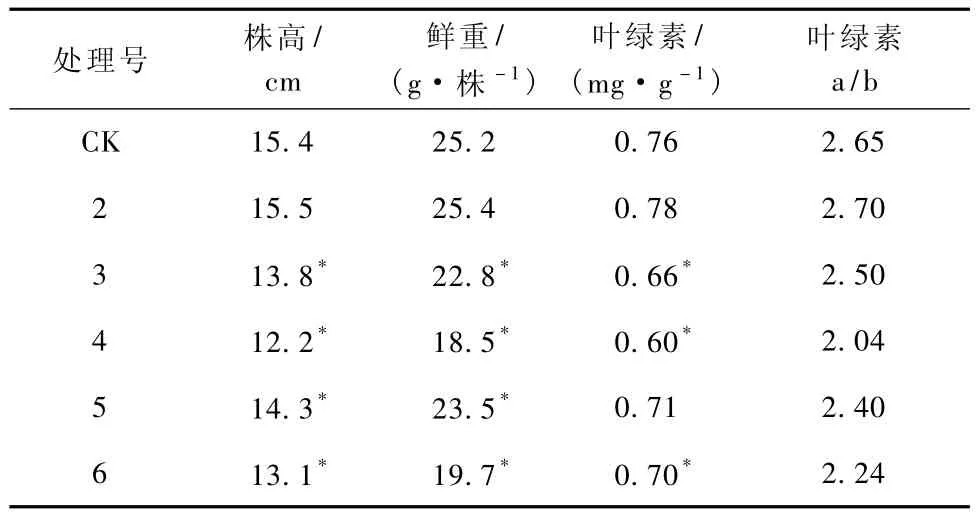
表1 各处理对生菜幼苗株高、鲜重和叶绿素的影响
2.3 对生菜幼苗抗氧化能力的影响
2.3.1 SOD活性
SOD是重要的活性氧清除酶,能保护细胞免受活性氧胁迫的伤害。图1看出,Cd胁迫下SOD活性有所提高,Ca存在时SOD活性比单一Cd胁迫的酶活性高,这可能与植物在逆境时通过提高SOD活性起应激反应有关。说明Ca有提高生菜幼苗抗重金属胁迫的能力。但随着胁迫时间的延长,各处理的SOD活性均大幅度下降。
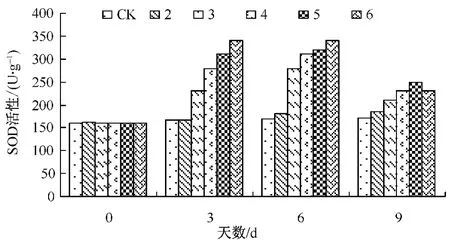
图1 Ca对Cd胁迫下生菜幼苗SOD活性的影响
2.3.2 CAT活性
CAT也是清除H2O2重要的保护酶,图2反映了Cd胁迫和Ca对生菜幼苗叶片CAT活性的影响。在处理的早期处理3~6生菜幼苗中的CAT活性均有明显的增加,Ca处理从开始到结束,CAT活性均比CK组高,但是差异并不明显。这说明Ca有提高CAT活性的能力,在处理后期,处理3~6幼苗中的CAT活性均下降。试验结果与生菜SOD活性变化相似,这可能与植物在逆境时也通过提高CAT活性消除活性氧有关。
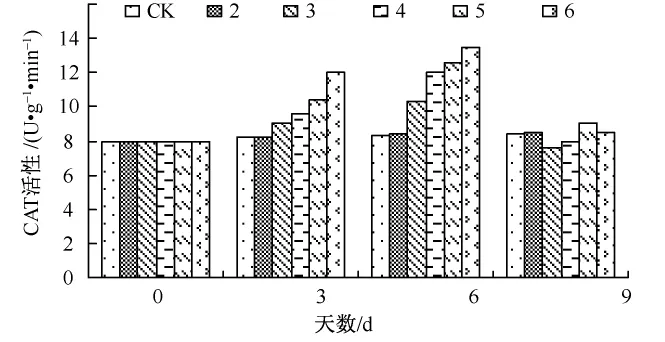
图2 Ca对Cd胁迫下生菜幼苗CAT活性的影响
2.3.3 POD活性
POD是植物体内重要的抗氧化酶,能有效清除H2O2。图3反映了镉Cd胁迫时Ca对生菜幼苗叶片POD活性的影响。
从图3可知,随着处理时间的延长,处理3~6的POD活性从开始到实验结束始终高于对照,处理5~6的POD活性一直比处理3~4的 POD活性高,反映Ca有提高POD活性的作用。
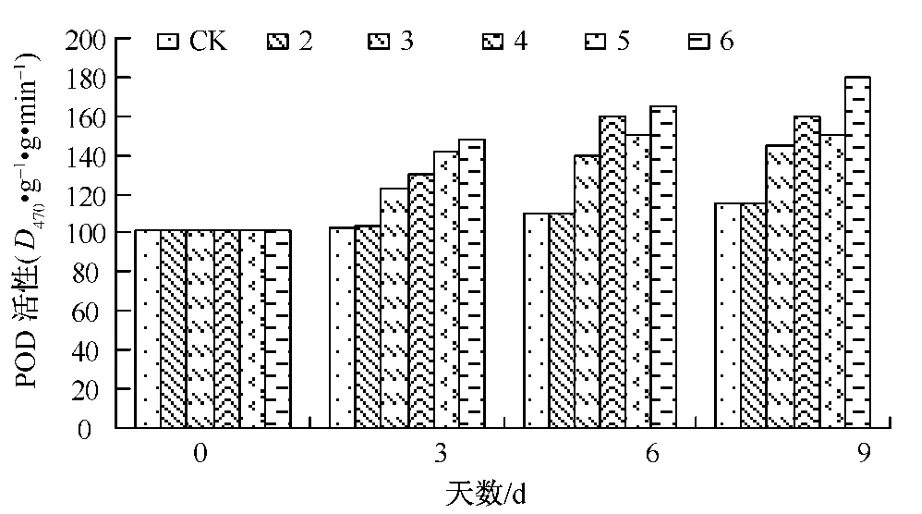
图3 Ca对Cd胁迫下生菜幼苗POD活性的影响
2.3.4 MDA含量
植物在逆境环境中,细胞膜中的不饱和脂肪酸会发生过氧化作用产生MDA,使质膜系统受到伤害,其选择性受到破坏,导致细胞内电解质外渗量增加,因而MDA含量可反映膜脂过氧化作用的强弱,质膜透性可表示膜伤害或变性的程度[12]。图4反映了镉胁迫下Ca对生菜幼苗叶片MDA含量变化的影响。在整个试验中,处理3~6植株叶片MDA含量明显比CK组多,达到了显著水平 (P<0.05),特别是处理5~6始终比处理3~4的MDA含量要低,并且达到了显著水平,这说明Ca离子在一定程度上增强了生菜抵抗重金属的能力。
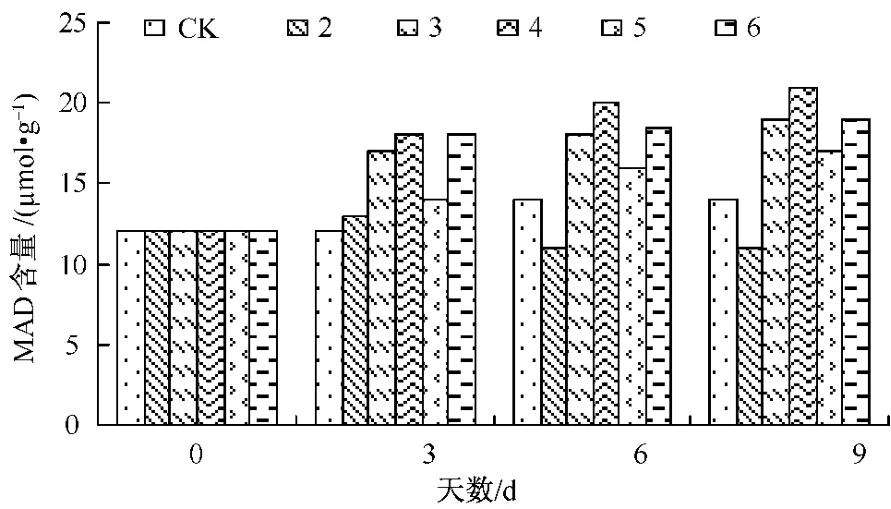
图4 Ca对Cd胁迫下生菜幼苗MDA含量的影响
2.4 对生菜幼苗可溶性糖含量的影响
可溶性糖是参与细胞渗透调节的主要物质之一,糖的积累能提高细胞渗透浓度,在植物抗性的维持中具有重要作用。图5反映了Ca对Cd胁迫下生菜幼苗可溶性糖含量的影响。从图5中可知,Cd胁迫使生菜幼苗体内的可溶性糖含量下降,加Ca处理组生菜幼苗中的可溶性糖含量均比未加Ca处理组幼苗中可溶性糖含量高,差异明显已达到显著水平,反映Ca处理有缓解镉毒害的作用。
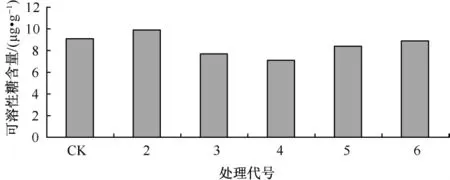
图5 Ca对Cd胁迫下生菜幼苗可溶性糖含量的影响
3 小结和讨论
活性氧 (如·O2-,·OH,H2O2等)是植物正常代谢中的产物,在逆境胁迫下,活性氧代谢系统的平衡会受到影响,植物体内的活性氧大量生成。活性氧的升高能启动膜脂过氧化或膜脂脱脂作用,从而破坏膜结构[8-9]。SOD和 CAT是存在于植物细胞中重要的清除自由基酶类之一,它的主要功能是清除超氧化物自由基·O2-和H2O[213],其活性的大小与植物抗逆的能力相关[14]。试验反映,在Cd胁迫的早期,加入Ca能提高SOD和CAT的活性,从而提高清除自由基的能力,缓解Cd胁迫对生菜幼苗的伤害;但是,随着Cd胁迫时间的延长,Ca逐渐失去提高幼苗SOD、CAT活性的作用,这可能是加入Ca使SOD、CAT活性提高是有限度的,随着胁迫的加重,保护机制可能被破坏,因此,随着Cd胁迫时间的延长,SOD、CAT活性又发生了急剧下降。在本次试验中 CAT、SOD两者活性的影响具有相关性,两者活性变化趋势基本一致,这说明加Ca能够通过提高抗氧化酶类活性从而提高对镉的抗逆性。
POD是植物体内重要的代谢酶,也是植物体内的抗氧化酶之一,过氧化物酶活性的高低不仅反映植物生长发育及内在代谢情况,同时也是植物抗性好坏的标志之一[12,15]。从试验可以看出 Cd胁迫时Ca对生菜幼苗叶片POD活性的变化与Cd胁迫时Ca对生菜幼苗叶片SOD、CAT活性的变化不一致,POD的活性是从试验开始到试验结束持续升高,表明其对Cd胁迫的适应性比较强,暗示POD活性在抗Cd胁迫的过程中起关键性的作用。
MDA是生物体代谢和生物膜系统脂质过氧化作用下的产物,MDA含量的多少反映了细胞膜脂过氧化的程度,是一种重要的逆境生理指标。本试验说明生菜体内膜脂过氧化的程度受Cd处理浓度和处理时间的双重影响,处理浓度越大、时间越长,生菜体内膜脂过氧化程度越大,植株受的伤害越大。Ca能维持细胞膜结构的稳定,在逆境下具有保护植物细胞膜免遭损伤、降低离子渗漏的功能,原因可能是Ca与膜磷脂的极性头部结合,通过交联作用,使膜脂分子和蛋白质分子结合紧密,从而降低了膜的透性[15]。说明Ca减轻了镉的生理毒害,可能与它维持细胞膜和细胞壁结构的完整性有关。
植物为了适应逆境条件,也会主动积累一些可溶性糖、脯氨酸等物质,以适应外界环境条件的变化。可溶性糖是参与细胞渗透调节的主要物质之一,糖的积累能提高细胞渗透浓度,增加抗性能力,在植物抗性的维持中具有重要作用。贺迪等[16]发现Zn2+胁迫下可溶性糖含量的增加有利于细胞或组织持水,防止脱水从而使得细胞内大分子糖类的分解加强而合成受抑,最终导致可溶性糖含量上升。本试验也表明,外源Ca可以促进生菜体内可溶性糖含量增加,这可能与提高生菜抗重金属镉毒害的能力有关。
据报道,Cd胁迫下,植物叶绿体结构受损,叶绿素生物合成受阻[16],导致叶绿素总量降低,植物光合作用受到抑制,从而引起植物长势较弱,生长量减少。本试验中,随着Cd浓度的增加和时间的延长,生菜叶绿素含量明显降低、植株矮小,这与王林在萝卜上的研究结果相似[17],导致了植物的光合能力降低,生长延缓。Ca处理叶绿素含量和组成均高于 Cd污染处理,这说明了Ca能增加生菜幼苗的抗Cd能力。
[1]罗春玲,沈振国.植物对重金属的吸收和分布 [J].植物学通报,2003,20(1):59-66.
[2]黄铭洪.环境污染与生态恢复 [M].北京:科学出版社,2003.
[3]张军,束文圣.植物对重金属镉的耐受机制 [J].植物生理学与分子生物学报,2006,32(1):1-8.
[4]李美茹,刘鸿先,王以柔,等.钙对水稻幼苗抗冷性的影响 [J].植物生理学报,1996,22(4):379-384.
[5]周青,黄晓华,王东燕,等.钙对酸雨伤害甜瓜幼苗的影响 [J].植物生态学报,1999,13(2):186-191.
[6]周卫,汪洪,林葆.镉胁迫下钙对镉在玉米细胞中分布及对叶绿体结构与酶活性的影响 [J].植物营养与肥料学报,1999,5(4):335-340.
[7]刘士哲.现代实用无土栽培技术 [M].北京:中国农业出版社,2001:124.
[8]张志良.植物生理学实验指导 [M].北京:高等教育出版社,2000:160-161.
[9]邹琦.植物生理学实验指导 [M].北京:中国农业出版社,2001:163-171.
[10]GiannopolitisC N,Ries S K. Superoxide dismutation Ⅰ.Occurrence in higher plants [J].Plant Physiol,1997,53:315.
[11]秦天才,阮捷,王腊娇.镉对植物光合作用的影响 [J].环境科学与技术,2000(1):33-44.
[12]孙文越,王辉.外来甜菜碱对干旱胁迫下小麦幼苗膜脂过氧化作用的影响 [J].西北植物学报,2001,21(3):487-491.
[13]孙赛初,王焕校,李启任.水生维管束植物受Cd污染后生理生化变化及受害机制初探 [J].植物生理学报,1985,11(2):147-153.
[14]石贵玉,康浩,段文芳.重金属镉对红树植物白骨壤和桐花树幼苗生理特性的影响 [J].广西植物,2009,29(5):644-647.
[15]李美茹,刘鸿先,王以柔,等.钙对水稻幼苗抗冷性的影响 [J].植物生理学报,1996,22(4):379-384.
[16]贺迪,刘元国,黄玉娥,等.钙对不同浓度镉胁迫下芦苇幼苗叶绿素及抗氧化酶系统的影响 [J].农业环境科学学报,2007,26(1):169-201.
[17]王林,史衍玺.镉、铅及其复合污染对萝卜生理生化特性的影响 [J].中国生态农业学报,2008,16(2):411-414.
