力竭运动后不同时相大鼠心肌细胞间粘附因子-1的变化
2010-05-12彭泽胄常芸
彭泽胄 常芸
国家体育总局体育科学研究所(北京 100061)
适宜负荷训练可引起心脏形态结构的可调节性、生理性重塑,有利于心脏功能的改善。然而,持续大强度运动和急性力竭运动等超负荷运动常引起运动性心肌微损伤[1]。在正常情况下,心肌细胞表面的细胞间粘附分子-1(intercellular adhesion molecules-1,ICAM-1)表达量极低,但炎性细胞因子(TNF、IL-1)[2,3]和脂多糖等可诱导心肌细胞中ICAM-1表达增强,促使炎性细胞的粘附和浸润,并释放有害物质,从而引起心肌细胞凋亡或坏死,但目前仍缺乏系统的有关过度训练引起的炎症反应导致心肌微损伤的研究。本实验采用一次力竭游泳和反复力竭游泳运动建立运动性心肌微损伤的实验动物模型,观察大鼠心肌ICAM-1蛋白在力竭运动后不同时相的表达变化,探讨其在运动性心肌微损伤及心肌功能异常发生中的调节作用,为心肌微损伤机制提供理论与实验依据。
1 材料与方法
1.1 实验对象与分组
健康雄性SD大鼠100只(维通利华公司提供),2月龄,体重219±8g。国家标准啮齿动物饲料喂养,自由饮食。饲养环境为室温18±2℃,光照时间12小时,相对湿度40%~55%。将大鼠随机分为一次力竭游泳运动组(n=40)、2周反复力竭游泳运动组(n=40)及相应对照组。
1.2 动物模型的建立
PVC特制泳池,规格为 1.6m×0.8m×1.2m,内壁光滑,水深约60cm,超过大鼠身长2倍,控制水温在30~33℃。安静对照组不运动。运动组大鼠进入动物房后首先适应3天,然后进行2天适应性游泳运动(20min/次)。一次性力竭组进行一次性游泳运动,游泳大鼠在尾部负重为体重3%,直至力竭,力竭标准参照Thomas DP报道[4],即连续沉入水中超过 10秒,捞出后置于平面不能完成翻正反射;在相对短时间内力竭的大鼠,将其捞出休息5分钟后,再继续进行游泳运动,使每次运动时间不少于2小时。反复力竭运动组每天1次力竭运动,游泳大鼠在尾部负重为体重3%。每周运动6天,共运动2周。每次运动时间不少于2小时。每次运动后,迅速用干毛巾擦干水分,电吹风吹干后,放回笼中休息。
1.3 实验方法
1.3.1 大鼠心肌取材
一次和反复力竭运动组分别于力竭运动后0、6、12、24小时取材。100只大鼠全部进入取材阶段。大鼠腹腔注射10%水合氯醛(1ml/100g体重)麻醉,开胸后迅速取出心脏,经灭菌生理盐水清洗后,滤纸吸干并称重,然后迅速垂直于心脏长轴切取心肌组织两块,其中近心尖心肌组织投入福尔马林液固定24小时后于石蜡切片机切取5μm厚的石蜡切片备用,近心底心肌组织置于冰冻切片样品托,滴加OCT包埋剂后置于液氮冷冻20秒后转入冰冻切片机恒温箱,切取7μm厚的冰冻切片置于-70℃超低温冰箱备用。
1.3.2 免疫荧光染色
主要试剂:羊多克隆ICAM-1抗体购于北京中杉生物技术有限公司;FITC标记兔抗山羊IgG购于北京中杉生物技术有限公司;德克萨斯红标记的麦芽凝集素购于美国Invitrogen公司。
由于ICAM-1在心肌细胞膜上表达,为了更好的定位,采用双荧光标记,用红色荧光标记细胞膜,绿色荧光标记ICAM-1,具体步骤如下:(1)将-70℃超低温冰箱保存的冰冻切片取出,室温复温30分钟后入4℃丙酮固定10分钟,PBS 漂洗 5分钟×3;(2)100μg/ml的德克萨斯红标记的麦芽凝集素(美国Invitrogen公司)室温孵育45min,经 PBS漂洗 5分钟×3;(3)10%正常兔血清封闭,室温孵育15分钟;(4)倾去血清,勿洗,滴加1∶50稀释的山羊抗大鼠ICAM-1(北京中杉金桥生物公司),4℃过夜,PBS漂洗 5分钟×3次;(5)滴加1∶50稀释的 FITC标记兔抗山羊IgG,37℃温箱孵育1小时,PBS漂洗5分钟×3次;(6)60%的缓冲甘油封片。
1.3.3 图像采集与分析
主要仪器:Leica 4000图像分析仪,单色激发光源,放大倍数为200倍,图像存为1024×1024像素类型。每张切片左室、室间隔、右室各随机选择6个视野,采用Leica Qwin图像分析系统对所拍图像荧光强度进行定量,荧光强度用积分灰度代表,参考阴性对照标本中荧光强度,灰度值在40~130之间为蛋白阳性表达。
1.3.4 统计学分析
所有数据均用SPSS15.0统计软件进行处理,结果用平均数±标准差表示,组间比较采用多因素方差分析,P<0.05为显著性水平,P<0.01为非常显著性水平。
2 结果
由图1可见,FITC标记蛋白阳性表达呈绿色荧光,基本位于德克萨斯红标记细胞膜(红色荧光)的位置,对照组荧光强度最弱,其他各组荧光染色面积及强度均有不同程度的升高。各运动组积分灰度值均不同程度高于对照组,其中以一次力竭即刻和反复力竭运动后6小时最为典型。

2.1 一次力竭运动后大鼠心肌细胞内ICAM-1蛋白表达的变化
如表1所示,一次性力竭游泳后,运动后即刻组大鼠心肌各部位ICAM-1快速表达,蛋白含量均显著高于对照组(P<0.01);运动后 6小时组ICAM-1蛋白含量达到峰值,并显著高于其他各时相组(P<0.01);运动后12小时组和24小时组心肌各部位ICAM-1蛋白表达呈下降趋势,但仍高于对照组(P<0.01)。

表1 一次力竭运动后心肌ICAM-1蛋白表达总灰度值变化情况
2.2 反复力竭运动后大鼠心肌细胞内ICAM-1蛋白表达的变化
如表2所示,反复性力竭游泳后,大鼠心肌各部位ICAM-1蛋白含量在力竭后即刻、6小时组和 12小时组与对照组相比均有显著性差异(P<0.01);左心室ICAM-1蛋白含量在力竭后24小时高于对照组(P<0.05),室间隔和右心室ICAM-1蛋白含量在力竭后24小时与对照组相比无显著性差异(P>0.05);反复力竭后6小时心肌各部位ICAM-1蛋白含量显著高于各时相组(P<0.01)。
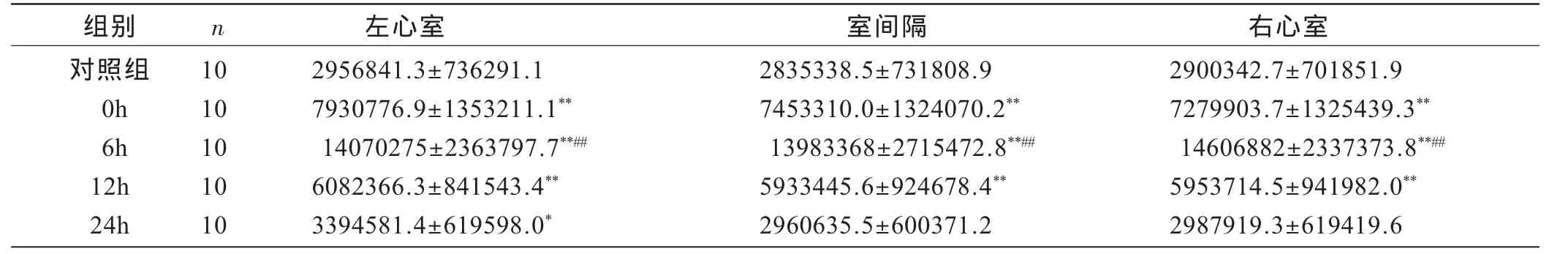
表2 反复力竭运动后ICAM-1蛋白表达总灰度值变化情况
2.3 两种不同力竭运动后心肌各部位ICAM-1蛋白表达的变化
两种不同力竭运动后各部位ICAM-1蛋白表达趋势基本一致,但表达量存在差异。大鼠心肌各部位ICAM-1含量反复力竭运动后即刻组显著高于一次力竭后即刻组(P<0.01);力竭运动后6小时蛋白表达均达到峰值,在左心室和室间隔处,两种力竭运动之间蛋白含量无显著性差异(P>0.05),而反复力竭后 6小时组右心室 ICAM-1含量显著高于一次力竭运动组(P<0.01);力竭运动后12小时组一次力竭运动后大鼠心肌各部位ICAM-1含量均显著高于反复力竭组(P<0.01);力竭运动后24小时组一次力竭后左心室ICAM-1含量高于反复力竭组(P<0.05),室间隔和右心室ICAM-1含量显著高于反复力竭组(P<0.01)。
3 讨论
研究表明,心肌局部或全身性炎症反应不管在缺血再灌注后,还是其他因素引起的心肌损伤的发病过程中都起着重要作用。王岚峰[5]通过研究发现急性心肌梗死(AMI)后炎症指标明显升高,并且炎症指标越高,微循环灌注越严重。另外,占成业等[6]对高血压左室肥厚时ICAM-1在心肌中的表达研究发现,心肌ICAM-1 mRNA和蛋白表达水平及巨噬细胞浸润显著增加,心肌ICAM-1过度表达及其介导的炎性细胞浸润可能参与了高血压左室肥厚的发病过程,提示炎症反应参与心肌损伤的发病过程。而赵敬国[7]从运动引起心肌损伤的角度,对一次性力竭运动后不同时相心肌的光镜结构呈现出一系列的动态改变进行观察,发现运动后即刻部分心肌纤维变性、浊肿、毛细血管充血;3小时可见有少数炎症细胞浸润;24小时则可见心肌细胞点状坏死及炎细胞浸润灶。这提示一次性力竭运动后有炎症反应参与。但炎症反应是否参与运动性心肌微损伤及具体机制目前还未见报道。
ICAM-1属于粘附分子免疫球蛋白超家族类,在体内分布广泛,包括各种上皮细胞、血管内皮细胞、成纤维细胞、网状结构、单核巨噬细胞和淋巴细胞,心肌细胞也能表达ICAM-1。在多种炎性介质(如TNF-α、IL-1等)的刺激下,通过激活核因子KappaB(NF-KB)、活化蛋白-1(AP-1)、信号转导子和转录激活子(STAT)等,可促进ICAM-1分子的表达上调[8,9],并参与多种细胞信号传导,引起细胞骨架蛋白结构与功能的改变,参与调节机体的正常生长发育,参与调节免疫细胞的分化,促进细胞间粘附及细胞活化,参与细胞因子产生。近年来的研究表明,ICAM-1的异常表达与心肌损伤、心功能衰竭、心脏移植排斥等多种心血管疾病的发病及治疗有关[10]。
研究发现,ICAM-1通过与LFA-1结合参与炎症反应免疫细胞作用、微血管损害、血栓形成及缺血再灌注损伤等病理过程。吴弘等[11]通过实验观察缺氧/复氧对心肌细胞与PMN粘附效应的影响及ICAM-1和淋巴细胞功能相关抗原(LFA-1)在PMN介导的心肌细胞损伤中的作用发现,缺氧/复氧使心肌细胞与PMN粘附效应增加,心肌细胞损伤加重,ICAM-1和LFA-1参与这一过程;毛宗珍等[12]通过建立不同负荷大鼠运动模型,观察大鼠心肌超微结构及ICAM-1表达发现,一般训练组心肌呈与有氧代谢相适应的生理性肥大,心肌细胞ICAM-1表达为阴性;过度负荷组大鼠心肌呈现病理性改变,心肌细胞ICAM-1表达明显增多。
本实验结果显示,反复进行的持续大强度运动和急性力竭运动可对机体产生不利影响,导致运动性疲劳与损伤发生,尤其是运动性心肌微损伤,这种损伤在6小时的时相较严重,24小时后有所减轻,这些特点与临床上的缺血再灌注损伤相似。由于运动性缺氧复氧过程对心肌结构功能的影响与临床医学中的心肌缺血再灌注损伤症状十分相似,我们可将这种由于剧烈运动及运动后恢复期而引发心肌缺氧复氧过程,从而导致心肌结构功能的改变称之为运动性缺血再灌注损伤。在对力竭运动后不同时相大鼠心肌ICAM-1的观察中发现,两种不同力竭运动后6小时组ICAM-1含量表达出现最高值,我们认为这是缺血再灌注的直接表现。正常情况下,心肌ICAM-1很少表达,心肌缺血时促使心肌分泌促炎因子,激活了ICAM-1的表达,促进中性粒细胞与心肌细胞的粘附并发挥其毒害作用,直接导致了心肌损伤,这与Lee等[13]对心肌损伤机制的研究结果类似。另外,Polazzo[14]用ICAM-1基因被敲出的鼠进行心肌缺血再灌注实验发现,与正常鼠相比,ICAM-1基因阴性鼠心肌损伤程度明显减轻;ICAM-1基因阴性鼠重新转染ICAM-1基因,再进行缺血再灌注实验,发现转染ICAM-1基因鼠心肌损伤程度明显加重,用ICAM-1单克隆抗体可减轻缺血再灌注造成的心肌损害,说明ICAM-1对心肌缺血再灌注损伤的发生有重要意义。当发生炎症反应等病理过程时,血管内滚动的PMN通过其LFA-1与VEC上的ICAM-1结合,从而相互粘附。一旦粘附发生后即可通过多种机制损伤组织,如进一步吸附血小板及PMN造成毛细血管堵塞呈无血流状态,此时微循环中的PMN与心肌细胞只隔一层VEC,PMN可释放OFR、弹性蛋白酶等损伤介质直接扩散到心肌细胞产生损伤;PMN还可跨膜迁移到ICAM-1表达增加的心肌细胞内,直接释放溶细胞物质如穿孔素等造成心肌细胞坏死。
本研究发现,两种力竭运动后即刻,大鼠心肌ICAM-1含量明显上升,力竭后6小时达到最高值,这与缺血再灌注对心肌造成的损伤时相趋势一致。虽然运动性缺血再灌注损伤是在机体保护性抑制系统监控下发生的一个主动缺氧的过程,但是反复力竭运动能够引起过度疲劳,导致心肌超微结构严重紊乱、心律失常、心功能降低等情况,使结构功能出现不可逆的改变。在一次力竭后24小时组中,大鼠心肌各部位ICAM-1含量与对照组相比均有显著差异,说明在一次力竭24小时后,导致心脏结构的损伤和功能上的改变,在短时间内无法完全恢复;而反复力竭24小时组心脏左心室ICAM-1含量显著高于对照组,右心室和室间隔ICAM-1含量与对照组无显著性差异,说明两种情况,一是反复力竭运动后心肌炎症反应基本恢复到正常水平;二是心肌细胞坏死导致产生ICAM-1减少,所以蛋白含量与对照组无显著性差异。
在对两种力竭运动各时相的观察中,在一次力竭运动后即刻、6小时和12小时左心室ICAM-1含量要高于室间隔和右心室,推测在力竭运动过程中,心脏以提高心率来维持心输出量,左心室在快速收缩泵血的过程中,容易造成自身缺血缺氧的状态,从而引起炎症反应的速度和程度要快于室间隔和右心室;一次力竭后24小时左心室ICAM-1含量低于室间隔和右心室,这可能与左心室在力竭运动后恢复时供血充足有关,也有可能是左心室心肌坏死程度要高于室间隔和右心室;而反复力竭后即刻、12小时和24小时左心室ICAM-1含量高于室间隔和右心室,由于反复力竭对心肌造成的损害远远大于一次性力竭,在恢复过程中,复氧复血反而加重了心肌损伤,这与缺血再灌注造成心肌损伤的结果类似。另外,在对大鼠心肌各部位进行一次性力竭和反复性力竭比较发现,反复性力竭后即刻组大鼠心肌各部位ICAM-1蛋白含量均显著高于一次性力竭后即刻组,说明反复力竭运动对大鼠心肌造成的缺血缺氧程度要大于一次性力竭运动,促使大鼠心肌快速表达;此外,两种力竭运动之间比较,一次性力竭运动后12小时组和24小时组大鼠心肌各部位ICAM-1含量要显著高于反复力竭运动,推测反复力竭后大鼠心肌对缺血缺氧耐受性提高,恢复程度要好于一次力竭运动;另外一种可能是反复力竭不仅仅只引起心肌发生微损伤,甚至引起心肌坏死,在ICAM-1介导炎症反应的过程中过度耗竭,或者是坏死的心肌细胞产生的ICAM-1减少等缘故。
4 总结
4.1 一次力竭运动和反复力竭运动后心肌各部位出现ICAM-1表达上升,心肌呈现缺血缺氧性改变,其改变可能与急性大强度运动引起炎症反应造成心肌损伤有关。
4.2 力竭运动后心肌在缺血缺氧的条件下刺激ICAM-1的快速表达,可诱导细胞因子的激活,介导心肌细胞的粘附和浸润等炎症反应,构成运动性心肌微损伤的发生机制之一。
4.3 力竭运动后24小时ICAM-1不同程度下降,也提示急性大强度运动后心肌ICAM-1改变可能具有可恢复性,有别于病理性改变。
[1]常芸.运动心脏理论与实践.第1版.北京:人民体育出版社,2008.106-132.
[2]Park S,Chang YH,Cho YJ,et al.Cytokine-regulated expression of vascular cell adhesion molecule-1inhuman glomerular endothelial cells.Transplant Proc,1998,30(5):2395-2397.
[3]Andersen CB,Blaehr H,et al.Expression of the intercellular adhesion molecule-1(ICAM-1)in human renal allograft sand cultured human tubular cells.Nephrol Dial,1992,Transplant(7):147-154.
[4]Thomas DP,Marshall KI.Effects repeated exhaustive exercise on myocardial subcellular membrane structures.Int J Sports Med,1988,9(4):257-260.
[5]王岚峰,修春红,李丹,等.急性心肌梗死后心肌微循环障碍与炎症反应的关系.中华超声影像学杂志,2008,17(8):106.
[6]占成业,潘李莉,等.高血压左室肥厚时细胞间黏附分子-1在心肌中的表达及其作用.中华急诊医学杂志,2005,14(1):47-50.
[7]赵敬国.大鼠力竭性游泳过程中心率变异性的动态观察.山东体育科技,2000,22(4):27-35.
[8]Kukielka GL,Hawkins HK,Michael L,et al.Regulation of intercellular adhesion molecule-1(ICAM-1) in ischemican dreperfused canine myocardium.J Clin Invest,1993,92(3):1504-16.
[9]Fan H,Sun B,Gu Q,et al.Oxygen radical strigge reactivation of NF-kappaB and AP-1 and up-regulation of ICAM-1 in reperfused canine heart.Am J Physiol Heart Circ Physiol,2002,282(5):1778-1786.
[10]Davani EY,Dorscheid DR,Lee CH,et al.Novel regulatory mechanism of cardiomyocyte contractility involving ICAM-1 and the cytoskeleton.Am J Physiol Heart Circ Physiol,2004,287(3):1013-1022.
[11]吴弘,沈茜,郎希龙,等.细胞粘附分子在中性粒细胞介导的缺氧/复氧心肌细胞损伤中的作用.中国应用生理学杂志,2001,17(3):27-229.
[12]毛宗珍,李恩荆,葛新发.运动对大鼠心肌ICAM-1表达与超微结构的影响.武汉体育学院院报,2004,18(6):64-67.
[13]Lee JR,Han JJ,Seo JW.Correlation between ICAM-1 and functional recovery of piglet myocardium with leukovytedepleted reperfusion.Ann Thorac Surg,2000,70:1531-35.
[14]Palazzo AJ,Jones SP,Girod WG.Myocardial ischemia-reperfusion injury in CD-18 and ICAM-1-deficient mice.Am J Physiol,1998,275(6pt2):2300-2307.
