西南桦遗传多样性研究
2010-05-09黄寿先桂仁意
董 蔚,黄寿先,陈 荣,桂仁意
(1. 浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300;2. 广西大学 林学院,广西 南宁 530004)
西南桦遗传多样性研究
董 蔚1,黄寿先2,陈 荣1,桂仁意1
(1. 浙江农林大学 亚热带森林培育国家重点实验室培育基地,浙江 临安 311300;2. 广西大学 林学院,广西 南宁 530004)
摘要:采用改良CATB法对采自云南大青山、平果等9个天然分布区的150个西南桦(Betula alnoides)样品进行DNA提取,获得了较高质量的DNA,然后利用AFLP分子标记技术,建立了适用于西南桦的AFLP反应体系;选择5对AFLP引物,对150个西南桦样品进行扩增,共检测出158个位点,其中多态性位点115个,占72.8%,每对引物获得20~55个位点,多态性比例为50.0%~84.0%。通过对西南桦的遗传多样性分析,结果表明:在种间个体水平上,多态性位点百分率为76.58%~93.04%,平均为83.8%;在群居水平上,多态性位点百分率为72.80%。可见西南桦天然居群内的变异大于居群间的变异;由Nei’s遗传距离分析得出,西南桦大部分居群间还是存在一定的遗传距离,其中最大在东兰和靖南之间,为0.020 1。
关键词:西南桦;遗传多样性;分子标记;AFLP;多态性
西南桦(Betula alnoides)为桦木科桦木属在北半球分布最南的一个种,集中分布在云南、广西和贵州南部地区,该分布区与越南、老挝和缅甸的分布区连成一片,形成西南桦中心分布区,垂直海拔200~2 800 m[1~2]。西南桦为强阳性树种,喜光,不耐荫蔽,适生于酸性土壤,具一定的耐贫瘠能力。西南桦生长迅速,是继桉树后又一优良速生珍贵树种,与阴性或中性偏荫树种红椎、木荷、樟树等混交成林,不仅生长好而且林分结构好,防护性能强,生态效果好[3~5]。
西南桦生态、经济价值高,属旱季落叶树种,保持水土能力强;林内VA菌等微生物可改良土壤,为改造热带、亚热带地区生态公益林的理想树种。西南桦树干通直圆满,木材密度适中、纹理优美、不翘不裂极易于加工,被广泛应用于高级建筑装饰及高档家具制作。在广西百色,西南桦原木山场价便高达1 700元/m3。西南桦树汁营养丰富,可开发桦树汁作为饮料;树皮鞣质含量为7.0%~11.6%,可用于提制栲胶[3~4]。
1 材料与方法
1.1 材料
从云南大青山、平果等9个西南桦天然分布区采集样本,每个样本间隔50 m以上,挑选新鲜无病虫害完整叶片放入封口袋中,加变性硅胶干燥后带回实验室常温保存[6~7]。共采集9个居群150个单株(表1)。
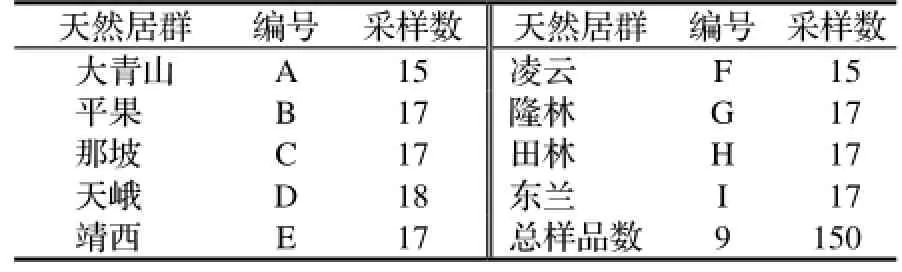
表1 西南桦采样情况Table 1 Sampling ofB. alnoides
1.2 方法
1.2.1 模板制备 参照李志真等[8]的改良CTAB法抽提基因组DNA,使用紫外分光光度计和0.8%琼脂糖凝胶电泳检测,并稀释到100 ng/μL备用。
1.2.2 AFLP分析 参照Vos等[9]的方法进行AFLP试验。基因组DNA用EcoRI和MseI双酶后,进行相应的接头连接,使用带一个选择性碱基的引物进行预扩,预扩产物稀释20倍作为选扩模板。经引物筛选,以扩增条带数目适中、多态位点比例高的5对引物组合(表2)进行选扩,并用6%聚丙烯酰胺凝胶进行电泳并银染显带。

表2 引物组合选择及序列Table 2 Selection of primer combination and the sequence
1.3 数据分析
根据电泳结果,读取100~800 bp中清晰的条带,将每个条带视为一个位点,有带记为“1”,无带计为“0”,建立二元数据矩阵,进行统计和计算。用AFLP_SURV1.0分析软件对统计结果进行分析,根据Nei等[10]计算遗传相似系数和遗传距离。
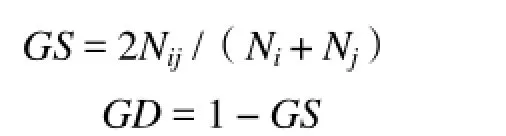
式中,GS为遗传相似系数,Nij为个体i和j的共有位点,Ni+Nj为总位点数,GD为遗传距离,当GD为1时表明遗传距离最远,当GD为0时表明遗传关系最近。
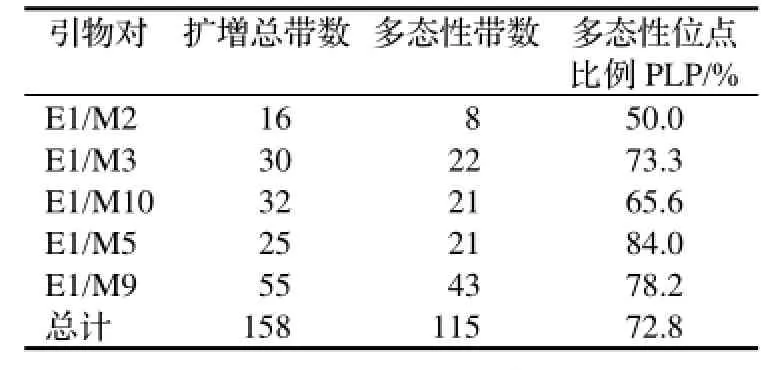
表3 扩增结果统计Table 3 Statistics of AFLP amplified
2 结果与分析
2.1 AFLP扩增结果
选用的5对引物组合共扩增出158条清晰有效条带,每对引物的扩增信息见表3。根据这158条AFLP标记条带进行西南桦的遗传多样性分析,各居群间多态性位点数、多态位点百分率以及基因多样性见表4。
从表4可以看出,云南西南桦各居群内的多态位点百分率为76.58%~93.04%,F居群的多态位点百分率最低,为76.58%,I居群的多态位点百分率最高,为93.04%,9个居群多态位点百分率平均为83.8%。根据多态位点百分率大小对9个西南桦居群进行排序为:I > G > C > A > B > D > E > H > F。

表4 西南桦群居遗传多样性Table 4 Genetic diversity of communities ofB. alnoides
Nei’s基因多样性分析结果则在0.209 8~0.284 0,F居群最低为0.209 8,I居群最高为0.284 0。9个居群Nei’s基因多样性分析结果由大到小排序为:I > C > G > A > B > D > H > E > F。

表5 Nei’s遗传距离Table 5 Nei’s genetic distance
在群居水平上,多态性位点共115个,占72.80%。由此可见,西南桦在群居间的遗传多样性低于居群内的遗传多样性,具一定的地域遗传变异。Nei’s基因多样性分析结果为0.244 2,为居群内数值的平均值。
2.2 群体间遗传结构
9个西南桦天然群体遗传距离见表5,大青山与靖南、田林,平果与田林、东兰,那坡与靖南、田林,天峨与东兰,靖南与东兰之间的遗传距离均超过了0.010 0,其中靖南与东兰遗传距离最大,为0.020 1。
3 讨论
由于西南桦天然林分布区地理位置偏僻,分布面积广,样品采集困难[11];并且桦木属植物中富含多糖和多酚类物质,不利于DNA提取[8];同时,从分子水平对西南桦遗传多样性进行分析尚未见报道。因此本试验通过AFLP分子标记对广西区西南桦天然林进行遗传多样性分析,结果表明,9个天然群体多态位点百分率为76.58%~93.04%,平均值为84.05%,明显高于居群间水平,同时群体内的遗传多样性和基因多样性也高于群体间水平。说明西南桦遗传多样性丰富,且不同天然分布区之间遗传变异不大。
西南桦的居群内的遗传多样性较居群间水平丰富,可能有以下几个原因引起:①曾杰等[11~12]认为尽管由于不适宜生境的阻隔以及森林破碎化导致西南桦居群之间存在一定程度的隔离,但它们是风媒植物,相互之间仍然存在较强的基因流,从而减少了遗传漂变;②根据葛锦芳等[13]的推测,广西壮族自治区的西南桦是从云南沿着南盘江扩散而来,居群分化的历史较短,所以居群间分化较小;③西南桦的大规模采伐始于20世纪80年代,由于时间较短,所导致的生境破碎化对西南桦天然居群的遗传变异尚无显著影响;④本研究的调查范围仅限于广西壮族自治区,而且西南桦只在其西南部、西部和西北部有分布,其经度、纬度和海拔跨度较小。
西南桦遗传多样性的研究对开展其良种选育、种质资源保存以及天然林防护和经营具有重要意义[11~12]。因此,应加强现有群体的遗传多样性保护及基因交流,以防遗传多样性缩小或丧失。同时,挑选基因多样性高的居群作为遗传资源收集与保存。西南桦分布较为广泛,有效保护和合理经营遗传多样性丰富的居群有助于适应环境变化。
参考文献:
[1] 曾杰,郑海水,翁启杰. 我国西南桦的地理分布与适生条件[J]. 林业科学研究,1999,12(5):479-484.
[2] 曾杰. 我国西南桦研究的回顾与展望[J]. 林业科学研究,2006,19(3):379-384.
[3] 王卫斌. 西南桦生物学特性及发展前景[J]. 福建林业科技,2005,12(32):177-178.
[4] 陈朝飞,陈安. 西南桦的生物学生态学特性及其在我省的引种现状[J]. 广东林业,2003,19(1):15-17.
[5] 马克平. 试论生物多样性的概念[J]. 生物多样性,1993,1(1):20-22.
[6] 张国防,陈存及,邢建宏. 樟树干叶DNA提取方法的研究[J]. 江西农业大学学报,2006,28(1):111-11.
[7] Chase MW, Hills HH, Silica Gel. An Ideal Material for Field Preservation of Leaf Samples for DNA Studies[J]. Taxon,1991,40(5):215-220.
[8] 李志真,黄勇,谢一青,等. 不同保存方法对光皮桦总DNA提取效果的影响[J]. 分子植物育种,2006,4(1):131-134.
[9] Vos P,hogers M,Reijans T,et al. AFLP: a new technique for DNA fingerprinting[J]. Nucleic Acids Res,1995,23(21):4 407-4 414.
[10] Nei M,Li W H. Mathematical model for studying genetic variation in terms of restriction endonucleases[J]. Proc Natl Acad Sci USA,1979,76(10):5269-5273.
[11] 曾杰,王中仁,周世良,等. 广西西南桦天然居群遗传多样性的研究[J]. 植物生态学报,2003,27(1):66-72.
[12] 曾杰,郑海水,甘四明,等. 广西西南桦天然居群的表型变异[J]. 林业科学,2005,41(2):59-64.
[13] 葛锦芳. 云南桦木科植物的分类与地理分布[J]. 西南林学院学报,1985(1):1-9.
中图分类号:S792.159
文献标识码:A
文章编号:1001-3776(2010)04-0050-03
收稿日期:2010-03-10;修回日期:2010-05-05
基金项目:广西大学中南速生材繁育实验室开放研究课题“广西区西南桦遗传多样性研究”(KF(2004)-004)
作者简介:董蔚(1984-),男,浙江临安人,硕士,从事林木生物技术研究。
Research on Genetic Diversity of Betula alnoides
DONG Wei1,HUANG Shou-xian2,CHEN Rong1,GUI Ren-yi1
(1. State Key Laboratory Cultivation Base of Subtropical Silviculture, Zhejiang A & F University, Lin’an 311300, China; 2. Forestry College, Guangxi University, Nanning 530004, China)
Abstract:Extraction of DNA was conducted by improved CATB from 150 samples ofBetula alnoidesat 9 natural communities in Daqingshan, Pingguo, etc. Yunnan province. And AFLP reaction system was established by AFLP molecular marker. Five EcoRI/MseI primer combinations were selected and amplified 158 clear bands, among them 115 bands of polymorphic, taking 72.8%. Each primer enzyme mix amplified 20-55 bands, 50.0%-84.0% of them polymorphic bands. Analysis on genetic diversity showed that interspecific percentage of polymorphic loci was 76.58%-93.04%, average 84.05%, community percentage of polymorphic loci was 72.80%, indicating that more variation in community than that among communities. Analysis of Nei’s showed that there is significant genetic distance among most natural communities ofB. alnoides. The biggest is 0.0201 between Donglan and Jingnan.
Key words:Betula alnoides; genetic diversity; molecular marker; AFLP; polymophic
