大白菜裂球性状的遗传与RAPD标记
2010-05-21郑晨光张鲁刚胥宇建张立志付文婷
牛 娜 郑晨光 张鲁刚 胥宇建 张立志 付文婷
(西北农林科技大学园艺学院,农业部西北园艺植物种质资源利用重点开放实验室,陕西 杨凌 712100)
大白菜〔Brassica campestris L.ssp.pekinensis(Lour)Olsson〕属十字花科芸薹属,原产于中国,栽培历史悠久,是芸薹属植物中具有较高经济价值、营养价值和研究价值的重要叶菜类作物之一。大白菜不仅在我国有着广泛的分布和栽培,而且在日本、朝鲜、韩国及南亚国家也有着广泛的栽培。大白菜结球性状直接关系到大白菜的产量和商品性,特别是早熟大白菜,一旦裂球,基本上失去商品价值。因此,提高大白菜的耐裂球性是大白菜育种的一个重要目标,但有关大白菜裂球方面的研究国内外尚未见报道。而在同属芸薹属的甘蓝作物上已有一些研究,主要集中在甘蓝裂球性的解剖学特征(王丽娟和秦智伟,1994)、性状的配合力和遗传力(庄木 等,2009)、内源激素的含量与裂球性的关系(曾爱松 等,2009)以及栽培因子对裂球性的影响(秦智伟 等,1994)等方面,尚未见分子标记方面的报道。
本试验以大白菜分离群体为试材,从建立大白菜裂球性状的分级标准入手,在分析大白菜裂球性状遗传规律的基础上,筛选与裂球基因连锁的分子标记,以期为耐裂球性大白菜品种选育和分子标记辅助育种提供参考。
1 材料与方法
1.1 材料
供试2套大白菜材料F1均有裂球现象,均由西北农林科技大学园艺学院大白菜课题组提供。
材料A:亲本分别为自交不亲和系06J31和自交系92S24,双亲正反交F1,F2为06J31×92S24的单株自交群体。06J31叶浅绿色,叶球叠抱,近圆球形,叶球高12 cm,球宽10.5 cm,生育期50~55 d;92S24叶绿色,叶球叠抱,倒卵形,生育期55~60d。F1叶球头球形,叠抱,结球极紧实,腋芽不发达,叶球高22.8 cm,球宽14.2 cm。
材料B:亲本分别为自交不亲和系06J31和05S50,双亲正反交F1,F2为06J31×05S50的单株自交群体。05S50株型小而紧凑,叶球高15 cm,球宽12 cm,外叶深绿,叶球叠抱,生育期50~55 d。F1叶球头球形,叠抱,结球极紧实,腋芽不发达,叶球高17 cm,球宽12 cm。
亲本材料结球松散,紧实度较低,正常情况下不表现裂球性。
1.2 方法
试验材料B、A分别于2008年8月5、10日大田播种育苗,育苗期间按常规管理(钮心恪,1984)。材料B于8月24日定植,材料A于9月2日定植。大田定植后,单株编号,叶球成熟期连续观察叶球变化。以材料A的F2群体为材料,进行RAPD分子标记研究。
1.2.1 F2群体裂球基准时期及最佳分析时期的确定 以F1群体的成熟期(80%植株叶球达到紧实时的天数)作为F2群体裂球记载的基准时期,在此基础上延迟生育期,每隔5 d观察1次裂球性状,至群体裂球率不再变化为止。分析群体裂球植株增加速率,统计裂球率。
1.2.2 裂球性状分级标准的确定及遗传规律分析 从裂球基准时期开始,连续观察大白菜叶球开裂变化情况,分析裂球变化的范围,确立了裂球性状分级标准。
零级:叶球无裂叶;一级:叶球有1~3片叶裂;二级:叶球有4~5片叶裂,且叶球裂缝深度/叶球纵经≤0.2;三级:0.2<叶球裂缝长度/叶球横径≤0.4,0.2<叶球裂缝面积/叶球表面积≤0.4,0.2<叶球裂缝深度/叶球纵径≤0.3;四级:0.4<叶球裂缝长度/叶球横径≤0.7,0.4<叶球裂缝面积/叶球表面积≤0.6,0.3<叶球裂缝深度/叶球纵径≤0.5;五级:叶球裂缝长度/叶球横径>0.7,叶球裂缝面积/叶球表面积>0.6,叶球裂缝深度/叶球纵径>0.5。
按照上述裂球性状分级标准完成田间调查后,参照孟德尔遗传规律进行初步分析,并进行适合性检验。数据采用DPS软件进行统计分析(唐启义和冯明光,2002)。
1.2.3 模板DNA提取 亲本、F1及F2单株的基因组DNA提取采用李永明和赵玉琪(2001)的方法,并参照王绮等(2006)方法,稍有改动。操作方法的改进点:取0.2 g幼嫩叶片装入1.5 mL离心管中,在盛有液氮的容器中预冷,取出时离心管中留一点儿液氮,然后迅速研磨成粉末。
1.2.4 RAPD扩增 采用25 μL反应体系,包括1×缓冲液,2.5 mmol·L-1MgCl2,0.2 mmol·L-1dNTPs,0.3 μmol·L-1引物,Taq DNA聚合酶1 U,模板DNA 20ng。反应条件为94 ℃预变性5 min,94 ℃变性1 min,36 ℃退火1 min,72 ℃延伸90s,45个循环,72 ℃延伸10min。最后在1.5 %琼脂糖凝胶上电泳分离,电泳缓冲液为1×TAE,电泳后先在EB中染色,然后在紫外灯下观察并照相。随机引物购自上海生工生物工程技术服务有限公司,Taq聚合酶购自宝生物(大连)有限公司,DNA Marker购自天根生化科技(北京)有限公司。
1.2.5 DNA混合池建立及引物筛选 根据田间调查结果,在F2分离群体中随机抽取零级(即不裂)和裂球程度较高的四至五级植株各10株,分别等量混合各单株DNA,构建裂球和不裂球DNA池,以两个DNA池为模板,采用BSA法(Ajisaka et al.,2001)进行RAPD引物筛选。为了使所得标记更加准确,不受环境等条件影响,选取早期裂球的植株。
1.2.6 遗传连锁分析 应用JoinMap 3.0软件计算遗传距离。
2 结果与分析
2.1 裂球性状统计基准时期和最佳分析时期的确定
材料 A、材料 B的 F2群体分别在定植后75 d和60d有80%叶球达到紧实,作为裂球情况统计的基准时期。对2套材料田间调查统计分析表明,大白菜裂球的多少受延长生育期天数的影响明显(图1),至大白菜植株几乎不再生长为止(一般为延长生育期 30~35 d),裂球株数随着生育期的延长而增多,这样很难分析裂球性状。为了明确其遗传规律,需要确定一个相对最佳时期来分析该性状。
从图1可以看出,2套材料F2群体单株的裂球发生规律基本一致,在延长生育期20d时可以看作裂球发生的临界期,此时裂球株数集中出现,形成裂球发生高峰,累计裂球株数约占总裂球株数的80%;此后裂球率日趋降低,至延长生育期30d以后,群体裂球株数几乎不再发生。因此,可将延长生育期20d作为裂球性状分析的最佳时期。
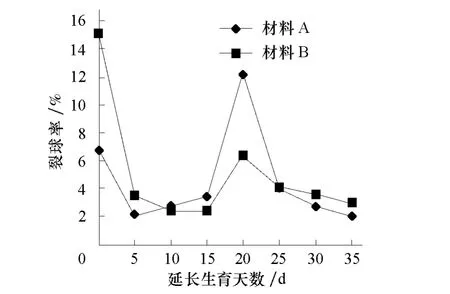
图1 F2群体裂球率的动态变化
2.2 正反交F1裂球期方差分析
在最佳分析时期,材料A正反交F1裂球率分别为26.3 %、31.5 %,材料B正反交F1裂球率分别为33.3 %、25.0%。
由表1可见,2套材料正反交组合间差异均不显著,这说明大白菜裂球性状是受核基因控制的,与细胞质遗传无关。
2.3 F2群体裂球性状遗传分析
分析2套材料在F2延长生育期20d时裂球性状的分离情况,发现2套材料均在延长生育期20d时不裂球株数与裂球株数的分离比例(不裂球∶裂球)接近3∶1,因此初步认为该性状可能受1对主效基因控制。
对2套材料F2群体裂球性进行χ2测验(表2),结果表明:延长生育期20d时,χ2值均<,符合3∶1的分离比例;由于材料A较材料B定植晚、生育期长,材料A延长生育期20~25 d和材料B延长生育期10~20d的不裂球与裂球的分离比例均符合3∶1的比例,2套材料所表现的这种时间差异,除了与材料本身(遗传因素)有关外,可能与生长期的环境条件尤其是温度也有很大的关系。

表1 F1裂球期的方差分析
上述分析表明大白菜裂球性状至少受1对主基因控制,同时还受到微效基因和环境条件的影响。
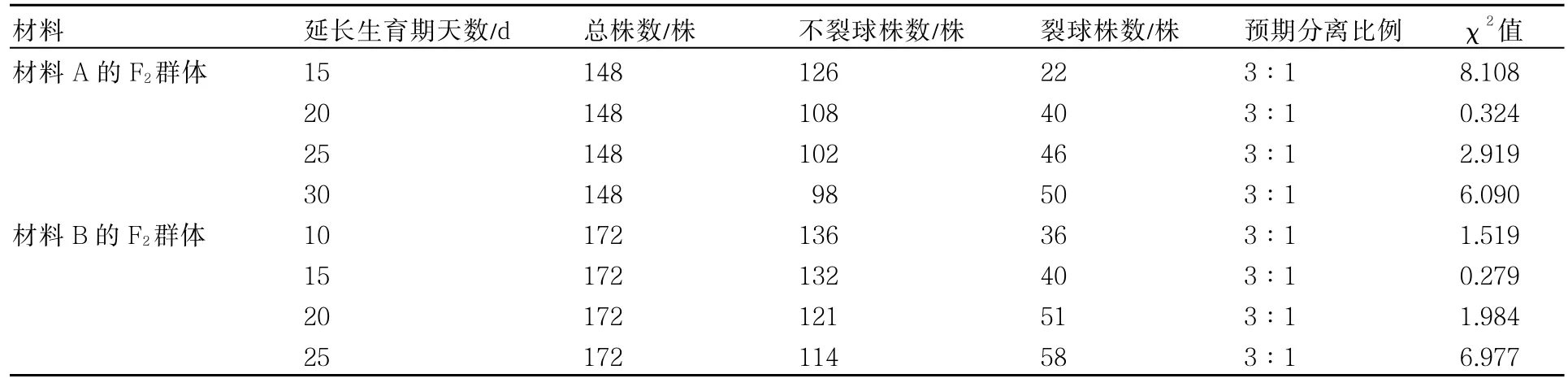
表2 大白菜F2裂球性状田间观察结果
2.4 RAPD引物筛选及标记分析
用本实验室保存的600个RAPD引物对裂球和不裂球DNA池进行扩增,不同引物扩增的条带数目为0~11条谱带,扩增片段大小为100~2000bp,有483个引物扩增出稳定而清晰的条带,其中只有87个引物在两池之间的扩增条带存在差异,表现出遗传多态性。
在87个引物中,表现出明显扩增多态性的引物有34个,占总引物的5.7 %。经3次以上重复筛选,有15个引物在两池间都表现出较好的多态性。经亲本、F1及F2验证,仅有1个引物S15在不裂球池内扩增出相同大小且清晰的特异条带(图2)。
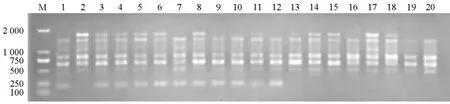
图2 引物S15对亲本、F1和F2的PCR扩增结果
对材料A的148个F2单株DNA用S15进行PCR扩增,有110个单株扩增出与亲本06J31相同的条带,38株未扩增出此特异条带,特异条带为222 bp。特异条带有/无(110/38)比率的卡方测验χ2值为 0.036(<=3.84),符合3∶1分离比例。认为该标记是与控制大白菜裂球性状的主效基因相关的RAPD标记。发生交换的植株有13株,交换率为8.784 %。应用JoinMap 3.0软件计算,该标记与裂球基因间的遗传距离为8.876 cM。
3 结论与讨论
3.1 关于裂球标准的划分
研究裂球性首先要进行裂球性状的判断与分级,大白菜裂球多以叶球开裂为主,但也有根裂现象的发生。实际生产中,大白菜叶球开裂的方式可以分为横裂、纵裂、混合裂和不规则裂等。因此,用单一指标难以准确评价单株裂球性,本试验采用叶球三维综合指标进行大白菜裂球性状的分级。该分级标准简单明了、且易于掌握,尤其方便田间调查使用,但由于试材有限,该分级标准比较粗,可在今后的研究中不断细化、完善。
本试验发现大白菜叶球开裂多发生在结球后期,裂球初始阶段发生裂球的植株少、叶球开裂程度低且变化缓慢,而在叶球成熟后期裂球植株逐渐增多、开裂速度加快。因此在分级标准划分时,裂球程度低的级别间级差小,以后逐渐增大,较高的裂球级别之间级差较大,使分级标准更加准确,为大白菜裂球程度的划分提供依据。
3.2 关于大白菜裂球性状的遗传规律
在传统的自交系选育中没有有意识地进行裂球性的选择,因此经济性状一致的自交系其控制裂球性的相关基因位点是随机的,如果双亲带有裂球基因,其配制的杂交一代就可能出现裂球植株,这不能简单的用显隐性解释。鉴于大白菜裂球现象的遗传复杂,本试验利用的是单株自交的F2群体,这可以消除亲本裂球性基因位点不纯合带来的遗传复杂性,取得相对准确的结果。
本试验通过对两套分离群体的调查分析认为大白菜裂球性状为质量-数量性状(莫惠栋,1993),耐裂球为显性,至少有1对主基因控制其性状的表现,同时还受到微效基因和环境条件的共同影响。这与秦智伟等(1994)认为甘蓝裂球性主要由遗传因素引起的研究结果相似。本试验使用的材料腋芽不发达,裂球的原因主要是内叶不断生长导致外叶断裂,其遗传规律反映了内叶生长和外叶韧度两方面的特性,具体以那一方为主尚不能确定,另外在实践中也常看到有些腋芽发达的大白菜品种容易裂球,这种情况与内叶生长关系不大,其与本试验材料可能属于不同的裂球类型,但尚无相关报道。
3.3 大白菜裂球性状最佳分析时期的确定
大白菜裂球性状为质量-数量性状,裂球率随着生育期延长而变化,因此确定适宜的统计时期十分重要,本试验以形成裂球发生高峰的时期作为该性状的最佳分析时期比较符合实际,分阶段统计分离比例表明此时的分离比例接近孟德尔遗传规律分离比例,也有利于主效基因的发现与分子标记研究,本试验是可行的。
3.4 大白菜裂球性状的分子标记
本试验以材料A的F2群体为材料,找到了与裂球基因紧密连锁的RAPD分子标记S15-222,其与裂球基因的遗传距离为8.876 cM,由于裂球性状也不同程度地受温度、水分等环境条件的影响,所以,因环境作用而表现为裂球的不裂球单株则不属于发生交换的范畴。因此,可以推测S15-222与裂球基因间的遗传距离≤8.876 cM,因此该标记可以有效地应用到大白菜裂球性状的分子标记辅助育种中,而且该标记是否适合材料B,还有待于验证。
李永明,赵玉琪.2001.实用分子生物学方法手册.北京:科学出版社:173-178.
莫惠栋.1993.质量一数量性状的遗传分析:I.遗传组成和主基因基因型鉴别.作物学报,19(1):1-6.
钮心恪.1984.大白菜抗霜霉病,病毒病原始材料的筛选及抗性遗传的研究.中国蔬菜,(4):28-32.
秦智伟,王丽娟,王超,许蕊仙.1994.甘蓝(Brassica oleracea var.capitata)裂球性研究Ⅰ.品种施肥及采收期对甘蓝裂球的影响.东北农业大学学报,25(4):344-346.
唐启义,冯明光.2002.实用统计分析及DPS数据处理系统.北京:科学出版社.
王丽娟,秦智伟.1994.春甘蓝裂球性的解剖特征//中国园艺学会.中国园艺学会首届青年学术讨论会论文集.北京:中国农业出版社:581-582.
王绮,张鲁刚,张战凤,惠麦侠,张明科.2006.白菜杂交种‘冠春’杂交率的RAPD分析.西北植物学报,26(4):677-682.
庄木,张扬勇,方智远,刘玉梅,杨丽梅,孙培田.2009.结球甘蓝耐裂球性状的配合力及遗传力研究.中国蔬菜,(2):12-15.
曾爱松,刘玉梅,方智远,杨丽梅,庄木,张扬勇,孙继峰,孙培田.2009.甘蓝结球过程中内源激素含量与裂球性的关系.中国蔬菜,(20):11-16.
Ajisaka H,Kuginuki Y,Yui S,Enomoto S,Hirai M.2001.Identification and mapping of a quantitative trait locus controlling extreme late bolting in Chinese cabbage(Brassica rapa L.ssp.pekinensis syn.campestris L.)using bulked segregant analysis.Euphytica,118(1):75-81.
