城市景观红竹林无性系分株生长特征
2010-06-04虞敏之陈双林郑连喜郭子武黄树荣邵宏军汪忠华
虞敏之,陈双林,郑连喜,郭子武,黄树荣,邵宏军,汪忠华
(1. 浙江省龙游县林业局,浙江 龙游 324400;2. 中国林业科学研究院亚热带林业研究所,浙江 富阳 311400)
红竹(Phyllostachys iridenscens)自然分布于浙江、江苏、安徽等地,具有生态适应性强,成林速度快,秆形高大(秆高8 ~ 12m,径可达10 cm),竹笋产量高,品质佳,竹材劈篾性好,经晒不裂等特点,是笋、材皆可利用的优良经济竹种[1]。可实施林地有机材料覆盖措施,促进笋芽提早萌发,显著提高竹林经济产出[2]。在我国一些主要竹产区有规模化栽培,已成为区域竹产业发展的重要资源。龙游县属刚竹属竹种中心分布区,红竹是20世纪90年代逐渐发展起来的主栽笋用竹种,全县有红竹林面积0.7万余hm2,已成为区域农村社会经济发展的重要资源和农民家庭经济收入的主要来源,也是龙游县建立地方特色竹产业的重要组成部分,并在城镇绿化中得到了较大规模的应用。
红竹形态优美,枝叶茂盛,竹叶碧绿,箨鞘紫红色,箨叶为颜色鲜艳的彩带状,边缘桔黄色,中部绿、紫色,也是优良的观赏竹种。红竹作为一种重要的生态经济竹种,开展的相关研究较多,涉及生长发育[3]、生产力评价[4]、林地养分补充[5~6]和有机材料覆盖[2]、新造林地上部分林分结构变化[7]和地下鞭系结构变化[8]、叶片叶绿素含量[9]等。但就城市景观绿化红竹林的相关研究尚无涉及。本文对立地条件、初植密度、栽培管理措施相同的,大密度全梢竹一次成景造林的2种造林年份的城市景观红竹林进行了连续3年的新竹数、新竹胸径和枝下高、全高等因子的调查,以分析红竹林立竹生长特征,旨在为城市景观红竹林良好更新生长、景观维护提供参考。
1 材料与方法
1.1 试验地概况
试验地位于浙江省龙游县城,属中亚热带气候,四季分明,年均气温17.3℃,极端高温41℃,极端低温-11.4℃,年均无霜期261.5d,年均降水量1621.9 mm,年均蒸发量1 392 mm,年平均相对湿度80%。地势平缓,土壤为红壤,土层厚度100 cm以上,速效N、P、K含量中等。
1.2 试验竹林
试验红竹林位于龙游县城江滨公园,分别于2003年、2005年11月营造,面积分别为0.3、0.2 hm2。为达到一次成景效果,采用大密度全梢竹造林方法,初植密度2 ~ 3株/m2。母竹采自未进行过林地有机材料覆盖的红竹林,选择1年生生长健壮,胸径2 cm左右,无病虫为害的立竹,带30 cm以上土球,有来鞭、去鞭。母竹不去梢,栽植前剪去母竹2/3枝叶。栽植后1周内,晴天每天浇水补充土壤水分。试验林自然生长,不实施土壤养分补充和林地垦复,不采笋、伐竹,仅在成林前(栽植后2 a)每年8月进行1次人工除草。栽植后第3年试验林立竹郁闭成密集状,不包括母竹的立竹密度达30 000株/hm2以上。
1.3 调查方法
2009年12月在2种造林年份的试验红竹林中分别随机设置3个5 m×5 m样地,调查样地2007-2009年出笋成竹的新竹数、新竹胸径和枝下高、全高。年度新竹数按照立竹秆色来分辨调查,立竹胸径用Guanglu电子数显游标卡尺(分辩率0.01 mm)测量,立竹枝下高用钢卷尺测量,立竹全高用测高仪测量,精度均为0.01 m。
1.4 数据处理
试验数据在Excel 2003统计软件中进行整理和作图,在DPS.v2.0统计软件中进行LSD法多重比较,分析不同造林年份不同年度新竹数、新竹胸径、枝下高、全高的差异性。为避免立竹胸径差异对立竹枝下高、全高的比较影响,以立竹胸径为自变量,立竹枝下高、全高为因变量,建立2种造林年份各个年度的新竹枝下高、全高与新竹胸径的线性回归关系方程,再分别将6个样地各个年度的新竹平均胸径代入方程,对所得的数据组进行差异性分析。
2 结果与分析

图1 城市景观红竹林年度新竹数变化Figure 1 New bamboo amount of different year in Ph. iridescens plantation for urban landscape
2.1 年度新竹数
由图1可知,2005年冬季造林的试验林2007-2009年林分均有大量出笋成竹,虽然 2009年新竹数较前 2年有12.4% ~ 18.1%幅度的下降,但年度间新竹数并无显著差异。2003年冬季造林的试验林 2008-2009年新竹数无显著差异,但均较2007年新竹数显著减少,降幅达31%以上,并呈随造林年限的延长,林分新竹数逐年下降的趋势。比较2个造林年份试验林,2003年造林林分2007年新竹数与2005年造林林分2007-2009年新竹数差异不显著,即造林后出笋成竹的前3年林分出笋成竹能力无显著差异,2008年新竹数显著下降,即造林后出笋成竹的第4年是林分新竹数显著变化的转折点。可以认为,高密度造林的红竹景观林从造林后第5年开始,林分出笋成竹能力显著降低,这与林分密度逐年大幅度提高,地下鞭系延伸生长空间受到限制,林内光照不足,无性系分株间竞争加剧,林分立竹年龄结构不合理,逐渐趋于老化状态等有关,影响了竹林自我更新和景观效果维持。2005年龙游县夏秋季出现严重干旱,影响了2006年的出笋成竹,但2007年仍有较大的新竹数,并且较距离造林年份相同年限的2005年造林林分2009年新竹数大,这与竹林林分密度相对较小,林内环境较好,鞭系生长空间较大,养分供应相对充足,营养退笋减少等有关,也说明红竹生态适应性强,环境资源利用效率高。
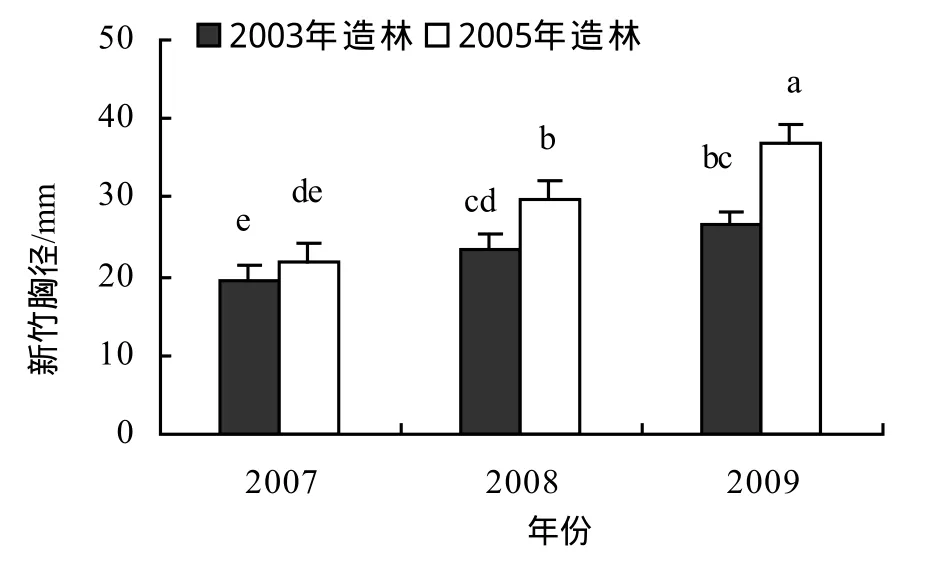
图2 城市景观红竹林年度胸径变化Figure 2 DBH of new bamboos of different year in Ph. iridescensplantation for urban landscape
2.2 立竹胸径
由图2可知,2003年、2005年造林的试验林年度新竹胸径均有逐年增大的趋势,造林后第6年高密度一次成景造林的景观红竹林虽然新竹数减少,但新竹胸径仍在增粗。2005年造林试验林2007-2009年新竹胸径均有显著差异,2008年新竹胸径比2007年新竹胸径增大36.2%,2009年又比2008年增大23.4%。2003年造林试验林2008-2009年新竹胸径显著大于2007年,2008-2009年间差异缩小,2008年新竹胸径比2007年新竹胸径增大21.7%,2009年比2008年增大13.7%,说明随着造林年限的延长,新竹胸径年际间增大的幅度逐渐减小,并趋于相对稳定。造林后第1个出笋成竹年新竹胸径较母竹小,但第2个出笋成竹年新竹胸径即可达到母竹胸径。2003年造林林分2007年新竹与2005年造林林分2009年新竹距离造林年份的年限相同,但新竹胸径差异极显著(p < 0.01),这与2005年夏秋季严重干旱,影响了2006年的成竹质量有关。试验林年度新竹最大平均胸径为36.84 mm,个体最大胸径49.51 mm,表明如进行适当的林分结构调控,控制林分密度,个体径级仍有较大的增长空间。
2.3 立竹枝下高
由表1可知,2003年造林试验林2007年、2009年的新竹枝下高差异显著,其他年度间新竹枝下高无显著差异。2005年造林试验林2007-2009年新竹枝下高均有显著差异。比较2种造林年份试验林,2005年造林试验林2009年新竹枝下高与2003年造林试验林3个年度的新竹枝下高均差异显著,2008年新竹枝下高与2003年造林试验林2009年新竹枝下高无显著差异,与2007-2008年新竹枝下高差异显著,2007年新竹枝下高与2003年造林试验林3个年度的新竹枝下高均无显著差异。已有研究表明,立竹枝下高是立竹胸径的从属因子[10],而2种造林年份试验林不同年度间新竹枝下高、新竹胸径差异性不一致,除立竹胸径对立竹枝下高的决定性影响外,是否不同年度差异显著的林分密度对新竹胸径也可能产生一定的影响呢?
为避免立竹胸径差异对立竹枝下高的比较影响,建立了 2种造林年份不同年度的新竹胸径(D)与枝下高(h)的关系方程:

式1 ~ 3、式4 ~ 6分别是2003年、2005年造林试验林2007-2009年新竹胸径与枝下高的关系方程。将6个样地 2007-2009年新竹平均胸径分别代入方程式,得到相同胸径条件下的不同年度新竹枝下高数据组。经LSD法方差分析,在相同的立竹胸径条件下,2种造林年份不同年度的立竹枝下高并无显著差异(表2),可见,在试验林林分密度范围内,立竹枝下高是由红竹固有的生物学和生态学特征所决定的。

表1 城市景观红竹林立竹枝下高、全高变化Table 1 Total height and clear bole height of Ph. iridescens plantation for urban landscape

表2 城市景观红竹林相同胸径条件下立竹枝下高、全高变化Table 2 Variation on total height and clear bole height of Ph. iridescens with the same DBH
2.4 立竹全高
2种造林年份试验林3个年度的新竹全高差异性与新竹枝下高总体上相同,主要差别在于2003年造林试验林2007年新竹全高与2008-2009年新竹全高差异显著,与各年度的立竹胸径差异性一致,说明立竹全高是立竹胸径的从属因子。与立竹枝下高相同的方法建立立竹胸径与立竹全高的关系方程,得出试验林不同年度相同立竹胸径条件下的立竹全高(表2),从中可知,2种造林年份试验林不同年度相同胸径立竹的全高总体上无显著差异,其中,2005年造林试验林2007年、2009年新竹全高差异显著,这可能与林分密度和竹林养分供应状况有关。

式7 ~ 9、式10 ~ 12分别是2003年、2005年营造的试验林2007-2009年新竹胸径(D)与全高(H)的关系方程。
3 结论与讨论
红竹与散生类竹种和一些克隆植物一样,其盘根错节的地下鞭系具有强烈的生理整合能力[11~14],营养物质通过分株间的转移使资源利用有效性提高,系统收益远超过耗费[15],并且能集团抵御不良环境。但是这种生理整合效果受制于系统结构状况,如无性系分株的繁殖力会随着密度的增加而降低[16]。本研究表明高密度造林的景观红竹林造林后前3个出笋成竹年新竹数无显著差异,而第4个出笋成竹年新竹数则显著下降,反映出林分密度对分株繁殖能力的影响,也说明为了促进景观红竹林的良好更新和景观效果维持,需要实施人工干扰措施,如疏笋、选择性伐竹等,如仍沿袭目前的自然生长状况,到一定年限后,竹林自我更新能力会进一步地急剧下降,甚至不出笋成竹,使竹林趋于老化和退化。红竹是中大型竹种,笋用林经营的立竹胸径一般为4 ~ 5 cm,但作为景观林,需要有较大的立竹胸径,才能充分体现出观秆、观笋的美学价值。本试验表明,景观红竹林造林后前3个出笋成竹年新竹胸径逐年显著增大,从第4个出笋成竹年开始,新竹胸径趋于相对稳定,试验林最大立竹胸径不超过5 cm,这也是受林分密度的影响,高林分密度使无性系分株间资源竞争激烈,地下鞭系向上层土壤分布,林内光照减弱,光合作用能力下降,并且竹林易滋生病虫害,这也说明景观红竹林实施林分结构调控的必要性。对此,需进一步开展包括立竹密度、立竹胸径和年龄结构等的景观红竹林合理的林分结构建立研究。
植物具有表型可塑性,器官在复杂环境中会产生一系列不同的相对适合的表现型潜能[17],表型可塑性是植物种群适应环境异质性的重要途径[18],植物在高林分密度条件下,一般具有促进高生长,抑制径级生长的趋势[19~20],较大密度的林分,在林分生长过程中会出现自然稀疏现象[21]。对于竹子而言,立竹枝下高、全高是反映表型可塑性的重要因子,竹林自然稀疏主要表现在出笋减少或退笋增多、新竹抽枝长叶不完全、弱小竹枯死、立竹中下部位枝叶自然整枝等。本试验表明2种不同造林年份试验林不同年度相同胸径的立竹枝下高、全高总体上无显著差异,反映出立竹枝下高、全高是立竹胸径的从属因子,而且除新竹数在造林后一定年限显著减少外,并没有出现其他的林分自然稀疏现象,是否试验林林分密度仍未达到发生立竹表型变化和自然稀疏的密度要求,需进一步开展研究。
[1] 朱石麟,马乃训,傅懋毅,等. 中国竹类植物图志[M]. 北京:中国林业出版社,1994. 128-129.
[2] 陈双林,吴柏林,张德明. 林地覆盖对红竹林温度和出笋时间分配的影响研究[J]. 林业科学研究,2004,17(专刊):90-94.
[3] 马乃训,黄树田,张文燕. 红竹生长发育的研究[J]. 林业科学研究,1990,3(4):358-362.
[4] 陈双林,董建新. 优良经济竹种红竹生产力评价[J]. 林业科技开发,2001,15(4):11-16.
[5] 蒋式洪,钟传声. 磷钾硼肥对红竹笋品质效应的研究[J]. 竹子研究汇刊,2000,19(1):48-51.
[6] 吴明,吴柏林,曹永慧,等. 不同施肥处理对笋用红竹林土壤特性的影响[J]. 林业科学研究,2006,19(3):353-357.
[7] 陈双林,洪游游,张德明,等. 退化红壤区笋用小径竹幼林结构变化规律研究[J]. 浙江林学院学报,2005,22(3):296-299.
[8] 洪游游,陈双林,吴柏林. 新造红竹林地下结构年际变化规律初步研究[J]. 浙江林业科技,2005,25(4):24-26.
[9] 李建华,杨清平,郭子武,等. 红哺鸡竹叶片叶绿素含量的影响因子初步研究[J]. 浙江林业科技,2009,29(4):65-68.
[10] 陈双林,吴柏林,虞敏之,等. 毛竹材积主要构件因子关系研究及材积表编制[J]. 福建林业科技,2008,35(2):30-33.
[11] 李睿,钟章成,M J A 维尔格. 毛竹的无性系生长与立竹密度和叶龄结构的关系[J]. 植物生态学报,1997,21(6):545-550.
[12] Marshall C. Source-sink relations of interconnected ramets[A]. Van Groenendael J,De Kroom H. Clonal growth in plants. regulation and function[M]. The Hague: SPB Acad Publ,1990. 23-41.
[13] 王昱生. 关于无性系植物种群整合作用研究的现状及其应用前景[J]. 生态学杂志,1994,13(2):57-50.
[14] 董文渊. 竹类无性系种群生态学研究现状及其应用前景[J]. 林业科学研究,2002,15(2):235-241.
[15] Stuefer J F,During H J,De Kroom H. High benefits of clonal integration in two stoloniferous species in response to heterogeneous light environments[J]. J Ecol,1994(82):511-518.
[16] De Kroom H,Kwant R. Density-dependent growth responses in two clonal herbs: regulation of shoot density[J]. Oecologia,1991(86):298-304.
[17] Dewitt T J. Costs and limits of pheno typic plasticity[J]. Trends Ecol Evol,1998,13(2):77-81.
[18] Hutchings M J,Price E, Sih A, et al. Dose physiological integration enable clonal herbs to integrate the effects of environmental hetero geneity?[J]. PlantSpec Biol,1993(8):95-105.
[19] 黄宝灵,吕成群,蒙钰钗. 尾叶桉人工林种群密度的研究[J]. 应用生态学报,2000,11(1):30-32.
[20] 朴顺姬,杨持,黄绍峰,等. 羊草种群密度与生长动态研究[J]. 植物生态学报,1997,21(1):60-66.
[21] 郑海水,曾杰,翁启杰,等. Nelder试验:大叶相思树生长与密度、年龄的相关研究[J]. 林业科学研究,1996,9(2):158-164.
