林木中的电阻抗参数与温度的关系
2010-06-21马凤翔陈晓阳
马凤翔,陈晓阳
(1.北京林业大学理学院,北京 100083;2.北京林业大学生物科学与技术学院,北京 100083;3.华南农业大学 林学院,广州 510642)
1 引言
电阻抗技术是一个边缘性的研究领域,是研究物质电学性质的一种方法。经过几十年的发展,电阻抗技术已经在材料研究、固体表面处理、器件研究、电化学领域、地球科学和生命科学等多个领域得到关注和青睐。国外生物电阻抗技术的研究水平较高,主要集中在生物电阻抗测量技术及测量装置的研究和模型构建及参数提取的研究。国内学者在医学工程方面的研究较为深入,涉及建模,算法、图像处理,临床应用等多方位多层次,在农业、林业、园艺方面的研究起步较晚且多停留在应用方面。
2 电阻抗法在林木抗寒研究中的应用
抗寒性是指植物不受冻害所能忍受的冰冻温度。电阻抗测量技术用于植物是将林木的茎干、针叶或样本作为一个电容器[1],通过测定其电阻抗及其相关的电特性参数,建立林木茎干、针叶或样本的电阻抗等参数与林木的抗寒性和受冻害程度的关系,来评价林木质量。
目前该法主要是用于测定林木抗寒性。植物在寒害和冻害锻炼期间以及在遭受冻害时植物组织细胞壁首先受到伤害,细胞膜发生透性改变,离子泄露,膜电阻随之发生改变,从而使组织电阻发生改变。这样,通过电阻抗法可以检测到冷冻胁迫对组织特性的影响[2]。Repo对欧洲赤松(Pinussy lvestris)幼树当年生枝条用电阻抗图谱(EIS)法和电导(EL)法测定为期一年的抗寒性试验[3],王爱芳以8年生樟子松(Pinus sy lvestris L.var.mongolica Litv.)苗为试材,在抗寒锻炼阶段(10月份)和脱锻炼阶段(3月份)分别取样进行电阻抗图谱(EIS)法和电导(EL)法对樟子松茎和针叶进行了抗寒性测定[4]。他们的研究结果均显示,EL法确定的针叶抗寒性与利用阻抗法中的胞外电阻率参数re求出的茎抗寒性之间存在很强的相关性(相关系数=0.97)。Mancuso对4种Callistemon属和2种G revillea属植物进行的抗寒性研究[5],得出非常类似结果。H urme、Repo等用电阻抗图参数re法求出的欧洲赤松茎抗寒性低于EL法求出的针叶抗寒性[6,7]。Repo、Ryyppö等的研究显示当样本抗寒性很强时,EIS法可能比EL法低估抗寒性[8,9]。但王爱芳的结果表明,EIS胞外电阻率re法比EL法求出的抗寒性高[4]。不同的结果可能的原因还不是清楚,有待于进一步的研究。另外,Repo、王爱芳的研究表明在抗寒性快速增强的阶段,不经冷冻处理的样本用阻抗谱中的驰豫时间估测抗寒性是一种快速有效的测定方法,具有潜在的实用性,是一种很有前途的方法[7]。同样,刘辉对金叶女贞(Ligustrum vicaryi)茎抗寒性和电阻抗图谱参数的影响分析指出电阻抗图谱法是一种测定植物抗寒性,尤其是估测不经冷冻处理样本抗寒性的可行的方法[10]。就现有的研究文献来看电阻抗技术主要是偏向于对针叶树种抗寒性的研究,且较为可靠,对其他逆境的研究少见报道,刘晓红等曾以不同生育期小麦叶片为材料进行水分胁迫实验,得出的结论是,电阻抗图谱参数尚不能用于表征小麦品种的抗旱性[11]。
3 尚未利用到的参数
3.1 极化率
利用阻抗法研究林木或植物的抗寒性时,把待测的样品作为电容器,也就是将研究的样品作为了电介质材料。根据电介质的极化机理,电介质的极化可以归结为5种形式:电子位移极化、离子位移极化、转向极化、热离子极化和空间电荷极化[12]。由于电子的分布与温度无关,电子位移极化与温度基本无关。离子位移极化只可能在离子晶体中建立,离子极化只是与离子结构的参数有关,与温度无关。空间电荷极化,指电介质在电压作用下有能量的损耗,在讨论温度与损耗角正切、介电常数时再作阐述。
作为研究对象的林木是一种比较复杂的电介质,一方面,作为极性电介质林木中一定含有偶极分子或偶极基团如HCl、NH3等,在无外电场作用时,具有一定的偶极矩,但是由于分子不规则的热运动,分子在各个方向的分布几率是相等的。因此,就整体来看,宏观偶极矩为零。当偶极分子受外电场作用时,偶极分子将受到电场力矩的作用而趋于转向电场方向,于是就介质整体而言,出现沿电场方向的宏观偶极矩。由于分子的热运动及分子间的相互作用,并非所有的分子都能整齐地沿电场方向排列。一般电介质的转向极化由于受到分子热运动的无序化作用和外电场的有序化作用以及分子间的相互作用,它的建立需要较长的时间,约为10-6~10-2s数量级或更长。在一般情况下,电介质的转向极化率σT与绝对温度T成反比[12]。另一方面,作为电介质的树木其中一定存在着一些N+a、K+等弱联系的离子,它们处于介质中能阶较低的平衡位置,并在平衡位置附近作热振动。在一定的温度下,离子的热振动能超过邻近分子对它的束缚势垒时,离子就能越过势垒而到达新的平衡位置,在没有外电场时,离子的随机分布没有改变,离子没有定向的迁移,不呈现宏观的偶极矩,但在外电场的作用下,外电场破坏了电介质中弱联系离子的随机均匀分布,便形成了宏观的偶极矩,从开始加上外电场到离子极化达到稳态时,同样需要一定的时间,一般电介质需要的时间约为10-10~10-2s数量级,其热离子极化率aT也与绝对温度 成反比[12],作为我们的研究对象林木被极化时,其转向极化和热离子极化的极化率是否与绝对温度成反比或者有何关系,这需要实验验证。
3.2 相对介电常数
对于植物组织,在外电场的作用下,是有能量损耗的,一种是由于电导(植物组织中含有水分,电解质和各种矿物质,具有导电性)引起欧姆损耗,主要由电学指标电阻率确定,另一种是由于极化引起的损耗。在恒定电场作用下,组织中没有周期性的极化过程,因此损耗仅由电导引起,这时组织的品质可用体积电阻率来描述。而在交变电场作用下,除了电导引起的损耗外,还有周期性的与热运动有关的松弛极化,也引起组织的能量损耗,即极化损耗。当植物组织被外加的电场——不同频率下的交流信号作用时,组织发生极化,极化了的组织能够产生束缚电荷,束缚电荷产生附加电场,组织内部的电场就是外加的电场与组织内部因被极化而产生的附加电场的叠加。当外加的电场撤消后,束缚电荷产生的附加电场不会马上消退而发生弛豫;导致组织内部的总场振幅和相位、阻抗(包括实部与虚部、阻抗模值、阻抗相位角)、品质因数、介电常数、损耗角正切等电学参数均会发生变化。现有的文献中普遍应用的方法是电学指标电阻率rx,也就是电导率的倒数,参数rx主要反映的是介质在电场作用下的欧姆损耗,实质上能量损耗包括电导即欧姆损耗和极化损耗。对于研究极化损耗,关键的电学参数应该是介电常数和损耗角正切。所以,当植物组织在交变电场作用下,单考虑电阻率是不够全面的,我们应该将与极化损耗相关的参数介电常数和损耗角正切同时考虑进去。
把待测的对象看作一个由生物组织为电介质的电容器时,由于电容的大小是导体系统的物理尺度和周围电介质的特性参数决定,比如对于最简单的平行板电容器来讲,其电容大小与两板之间的距离成反比,与极板面积大小成正比,与板间填充的介电常数成正比。当所取的样品确定后,距离和面积参数确定,所以反映物质特性的介电常数是反映电容大小的一个关键的电学参数。在电场作用下,建立极化通常需要经过一定的时间才达到稳定状态,这对于恒定电场中的电介质来说是不成问题的,总有足够的时间让极化建立完全而到其稳定状态,相应恒定电场中的介电常数——静态介电常数。然而在交变电场中电介质的极化情况就不同了,极化将随着电场的变化而变化,如果电场随时间的变化很快,也就是电场的频率较大时,可以与极化建立的时间相比拟,极化就可能跟不上电场的变化了。这样,介质在交变电场下的动态介电常数与静态介电常数是不同的,并引起介质的极化损耗。这时表征介质电极化特性的常数用复数,″表示,ε′相当于通常的介电常数 εε″=ε′,由 ε′和 ε″组成的复介电常数和介质温度和电场频率有关[13]。
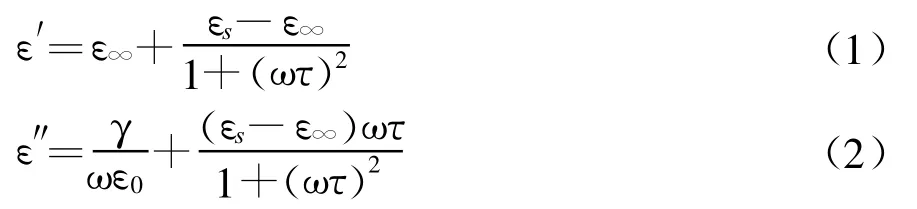
式中τ为松弛极化时间;εs为介质的静态介电常数,即介质在直流电场作用下的介电常数;ε∞为相应于电子位移极化的介电常数,等于光折射率的平方(ε∞=n2),称为光频介电常数。γ为电介质的电导率即电阻率的倒数;在高频下可以忽略。当温度升高时(T2>T1),ε′和ε″同时向高频方向移动,而εs和εmax值稍有下降。ε′随温度变化的曲线如图1所示[13]。
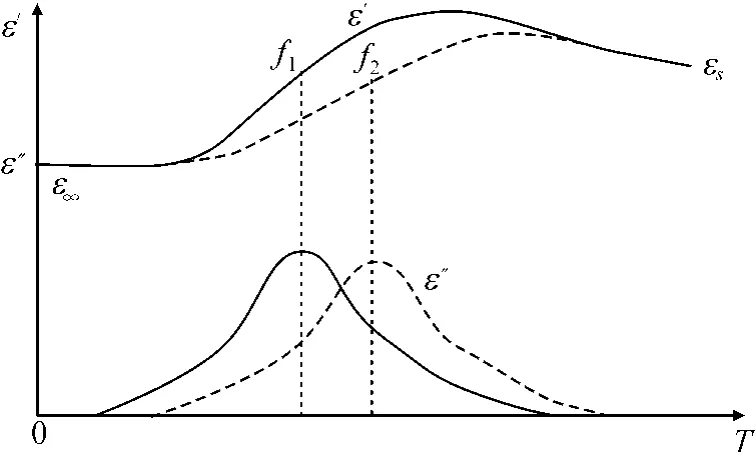
图1 ε′和 ε″的温度特性曲线
低温区反映ε∞随温度的变化,高温区反映εs随温度的变化,而在中间温度范围的ε′随温度上升而增大。从ε″的表达式可见,电阻率rx的倒数电导率γ只是ε″的组成部分,显然用rx不及ε″更能够准确地反映植物的特性。
3.3 损耗角正切
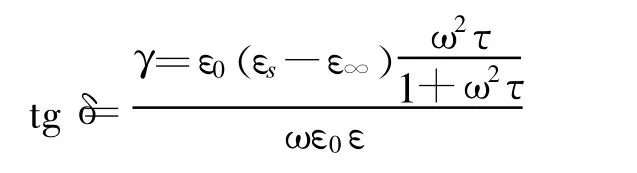
ε0为真空中的介电常数。从tgδ的表达式可见,电阻率rx的倒数电导率γ只是tgδ的组成部分,显然用rx不及tgδ更能够准确地反映植物的特性。
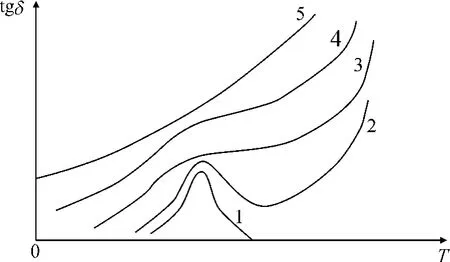
图2 电导率不同的介质的tgδ和T的关系
3.4 松弛极化时间
热离子极化和转向极化的粒子极化率均与温度成反比,建立热离子极化和转向极化又需要较长的时间——松弛时间。松弛时间τ与绝对温度呈指数关系,τ=(eu/kT)/(2v)[12],其中u是离子的活化能,k玻耳兹曼常数,v离子本征振动频率,对τ的名称,不同的文献称谓不同,有称弛豫时间的[14]。由表达式(1)和(2)同样可以看出,无论是复介电常数的实部ε′,还是虚部 ε″它们都是极化松弛时间 τ的函数,所以在利用阻抗法研究林木或植物与温度的关系时,τ是一个重要的参数,不可缺失。
如果在均匀介质中,弛豫时间τ为介电常数与电导率的比值[15],所以它能够比电阻率(电阻率与电导率互为倒数)更可靠地反映客观状态。近几年来对几个树种不经冷冻处理的样本研究中所发现的弛豫时间τ与抗寒性的相关最高。为研究抗寒性最重要、最精确的参数[2]。Repo用τ求出的抗寒性与常规方法求出的抗寒性的差异不超过±2℃范围[8]。刘辉通过对抗寒锻炼初期,短日照处理的金叶女贞茎的抗寒性分析,得出样本不经冷冻处理的情况下,可通过茎的弛豫时间τ估测抗寒性[10]。王爱芳等采用电阻抗图谱法和电导率法对不同发育时期的樟子松茎和针叶进行抗寒性测定,得出针叶的细胞膜时间量τ和茎的τ可以作为计算抗寒性的参数,不经冷冻处理的样本,用茎的τ和针叶的τ估计樟子松不同发育阶段抗寒性是很有前途的方法[4]。
4 结语
(1)由前面的分析得出弛豫(松弛)时间、极化率均与温度直接有关,介电常数、损耗角正切间接与温度有关,作为由于高温引起的植物伤害的现象——热害,从电介质物理学的角度可以得出,是可以利用阻抗谱参数研究植物的耐热性,就目前的研究文献看,阻抗谱参数只是局限在研究植物的抗寒性方面。在耐热性方面未见报道。事实上在冰冻、低温、高温、干旱、盐渍、土壤过湿和病害等各种逆境发生时,植物体内的水分状况有相似的变化,吸水力降低[16],组织电阻一定会发生改变,这样,通过组织电阻发生的变化就可以直接地反映植物的抗逆程度或受害程度。从理论上说利用电阻抗法可以研究植物抗逆性且比其他方法更为直观和灵敏。
(2)在外电场作用下,植物组织中存在电极化损耗,损耗角正切tgδ、电导率γ、频率ω、复介电常数的实部ε′、复介电常数的虚部ε″之间以及它们与温度之间存在关联性。目前的研究文献来看,没有考虑这些物理量与温度之间的关联性,以及这些物理量之间的关联性。所以,在利用电阻抗法研究植物的抗逆性时,参数应该综合考虑 。
(3)一方面,媒质属于电介质还是良导体,不仅与媒质参数有关,而且与频率有关。随外加电场频率的增加,生物组织介电常数从高值向低值变化和电导率随外加电场频率的增加从低值向高值变化的效应[17],另一方面,由于参数 ε′、ε″、tgδ、γ均与频率ω有关,所以在利用阻抗法研究物植物的抗性时,需要说明所测参数的外场频率。
(4)在高频场作用下,导电媒质电导率和介电常数的总的宏观效应引入等效的复介电常数表示,由前面的表达式可知,电导率变成等效的复介电常数的虚数组成部分。所以,在利用电阻抗研究植物的抗逆性时,植物组织的介电常数应该是比电导率更能够可靠和准确地反映植物组织的抗逆程度。这需要进一步的实验验证。
(5)电阻率rx不及tgδ更能够准确地反映植物的特性。这也需要进一步的实验验证。由于温度显涵于弛豫(松弛)时间的表达式中,利用阻抗法研究林木或植物与温度的关系时,是一个重要的参数,不可缺失。刘晓红的实验结论是,电阻抗图谱参数尚不能用于表征小麦品种的抗旱性[11]。作者从物理学的视野分析,因为在利用该方法,建立阻抗模型时是把待测的对象——生物组织看作了一个圆柱形的电容器,而林木的茎和针叶又是圆拄形的,所以实验样本取材茎和针叶,实验效果较好。如果在测定叶片时修改电容器的模型为平板电容器,作者推断实验效果也许是可行的。建议我们利用阻抗技术研究植物的抗逆性时,电容器理论模型应根据具体的实验样品作相应的调整。
(6)由于植物组织属于一种非线性、各向异性的电介质,在不同的方向施加电场,其极化程度和极化方向是不一样的,所以在利用电阻抗研究植物的抗逆性时,需要研究所施加的电场的方位,例如电场的方向是平行还是垂直于组织。目前的研究文献来看,都没有考虑外加场的方位。由于并联支路的导纳可以直接相加,所以,在利用电阻抗研究植物的抗逆性时,在构建物理模型时,相信在某些情况下,若采用导纳进行计算更为方便。这需要进一步的实验验证。目前的研究文献来看,未见有关研究植物抗性的导纳法的应用。
(7)植物电阻抗成像技术或许是未来一个具有潜在价值和奥妙的新兴研究点。生物电阻抗成像技术(biological electrical im pedance im aging,BEI)是医学成像技术的一个新方向,是当今生物医学工程学的重要研究课题之一。已开始从实验室研究向临床应用过渡[18]。与之相比,研究植物的生物电阻抗成像技术未见相关文献报道。电极系统的选择也是应用中的一个关键问题。
[1]喻方圆,徐锡增.苗木生理与质量研究进展[J].世界林业研究,2000,13(4):17~24.
[2]张 钢,肖建忠,陈段芬.测定植物抗寒性的电阻抗图谱法[J].植物生理与分子生物学学报,2005,31(1):19~26.
[3]Repo T,Leinonen M,Pääkkönen T.The application of electrical impedance spectroscopy in assessing the frost hardiness of Scots pine[O].//ICPRBI.Proceedingsof the IX International Conferen ce on Electrical Bio-Impedance.H eidelberg:ICPRBI,1995.
[4]王爱芳,张 钢,魏士春,等.不同发育时期樟子松的电阻抗参数与抗寒性的关系[J].生态学报,2008,28(11):5741~5749.
[5]Man cuso S,Nicese F P,Masi E,et al.Comparing fractal analysis,electrical im pedance and electrolyte leakage for the assessm ent of cold tolerance in Callistem on and Grevillea sp p[J].H orticulturalScience and Biotechnology,2004,79(4):627~632.
[6]H urme P,Repo T,Savolainen O,etal.Climatic adaptation of bud set and frost hardiness in Scots pine(Pinus sy lvestris)[J].Canadian Journal of Forest Research,1997,27(5):716~723.
[7]Repo T,Zhang G,RyyppöA,et al.The electrical im pedance spectroscopy of Scots pine(Pinus sylvestris L.)shoots in relation to cold acclim ation[J].Jou rnal of Experimen tal Botany,2000,51(353):2095~2107.
[8]Repo T.Influence of different electrodes and tissues on the impedance spectra of Scots pine shoots[J].E lectro and M agnetobiology,1994,13(1):1~14.
[9]RyyppöA,Repo T,Vapaavuori E.Developmen t of frost hardiness in roots and shoots of Scots pine seedlings at non-freezing temperatu res[J].Canadian Journal of Fo rest Research,1998,28(4):557~565.
[10]刘 辉,张 钢.短日照对金叶女贞茎抗寒性和电阻抗图谱参数的影响[J].华北农学报,2008,23(2):173~179.
[11]刘晓红,王国栋,张 钢.小麦叶片的电阻抗图谱参数对水分胁迫的响应[J].兰州大学学报:自然科学版,2007,43(5):48~53.
[12]孙目珍.电介质物理基础[M].广州:华南理工大学出版社,2000.
[13]陈季丹,刘子玉.电介质物理学[M].北京:机械工业出版社,1982.
[14]殷之文.电介质物理学[M].北京:科学出版社,2003.
[15]马冰然.电磁场与微波技术(上册)[M].广州:华南理工大学出版社,1999.
[16]郝建军,康宗利.植物生理学[M].北京:化学工业出版社,2005.
[17]马 青.频域阻抗法研究细胞介电特性[J].中国医学物理学杂志,2004,21(4):187~195.
[18]董秀珍.生物电阻抗成像研究的现状与挑战[J].中国生物医学工程学报,27(5):641~643.
