鳗弧菌W-1金属蛋白酶基因真核表达载体的构建及其在动物细胞中的表达
2010-06-04杨慧,陈吉祥,张晓华等
鳗弧菌(Vibrioanguillarum)是危害鱼类的主要的条件性病原菌之一,在世界各地流行,也是我国海水养殖鱼类的重要病原菌,给水产养殖业造成了较大的危害[1,2]。鳗弧菌的致病性与许多毒力因子有关,除了由65 kb质粒pJM1编码的铁吸收系统[3,4]、细胞外溶血毒素[5,6]、脂多糖[7]、细菌鞭毛蛋白[8,9]等毒力因子外,胞外蛋白酶也被认为是鳗弧菌与其它许多细菌病原的毒力因子[10~16]。Milton等[14]首先从鳗弧菌 NB10中克隆了金属蛋白酶基因,编码 611个氨基酸残基组成的多肽链,完整蛋白的分子量为 66.7 kDa。
陈吉祥等[15]从山东沿海养鱼场发病鱼分离的致病性鳗弧菌 W-1的胞外产物中分离纯化了一种金属蛋白酶,并在原核细胞中克隆、表达了该酶的基因——empA基因[17],同时对其活性中心的氨基酸残基进行了定点突变[18]。序列分析结果发现,其序列同源性较高,活性位点氨基酸序列与其它病原弧菌有较大的相似度,不同血清型的鳗弧菌其活性中心的氨基酸残基相同;活性实验结果表明,突变的金属蛋白酶丧失其蛋白水解活性和细胞毒性,但仍具有较强的免疫原性,因此是一个颇具潜力的鳗弧菌疫苗候选抗原蛋白。
作者在此利用分子生物学技术将突变的鳗弧菌W-1金属蛋白酶基因m-empA7克隆入真核表达载体中,使其在真核细胞中得到表达,为进一步研究该疫苗在动物实验中的免疫效果以及研制鳗弧菌金属蛋白酶基因疫苗奠定基础。
1 实验
1.1 材料
质粒pET24d(+)/m-empA7、兔抗鳗弧菌W-1金属蛋白酶抗体,自制;pCDNA3.1(+)、Trizol Reagent,Invitrogen公司;大肠杆菌JM109、真核细胞系CHO和HEK293T,中国海洋大学海洋生命学院;ExTaq DNA聚合酶、T4 DNA连接酶、dNTP、IPTG、各种限制性内切酶、Wide Range DNA Ladder Marker、DNA凝胶回收纯化试剂盒,大连宝生物工程有限公司;辣根过氧化物酶标记的山羊抗兔IgG,武汉博士德公司;除内毒素质粒小量提取试剂盒,北京天根生物工程有限公司;磷酸钙法质粒转染试剂盒,碧云天生物工程有限公司;逆转录试剂盒,Promega公司;MEM、DMEM细胞培养基、D-Hank′s平衡液、胎牛血清和胰蛋白酶,Gibco公司;其它试剂均为国产生化试剂或分析纯试剂。
1.2 方法
1.2.1 重组真核表达载体的构建及鉴定
根据已突变的鳗弧菌W-1金属蛋白酶基因m-empA7的碱基序列[18]及真核表达载体pCDNA3.1(+)多克隆位点的序列,设计一对特异性引物(VP61:5′-CGGGATCCATGAAAAAAGTACAACGTCAA-ATG-3′,VP62:5′-CCGCTCGAGTTAATCCAGTC-TTAACGTTACACC-3′)。以pET24d(+)/m-empA7突变质粒为模板,以VP61和VP62为引物,PCR扩增得到5′和3′端分别带有BamHI和XhoI酶切位点的突变的金属蛋白酶基因m-empA7。PCR产物和真核表达载体pCDNA3.1(+)分别用XhoI和BamHI双酶切,回收纯化后加T4 DNA连接酶于16℃连接16 h,氯化钙法转化E.coliJM109,挑取Amp抗性的单菌落鉴定,筛选阳性克隆,得到的重组真核表达质粒命名为pCDNA3.1(+)/m-empA7。酶切、连接、转化以及阳性重组子的筛选和鉴定均按《分子克隆实验指南》进行。
1.2.2 真核细胞转染
除内毒素质粒DNA的提取方法按照Promega公司的除内毒素质粒提取试剂盒说明书进行,用紫外分光光度计法测定质粒DNA浓度和纯度。细胞转染的方法按照碧云天生物工程有限公司的磷酸钙法细胞转染试剂盒说明书进行。转染前24 h,通过胰酶消化收集细胞,用完全培养基(含抗生素)将细胞平铺于6孔板上,于含5% CO2的37℃培养箱中孵育20~24 h(细胞汇合度在70%~80%为宜)。转染前30 min,吸去细胞培养液,加入2 mL新鲜的不含抗生素的完全培养基。取2 μg待转染的质粒,加入到100 μL CaCl2溶液中,混匀。均匀滴加DNA-CaCl2-BBS的混合物到整个6孔板内,轻轻旋转以混匀。在含3%CO2的37℃培养箱中培养24 h。然后轻轻晃动培养板数次以充分悬浮一些磷酸钙沉淀。吸去含磷酸钙沉淀的培养液,加入2 mL新鲜的完全培养基继续培养。在转染约36 h后检测转染基因的表达。
1.2.3 RT-PCR检测转染细胞中m-empA7基因的转录
细胞转染36 h后收集细胞,按Trizol Reagent说明书提取细胞中总RNA,用逆转录试剂盒以提取的RNA为模板,反转录合成cDNA第一链。根据突变的金属蛋白酶基因m-empA7的序列,在Genbank中进行序列同源比对,选择保守区段的第1485~1731核苷酸序列,设计一对特异性引物(VP51:5′-CCTTTAACCAAGTGGGCGTA-3′,VP52:5′-CGATTTGTAAGGGCGACAAT-3′),扩增目标产物为248 bp。以该引物进行反转录聚合酶链反应(Reverse transcription PCR, RT-PCR)检测m-empA7基因的转录水平。PCR反应条件为:94℃预变性5 min;94℃变性1 min,52℃退火1 min,72℃延伸1 min,30个循环;72℃延伸10 min。PCR反应产物用1%琼脂糖凝胶电泳检测。
1.2.4 Western-blot检测转染细胞中m-empA7基因的表达产物
细胞转染36 h后收获6孔组织培养板中的细胞,分别加入上样缓冲溶液,煮沸10 min,用10% SDS-PAGE进行电泳。然后用半干法将蛋白转印到NC膜上,用含5%脱脂奶的PBST(含0.05% Tween20 1×PBS)4℃封闭过夜。以兔抗EmpA蛋白的多克隆抗体[17]作为一抗,以辣根过氧化物酶标记的山羊抗兔IgG作为二抗,利用DAB进行显色,检测转染细胞中m-empA7基因的表达产物。
2 结果与讨论
2.1 重组真核表达质粒pCDNA3.1(+)/m-empA7的构建及鉴定
突变的鳗弧菌W-1金属蛋白酶基因m-empA7经过PCR扩增后,得到了约1.8 kb的片段(图1),PCR产物经酶切后插入到pCDNA3.1(+)中,构建了重组真核表达质粒pCDNA3.1(+)/m-empA7,经XhoI和BamHI双酶切后凝胶电泳,在1836 bp处有一目的条带出现(图2)。

M. DL2000 DNA Marker 1,2. m-empA7 Gene amplified by PCR

M1. λ-HindⅢ digest 1. pCDNA3.1(+) 2. pCDNA3.1(+)/m-empA7 3. m-empA7 Gene amplified by PCR M2. DL2000 DNA Marker
经酶切鉴定的阳性重组质粒由大连宝生物工程有限公司进行克隆片段的DNA序列测定,测序结果用http://www.ebi.ac.uk/clustalw/软件与m-empA7的核酸序列进行比对,结果证实它们的序列完全一致。
2.2 转染细胞中m-empA7基因的转录
采用磷酸钙法将重组质粒pCDNA3.1(+)/m-empA7转染至CHO和HEK293T细胞中,同时以转染空质粒pCDNA3.1(+)的细胞作空白对照、以未转染质粒的正常细胞作阴性对照,在细胞转染36 h后,提取总RNA进行RT-PCR检测,结果如图3所示。

M. 100 bp DNA Ladder marker 1. CHO Transfected with pCDNA3.1(+) 2. CHO Untransfected 3. CHO Transfected with pCDNA3.1(+)/m-empA7 4. HEK293T Transfected with pCDNA3.1(+) 5. HEK293T Untransfected 6. HEK293T Transfected with pCDNA3.1(+)/m-empA7
由图3可看出,在重组质粒转染的CHO细胞和HEK293T细胞中都扩增出了一条约248 bp的小目的片断,说明m-empA7基因的mRNA在真核细胞中得到了转录。而转染空质粒pCDNA3.1(+)和未转染质粒的对照细胞中均未扩增得到目的片断。
2.3 转染细胞中m-empA7基因的表达
以转染空质粒pCDNA3.1(+)的细胞作空白对照、以未转染质粒的正常细胞作阴性对照,在细胞转染36 h后,收集细胞并裂解进行Western-blot检测,结果如图4所示。
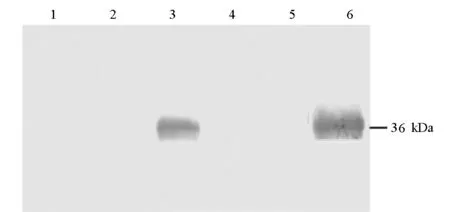
1. CHO Transfected with pCDNA3.1(+) 2.CHO Untransfected 3. CHO Transfected with pCDNA3.1(+)/m-empA7 4. HEK293T Transfected with pCDNA3.1(+) 5.HEK293T Untransfected 6. HEK293T Transfected with pCDNA3.1(+)/m-empA7
由图4可看出,在重组质粒转染的CHO细胞和HEK293T细胞中都检测到了一条分子量约36 kDa的特异性免疫反应带,比由m-empA7碱基序列推测的蛋白质分子量(66.7 kDa)小得多,这与在原核细胞中表达的情况相一致[17,18],说明目的基因能够在细胞内正确转录、翻译、表达,并可与抗EmpA蛋白的抗体特异性结合。而转染空质粒pCDNA3.1(+)和未转染质粒的对照细胞中未见有免疫反应带。
2.4 讨论
DNA 疫苗是编码免疫原或与免疫原相关的真核表达质粒DNA(或RNA),它可经一定途径进入动物体内,被动物宿主细胞摄取后转录和翻译表达出抗原蛋白,此抗原蛋白能够刺激机体产生非特异性和特异性两种免疫应答反应,从而起到免疫保护作用。早期DNA疫苗的研究主要是以哺乳动物为模型,鱼类DNA 疫苗的研究起步较晚。据统计,每年大约有10%的养殖鱼类死于传染性疾病,而大规模的爆发性疾病会引起更严重的损失[19]。因此,开展鱼类DNA疫苗的研究是非常必要的。1991年,Hansen等[20]首先报道了外源基因能够在鲤鱼肌肉组织中表达。1996 年,Anderson等[21]第一次报告了带有IHNV糖蛋白基因的质粒能够在虹鳟中表达,并且能够保护IHNV病毒对受免疫鱼的感染。随后的一些实验也证明,在多种鱼体内,外源基因不仅能够高效、持久地表达,而且能够引起宿主鱼类对外源蛋白的免疫应答。这些为构建鱼类DNA 疫苗提供了基础。
病原菌的致病性是由一些可遗传的毒力因子所决定,其中胞外蛋白酶被认为是许多病原菌的一个重要毒力因子[10~16]。 不同弧菌胞外蛋白酶氨基酸序列保守性很高,且都具有活性位点,通过各自不同的方式发挥作用,致使受感染动物发病或死亡,但同时这些毒素又是良好的抗原,能诱发机体产生强烈的免疫反应,因此常作为首选的疫苗抗原,在DNA疫苗中有较好的应用。
作者所在研究组从鳗弧菌W-1中分离得到高活性的蛋白酶,通过对其理化性质及末端氨基酸序列分析发现,该蛋白酶是一种金属蛋白酶,由611个氨基酸残基组成,完整蛋白的分子量为66.7 kDa[15]。对来源于不同养殖环境致病菌的蛋白酶基因进行克隆和序列分析发现,其序列同源性较高,通过对比发现酶的可能活性位点氨基酸序列与其它病原弧菌有较大的相似度,不同血清型的鳗弧菌其活力中心的氨基酸残基相同,但有关酶活性位点特定氨基酸残基在维持酶活力和对细胞的毒性作用等方面,仍缺乏深入研究。
研究组在原核细胞中克隆、表达了鳗弧菌W-1金属蛋白酶基因empA基因,并对其活性中心的氨基酸残基进行了定点突变。活性实验结果表明,将empA基因活性中心的Glu347突变为Lys后,表达的突变蛋白m-EmpA7完全丧失了其蛋白水解活性和细胞毒性,但仍保持了其良好的免疫原性[18],因此,可将没有蛋白水解活性和细胞毒性的突变蛋白作为首选的疫苗抗原在真核细胞中表达,并在此基础上进行鱼类鳗弧菌病DNA疫苗的研制。
3 结论
利用分子生物学技术,在已克隆的突变的鳗弧菌W-1金属蛋白酶基因m-empA7的基础上,成功构建了鳗弧菌突变金属蛋白酶m-EmpA7的真核表达质粒pCDNA3.1(+)/m-empA7,转染细胞后经 RT-PCR和Western-blot检测表明,m-empA7基因能够在真核细胞中转录和表达,而且表达的目的蛋白可与抗EmpA蛋白的抗体特异性结合,具有抗原性,因此,可考虑将pCDNA3.1(+)/m-empA7作为基因候选疫苗用于后续的动物实验研究。这为进一步研究鳗弧菌金属蛋白酶基因疫苗在鱼类弧菌病害防治中的作用奠定了一定的基础。
参考文献:
[1] Egidius E. Vibriosis: Pathogenicity and pathology. A review[J]. Aquaculture, 1987, 67: 15-18.
[2] Austin B,Austin D A.Bacterial Fish Pathogens:Diseases of Farmed and Wild Fish[M]. London, United Kingdom:Springer, 1999.
[3] Crosa J H. A plasmid associated with virulence in the marine fish pathogenVibrioanguillarumspecifies an iron-sequestering system[J].Nature, 1980, 284(5756): 566-568.
[4] Actis L A, Tolmasky M E, Farrell D H, et al. Genetic and molecular characterization of essential components of theVibrioanguillarumplasmid-mediated iron-transport system[J]. J Biol Chem,1988, 263(6): 2853-2860.
[5] Munn C B. Hemolysin production byVibrioanguillarum[J].FEMS Microbiol Lett, 1978,3(5): 265-268.
[6] Munn C B. Production and Properties of a Hemolytic Toxin byVibrioanguillarum. Fish Diseases[M]. Berlin: Third copraq session, Springer-Verlag, 1980: 69-74.
[7] Boesen H T, Pedersen K, Larsen J L, et al. Vibrio anguillarum resistance to rainbow trout (Oncorhynchusmykiss) serum: Role ofO-antigen structure of lipopolysaccharide[J]. Infect Immun, 1999, 67(1): 294-301.
[8] Singer J T, Schmidt K A, Reno P W. Polypeptides p40, pOM2, and pAngR are required for iron uptake and for virulence of the marine fish pathogenVibrioanguillarum[J]. Bacteriology, 1991, 173(3): 1347-1352.
[9] Norqvist A, Wolf-Watz H. Characterization of a novel chromosomal virulence locus involved in expression of a major surface flagellar sheath antigen of the fish pathogenVibrioanguillarum[J]. Infect Immun, 1993, 61(6): 2434-2444.
[10] Inamura H, Nakai T, Muroga K. An extracellular protease produced byVibrioanguillarum[J]. Bull Jpn Soc Sci Fish, 1985,51(12): 1915-1920.
[11] Kanemori Y, Nakai T,Muroga K. The role of extracellular protease produced byVibrioanguillarum[J].Fish Pathology, 1987, 22(3): 153-158.
[12] Norqvist A, Norrman B, Wolf-Watz H. Identification and characterization of a zinc metalloprotease associated with invasion by the fish pathogenVibrioanguillarum[J]. Infect Immun, 1990,58(11): 3731-3736.
[13] Farrell D H, Crosa J H. Purification and characterization of a secreted protease from the pathogenic marine bacteriumVibrioanguillarum[J]. Biochem, 1991, 30(14): 3422-3436.
[14] Milton D L, Norqvist A, Wolf-Watz H. Cloning of a metalloprotease gene involved in the virulence mechanism ofVibrioanguillarum[J]. J Bacteriol, 1992, 174(22): 7235-7244.
[15] 陈吉祥,刘霜,李筠,等.一种致病性鳗弧菌胞外蛋白酶的分离纯化及性质[J].中国水产科学,2002,9(4):318-322.
[16] Denkin S M, Nelson D R. Regulation ofVibrioanguillarumempAmetalloprotease expression and its role in virulence[J]. Appl Environ Microbiol, 2004, 70(7): 4193-4204.
[17] Yang Hui,Chen Jixiang, Yang Guanpin, et al. Characterization and pathogenicity of the zinc metalloprotease EmpA ofVibrioanguillarumexpressed inEscherichiacoli[J]. Current Microbiology, 2007,54(3):244-248.
[18] Yang Hui,Chen Jixiang,Yang Guanpin, et al. Mutational analysis of the zinc metalloprotease EmpA ofVibrioanguillarum[J]. FEMS Microbiology Letters, 2007, 267(1): 56-63.
[19] Heppell J,Davis H L. Application of DNA vaccine technology to aquaculture[J].Adv Drug Delivery Rev, 2000, 43(1): 29-43.
[20] Hansen E, Fernandes K, Goldspink G, et al. Strong expression of foreign genes following direct injection into fish muscle[J]. FEBS Lett, 1991, 290(1-2): 73-76.
[21] Anderson E D, Mourich D V, Fahrenkrug S C,et al. Genetic immunization of rainbow trout (Oncorhynchusmykiss) against infectious hematopoietic necrosis virus[J]. Mol Marine Biol Biotechnol, 2000, 5(2): 114-122.
