si RNA沉默 Annexin-1基因对膀胱癌 T24细胞顺铂耐药的影响
2010-05-31杨远航徐万海林相国李建章杨德君王晓民
高 琳 杨远航 徐万海 林相国 李建章 杨德君 王晓民
(哈尔滨医科大学附属第四医院泌尿外科,黑龙江 哈尔滨 150001)
膜联蛋白-1(Annexin-1)是上皮生长因子(EGFR)激酶的主要底酶,糖皮质激素刺激其产生和活性。Annexin-1增强感染局部的凋亡表明它对肿瘤的诊断和治疗有重要意义,因此提出该蛋白质与肿瘤细胞的活性调节相关〔1,2〕。Annexin-1与膀胱癌发生、发展密切相关。本研究针对 Annexin-1基因 mRNA序列设计合成与之互补的双链小分子干扰 RNA(siRNA),在脂质体介导下转染膀胱癌T24细胞系,观察对 Annexin-1基因 mRNA和蛋白表达及对顺铂敏感性的变化。
1 材料与方法
1.1 材料
1.1.1 细胞株 膀胱癌T24细胞株。
1.1.2 主要试剂 RNA干扰试剂盒(SilencerTMsiRNA Construction Kit)购自美国 Ambion公司。阳离子脂质体转染试剂盒 LipofectamineTM2000、Trizol、M-MLV购自 Invitrogen生命技术公司。RPMI1640、IMDM、胰蛋白酶购自 Gibco公司。
1.2 方法
1.2.1 siRNA的设计及合成 根据基因库中 Annexin-1基因序列(GenBank Accession No.NM000700)和 siRNA设计原则,应用 Ambion生物公司的设计软件(siRNA Target Finder and Design Tools),自 Annexin-1 mRNA的起始密码子开始,寻找“AA”二连序列,记下其 3′端的 19个碱基序列作为候选的 siRNA靶序列。将候选序列在基因库中用 Blast软件进行同源序列搜索,排除那些和其他基因编码序列或基因表达序列标签(EST)同源的候选序列。最终选择了 3段 21个碱基的siRNA序列(TⅠ 、TⅡ 、TⅢ),分别靶向 Annexin-1基因的 240-260(SⅠ),435-455(SⅡ)和 506-526(SⅢ)区域(表 1)。另外,根据转染后细胞形态学变化,设计与 TI(240 siRNA)序列含有同样核苷酸比例的随机对照 siRNA(240c1,240c2)做为阴性对照,用 Blast验证随机序列与基因库中其他已知人类基因序列没有同源性。利用针对三磷酸甘油醛脱氢酶(β-actin)基因的正、反义寡核苷酸模板作为 RNA干扰实验的阳性对照。设计的正反义寡核苷酸模板 3′端均加上 T7启动子引物 5′CCTGTCTC3′,由上海生工生物工程技术服务有限公司合成。
1.2.2 引物的设计与合成 设计了 1对 Annexin-1的引物,序列:上 游:5′-GACCACCTCAACTCAAGGAC-3′, 下 游:5′-AGGAAGCTGGATGAAGAGAC-3′,扩增片段长度 546 bp,同时设计了 1对管家基因 β-actin的引物作为内参照,序列:上游:5′-ACCACAGCTGAGAGGGAAATCG-3′, 下 游:5′-AGAGGTCCTTACG-GATGTCAACG-3′,扩增片段长度为 290 bp,由上海生工生物工程技术服务有限公司合成。
1.2.3 细胞培养 常规 RPMI1640培养,至转染前夜,细胞培养液更换为用无血清和无抗生素的 IMDM,置于 37℃、5%CO2孵箱,转染后用 20%胎牛血清(FBS)IMDM培养液,按常规方法培养,取处于对数生长期的细胞用于实验。用 Opti-MEMⅠ转染液及脂质体LipofectamineTM2000转染试剂,按试剂盒说明优化转染条件,分别将 3对 siRNA和阳性对照筛选出的干扰序列的scramble siRNA(siRNA-neg)以 400 nmol/L的终浓度加入细胞培养液,孵育 24、48、72 h后收获细胞进行检测。实验重复 3次。
1.2.4 半定量RT-PCR检测转染前后膀胱癌 T24细胞 Annexin-1 mRNA的表达水平 采用Trizol试剂盒提取细胞总RNA,用 2μg总 RNA进行 cDNA的合成,半定量 RT-PCR方法检测siRNA转染后不同时间点 24、48、72 h细胞中 Annexin-1 mRNA表达水平的变化,β-actin作内对照。PCR扩增条件Annexin-1:95℃预变性 5 min;94℃变性 40 s,58℃复性 1 min,72℃延伸1 min,共 33个循环;72℃延伸 10 min。 β-actin:94℃预变性5 min,94℃变性 1 min,63℃退火 1 min,72℃延伸 2 min,共 30个循环;72℃延伸 10 min。取适量 PCR扩增产物,在 2%琼脂糖凝胶电泳(80V,25min),溴化乙啶(EB)染色,紫外灯下观察结果并拍照。用凝胶成像分析仪进行半定量分析,以检测基因β-actin的相对亮度表示该基因 mRNA的相对表达量。
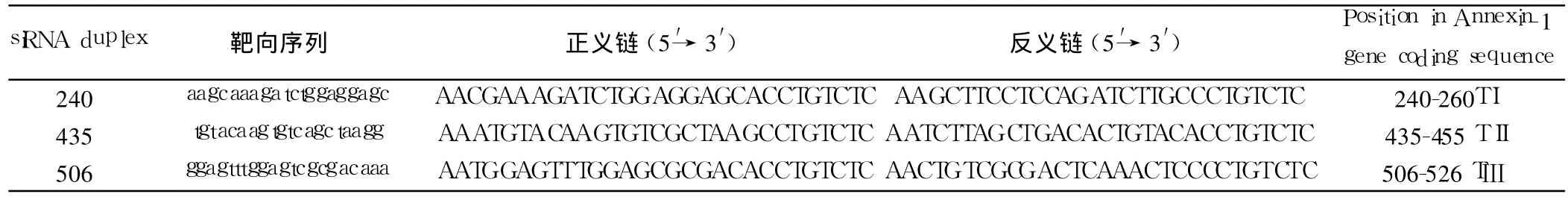
表1 Nucleotide sequences of the Annexin-1靶向siRNA的序列
1.2.5 Western印迹检测转染前后膀胱癌T24细胞 Annexin-1蛋白的表达 取状态良好对数生长期细胞 5×106个,用磷酸盐缓冲液(PBS)洗 2遍,于冰上加冷的细胞裂解液,制备细胞总蛋白,Lowry法蛋白定量。将样品置于 5×十二烷基硫酸钠(SDS)凝胶加样缓冲液中,100℃加热 10 min使蛋白质变性,聚丙烯酰胺凝胶电泳,蛋白免疫法(Western)印记,按 0.1 ml/cm2的量加入封闭液和适量稀释的抗体,于摇床上室温 2 h,PBS漂洗滤膜3次,加入辣根过氧化物酶标记二抗,将漂洗过的硝酸纤维素膜移至上述底物溶液中,轻轻摇动,蛋白条带的颜色达到要求,即用水冲洗,滤纸吸干,拍摄膜的照片,结果保存。内参对照使用小鼠抗人 β-actin(稀释度为 1∶10 000),以保证上样量一致。
1.2.6 siRNA和脂质体的细胞毒性检测 细胞用无血清和无抗生素的培养液洗涤重悬,每孔 2×104加入 96孔板。分别加入用 Opti-MEMⅠ稀释的 siRNA(终浓度为 100 nmol/L)或 Lipofectamine2000(终浓度为 2 ml/L),6 h后加入含 20%血清的培养液补足血清,72h后分别加入5 mg/ml的噻唑蓝(MTT),每孔20μl,37℃置 4h,弃上清,每孔加入 100μl二甲基亚砜(DMSO),待完全溶解后,测定其在 570nm的吸光度,并计算细胞存活率。对照孔细胞中加入等量的 Opti-MEMⅠ代替脂质体或SiRNA,其余培养条件与处理组相同。
1.2.7 MTT法检测细胞对顺铂敏感性的变化 取对数生长期的细胞,用含有 10%血清的RMPI1640培养液调整细胞浓度为1×105/ml,接种于 96孔培养板中,每孔 180μl,在 37℃,5%CO2条件下培养 24h。分组加药,每个浓度设3个平行孔,处理组加不同浓度的顺铂,阴性对照组加等体积的生理盐水,使每孔终体积为 200μl,培养 68 h后,每孔加入 5 mg/ml的 MTT 20μl,37℃继续培养 4h,2 000 r/min离心 10min,弃上清,每孔加入 100μl DMSO,振荡至沉淀完全溶解,在酶标仪上检测570nm光密度(OD)值。按以下公式计算肿瘤细胞生长抑制率。以同一药物的不同浓度对肿瘤细胞生长抑制率作图可得到剂量反应曲线,求出该药物的半数抑制浓度IC50,即细胞存活率减少 50%时的药物剂量。

1.3 统计学处理 应用 SPSS11.0软件进行统计分析,数据以x±s表示,两组均数间比较采用 t检验,多组均数间比较采用方差分析。
2 结 果
2.1 siRNA对膀胱癌 T24细胞 Annexin-1mRNA表达的影响经 AlphaEaseFCTM(Alpha Innotech)软件分析,以 40nmol/L浓度转染 T24细胞 24 h后 siRNAⅠ、Ⅱ、Ⅲ组 mRNA表达水平明显降低,在 24h下调(80.63±0.24)%,(t=474.3,P=0.001);在48 h下调(38.9±0.85)%,(t=64.833,P=0.01),72 h恢复正常。见图 1。
2.2 siRNA对膀胱癌 T24细胞Annexin-1蛋白表达的影响与随机对照的 siRNA相比,靶向Annexin-1的 siRNA转染后细胞内Annexin-1蛋白表达明显下降,并且与 RT-PCR结果一致。见图 2。
2.3 siRNA转染后膀胱癌 T24细胞对顺铂敏感性的变化MTT法显示,以 40nmol/L浓度转染 T24细胞 24 h后 siRNAⅠ、Ⅱ、Ⅲ组细胞对顺铂的 IC50值(μg/ml)均有不同程度的下降,从30.27下降到 7.81、6.94和 5.75,顺铂敏感性显著增加(P<0.05)。同时以不同浓度顺铂培养后细胞生存率亦明显下降,与对照组比较差异显著(P<0.05),见图 3。
2.4 siRNA和脂质体的细胞毒性检测结果 T24细胞经靶向Annexin-1基因的 siRNA或 oligolipofectamine处理后计算细胞存活率,与未处理的 T24细胞相比,没有显著统计学差异,显示siRNA及脂质体本身在实验条件下对细胞基本没有毒性作用。
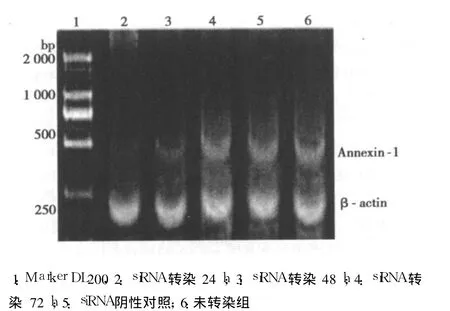
图1 siRNA对膀胱癌 T24细胞 Annexin-1mRNA表达的影响

图2 siRNA对膀胱癌 T24细胞 Annexin-1蛋白表达的影响
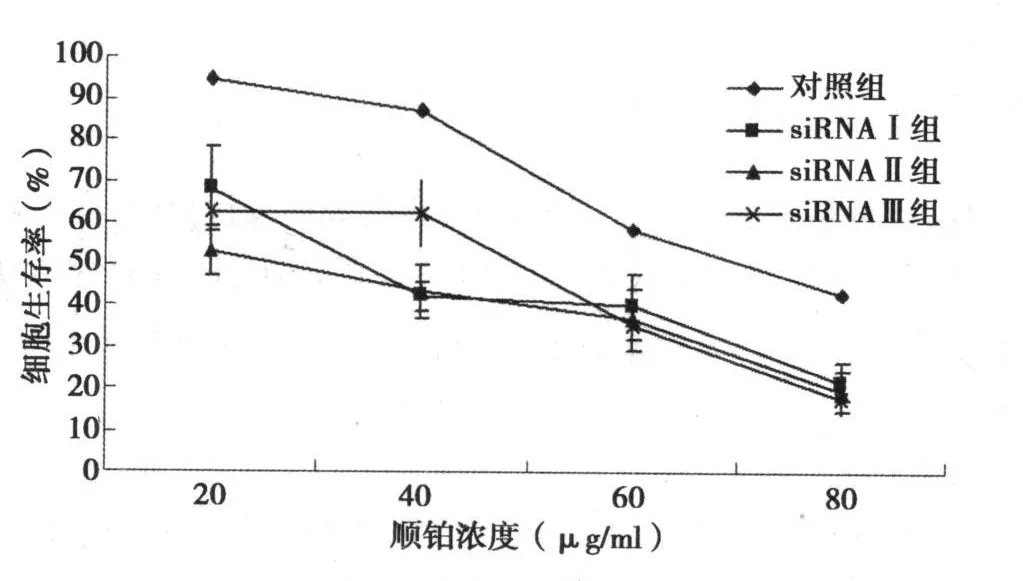
图3 不同浓度顺铂培养后T 24细胞生存率变化
3 讨 论
膀胱癌是泌尿系统中最常见的肿瘤之一,在美国其发病率居全身各部位肿瘤的第四位,在我国则排至第八位,近年来发病率呈上升趋势〔3〕。目前,就 Annexin-1的表达缺失是否与膀胱癌细胞的异常增殖潜力相关以及对化疗药物的敏感性尚未明了。
RNA干扰是由双链 RNA分子在mRNA水平关闭相应序列基因表达使其沉默的过程,是一种典型的转录后基因调控方法,此时启动子是活跃的,靶基因能够被转录,但不能正常累积mRNA。其优点首先表现在 siRNA只引起与其同源的mRNA降解,而其他基因的表达不受影响,因此具有高度的序列专一性。实验证实,导入细胞的 siRNA只能引起同源基因表达的抑制,而无关基因不受影响〔4〕。在 siRNA序列中配对的 19~21nt中如果只改变一个核苷酸,就可以使该 siRNA序列不对靶 mRNA起作用,证明RNA干扰有明显的特异性。其次RNA干扰是通过自身放大机制来发挥作用,极少量的dsRNA就能有效地抑制靶基因的表达,因此与反义寡核苷酸等基因封闭技术相比,具有高效性〔5〕。另外 RNA干扰操作简便快捷,利用RNA干扰技术,甚至可以在 1 w之内关闭 10个基因。
Elbashir等〔6〕首次报道 siRNA在哺乳动物体外培养细胞中能够成功的诱导特异基因受阻。作为一种有效抑制基因表达的方法,RNA干扰技术在近几年被广泛用于基因的功能研究。双链 siRNA的合成是进行 RNAi研究的关键〔7〕,本研究中采取了体外转录法,即在噬菌体RNA聚合酶、Klenow DNA聚合酶等作用下,以连有噬菌体启动子的线性DNA为模板,直接合成出特异双链siRNA。RNA聚合酶是体外转录合成 siRNA的关键酶,除了在本实验中用到的 T7 RNA聚合酶,其他通用的 RNA聚合酶还有 T3、SP6 RNA聚合酶,它们均是以 DNA为模板的RNA聚合酶。DNA模板必须连有特异的启动子序列,启动子区域是噬菌体 RNA聚合酶结合和 RNA开始合成的部位,因此本实验在设计寡核苷酸模板的时候,3′端均加上T7启动子引物 5′CCTGTCTC3′。在 RNA聚合酶结合到双链 DNA启动子后,聚合酶分开双链 DNA模板,并以 3′~5′链作为模板合成出互补的5′~3′RNA链,合成出的正反义 RNA链经杂交即可得双链RNA。在本研究中,以 AA开头针对 597 bp的 Annexin-1 cds区的靶序列共有 26个,其中 GC碱基含量在 50%以下的有 25个,对其进行 Blast分析,最终选择了 240~260(SⅠ),435~455(SⅡ)和 506~526(SⅢ)的 3段序列,其中 SⅠ siRNA靶序列的GC含量为 43.9%,SⅡ siRNA靶序列的 GC含量为 39.2%,SⅢsiRNA靶序列的 GC含量为 41.2%。结果表明 3段 siRNA序列都有抑制 Annexin-1基因表达的作用,但是SⅡ siRNA抑制作用要更强一些。通过研究 3对不同的靶向干扰序列和1对阴性对照序列,发现在mRNA水平 3对靶向序列均可抑制 Annexin-1的表达,在蛋白水平也获得较高的抑制效果。
本研究还提示转染 T24细胞后 siRNAⅠ、Ⅱ、Ⅲ组细胞对顺铂的 IC50值(μg/ml)分别下降了 3.8、4.4和 5.3倍,顺铂敏感性显著增加。通过干扰 Annexin-1基因,可以改变膀胱癌 T24细胞生存能力,对照组在不同的顺铂浓度下(20~80μg/ml)生存率分别为 94.54%、87.22%、58.39%和 42.83%,经 3个 siRNA干扰后生存率明显下降。膀胱癌 T24细胞是顺铂耐药的细胞〔8〕,siRNA干扰后对顺铂敏感性增加,从而为顺铂耐药的治疗提供了一个很好的思路。
1 Silistino-Souza R,Rodrigues-Lisoni FC,Cury PM,et al.Annexin-1:differential expression in tumor and mast cells in human larynx cancer〔J〕.Int J Cancer,2007;120(12):2582-9.
2 Ang EZ,Nguyen HT,Sim HL,et al.Annexin-1 regulates growth arrest induced by high levels of estrogen in MCF-7 breast cancer cells〔J〕.Mol Cancer Res,2009;7(2):266-74.
3 Baffa R,Letko J,Mc Clang C,et al.Molecular genetics of bladder cancer:targets for diagnosis and therapy〔J〕.J Exp Clin Cancer Res,2006;25(2):145-60.
4 Dalzell JJ,Mc Master S,Fleming CC,et al.Short interfering RNA-mediated gene silencing in Globodera pallida and Meloidogyne incognita infective stage juveniles〔J〕.Int J Parasitol,2010;40(1):91-100.
5 Pal-Bhadra M,Bhadra U,Birchler JA.RNAi related mechanisms affect both transcriptional and posttranscriptional transgene silencing in Drosophila〔J〕.Mol Cell,2002;9(2):315-27.
6 Elbashir SM,Harborth J,Lendeckel W,et al.Duplexes of 21-nucleotide RNAsmediate RNA interfernce in cultured mammalian cells〔J〕.Nature,2001;411(6836):494-8.
7 Berezhna SY,Supekova L,Supek F,et al.siRNA in human cells selectively localizes to target RNA sites〔J〕.Proc Natl Acad Sci U SA,2006;103(20):7682-7.
8 Konstantakou EG,Voutsinas GE,Karkoulis PK,et al.Human bladder cancer cells undergo cisplatin-induced apoptosis that is associated with p53-dependent and p 53-independent responses〔J〕.Int JOncol,2009;35(2):401-16.
