湛江硇洲岛文昌鱼(Branchiostoma belcheri)的食性研究
2010-03-14颜云榕卢伙胜白秀娟赵浩波廖传翼
颜云榕,卢伙胜,白秀娟,赵浩波,廖传翼
(1.广东海洋大学水产学院,广东 湛江 524088; 2.中国科学院 海洋研究所,山东 青岛266071)
文昌鱼作为国家二级保护动物,在生物进化方面的研究中起着非常重要的作用。当代分子生物学研究证实,文昌鱼是现存的和脊椎动物亲缘关系最近的无脊椎动物[1]。对研究脊椎动物如何由无脊椎动物进化而来有着极其重要的研究价值,是研究脊椎动物起源和进化的模式动物[2],被誉为 “进化史中的标志性生物”(an iconic organism in the history of evolutionary biology)[3]。
我国学者在50多年的文昌鱼生物学研究中,对其发育、免疫和进化方面的研究取得很大进展[4],同时在文昌鱼分类学[5~8]、发育与生殖生物学[9,10]、人工繁殖[11~14]、资源调查与保护[15,16]和细胞系统[17]等领域的研究也有报道。国外学者对文昌鱼的研究在进化和发育学等方面也取得了较快的发展[18,19]。
长期以来,文昌鱼主要是作为进化方面的模式生物来研究,而对于其生态方面尤其是在海洋食物网中的作用研究甚少。Lin[20]利用扫描电镜观察文昌鱼胃含物,确认食物中的硅藻碎片; Nielsen等[21]研究发现文昌鱼通过其黏液状过滤器摄食亚微米级颗粒; Chen等[22]则利用脂肪酸和稳定同位素,开展文昌鱼食性研究,并指出文昌鱼在海洋食物网中具有重要作用。
广东海洋大学文昌鱼课题组对广东茂名大放鸡海域文昌鱼(Branchiostoma belcheriGray)形态[23]、分布[24~26]、生长[27]和耐受环境因子[28]等开展了研究,并对湛江硇洲岛海域文昌鱼基因[29]、生长与繁殖特性[27]等进行分析。本研究通过文昌鱼胃含物观察,研究其食性及变化特点,探讨文昌鱼在海洋食物网中的作用,为保护文昌鱼资源提供科学依据。
1 材料与方法
文昌鱼的样品均系2006年4~7月在湛江硇洲岛海区 (E110º31.538′~110º32.204′,N20º55.048′-20º55.620′)进行的文昌鱼资源调查中,由潜水员携0.075m2文昌鱼专用采集器[30]采集所得。在采集的泥沙中,用人工方法拣出文昌鱼,放进装有在同一地方采集的沙和海水的小桶里带回实验室,然后对文昌鱼进行速冻保存。前后共选择80个完整样品供定量分析。
食性分析时,先将文昌鱼消化道残留物取出,再用Leica三目显微镜接电脑,在显微镜下鉴定饵料生物的种类并统计数量,对胃中饵料生物的数量划分为几个等级:D表示优势(>80个); A 表示丰富(50~80个); C表示常见(16~49个); R表示稀少(<16个),同时拍照有代表性种类。尽量鉴定到最低分类单元。
2 结果与分析
2.1 文昌鱼的食物成分
湛江硇洲岛文昌鱼的消化道中可辨成分约有 34种,包括了硅藻、甲藻、蓝藻、线虫动物、原生动物、节肢动物(瓣鳃类幼虫、桡足类和枝角类)等 6个类别。食物成分可分为4类:植物性食物、动物性食物、有机碎屑和无机颗粒。从出现频率和出现数量来看,在植物性食物中,硅藻类(小环藻、圆筛藻、曲舟藻,根管藻)占绝对优势; 在动物性食物中,瓣鳃类幼虫占优势。各物种分类详见表1。
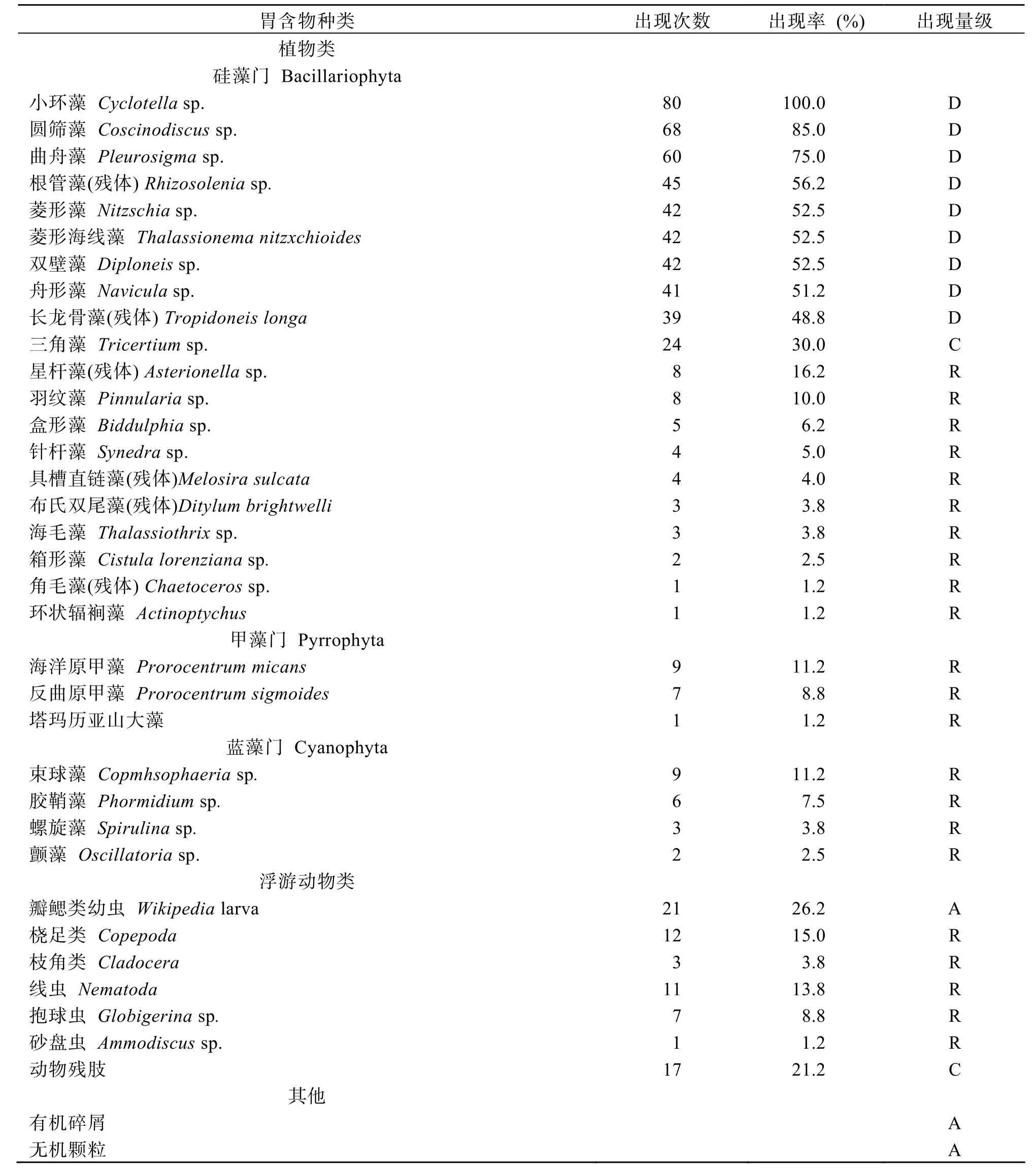
表1 湛江硇洲岛文昌鱼消化道中食物的生物种类Tab.1 Food composition in the alimentary canal of lancelet collected at the Naozhou Island of Zhanjiang
2.2 春夏季文昌鱼的食性比较分析
取文昌鱼胃中出现量级为常见(R)及以上级别饵料生物进行出现率的分析,发现春、夏的文昌鱼的摄食喜好不尽相同。春夏季度的饵料生物的出现率随着季节的变化有所变化。如春季文昌鱼消化道中根管藻(残体)Rhizosoleniasp.、菱形海线藻Thalassionema nitzxchioides、动物残肢等出现率明显高于夏季,而夏季文昌鱼消化道中双壁藻Diploneissp.、长龙骨藻(残体)Tropidoneislonga、瓣鳃类幼虫Wikipedialarva等出现率则明显高于春季(表2)。
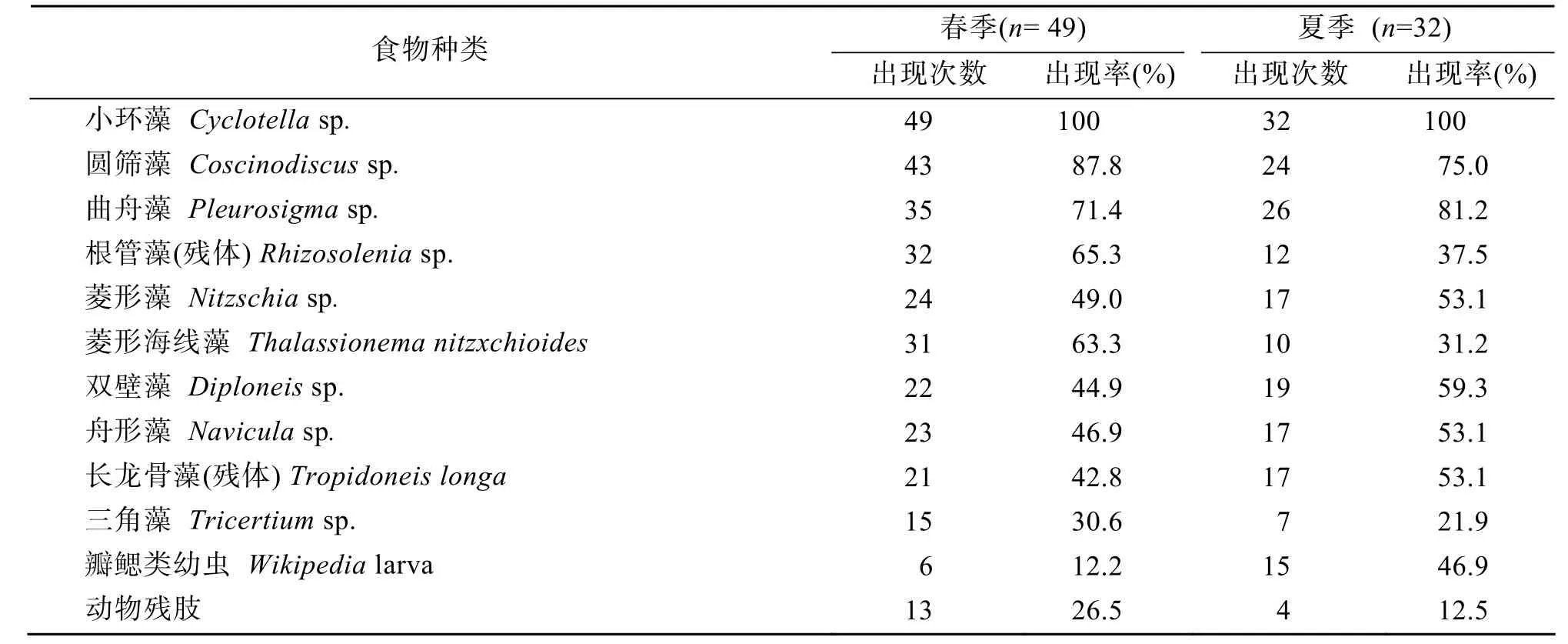
表2 湛江硇洲岛文昌鱼春夏季节的食性比较Tab.2 Comparison of the seasonal feeding habits of lancelets collected at the Naozhou Island of Zhanjiang
2.3 不同性别文昌鱼的食性比较分析
取文昌鱼胃中出现量级为常见(R)及以上级别饵料生物进行出现率的分析,发现不同性别的文昌鱼的摄食喜好比较相近。但雄性文昌鱼更喜食菱形藻Nitzschiasp.(♂:71.2% ♀:21.0%)、双壁藻Diploneissp.(♂:61.5% ♀:36.8%)和瓣鳃类幼虫Wikipedialarva(♂:38.5% ♀:5.3%)(表 3)。
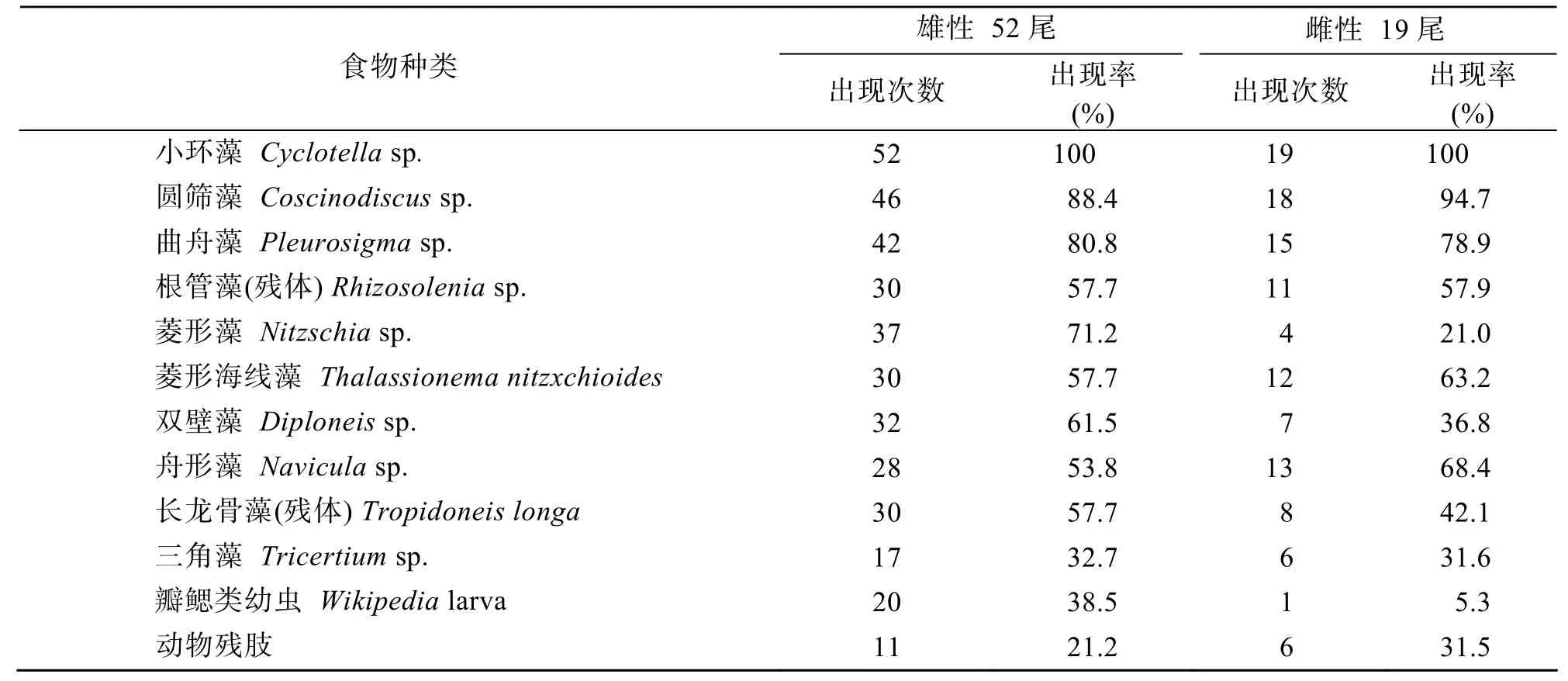
表3 湛江硇洲岛文昌鱼不同性别的食性比较Tab.3 Comparison of the feeding habits of different genders of lancelets collected at the Naozhou Island of Zhanjiang
2.4 不同体长组文昌鱼的食性分析
取文昌鱼胃中的出现量级为常见(R)及以上级别藻类及全部动物性食物进行出现率的分析,如表4所示。
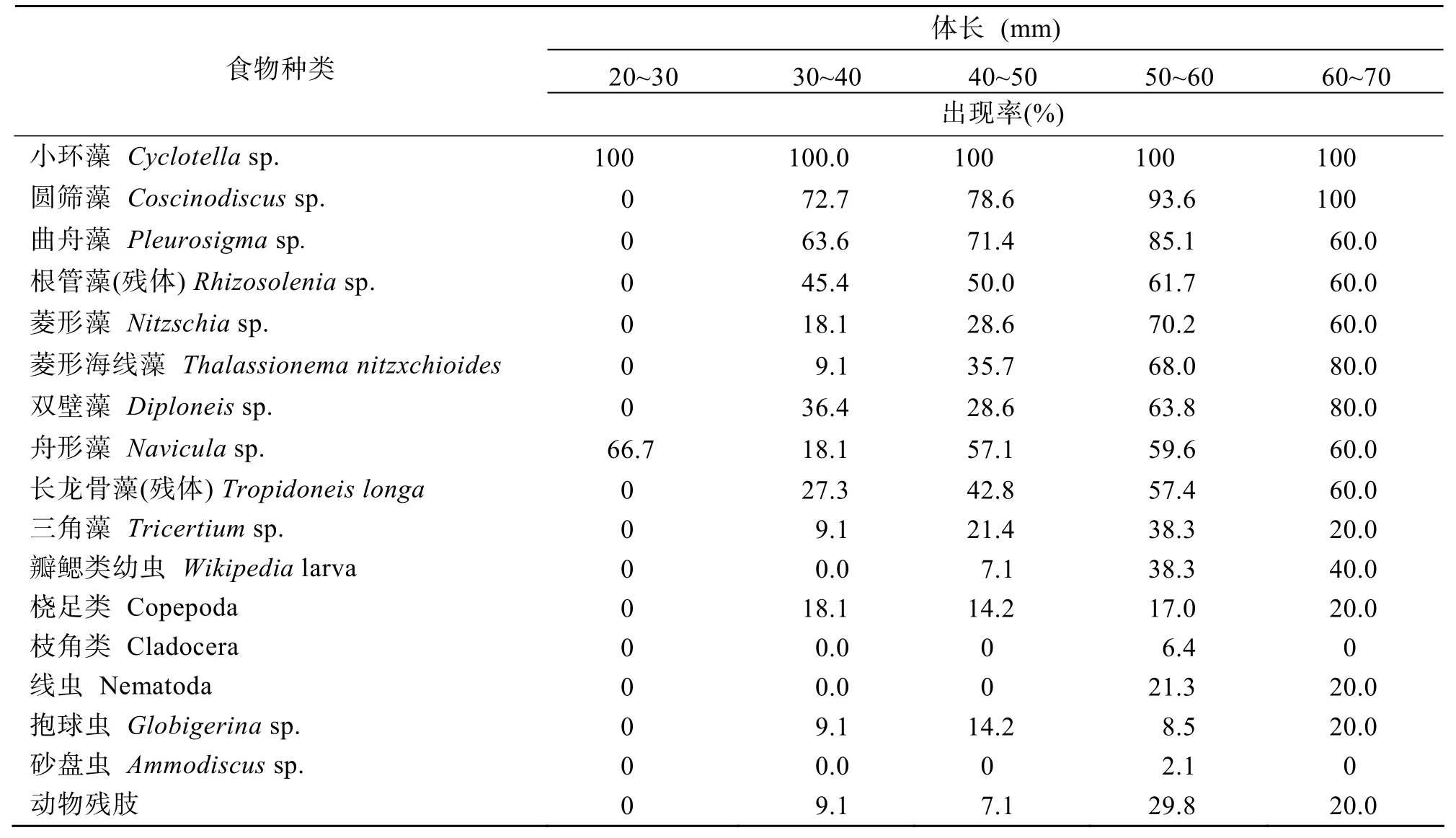
表4 湛江硇洲岛海域文昌鱼不同体长组的食性比较Tab.4 Comparison of the feeding habits of lancelets with various body lengths collected at the Naozhou Island of Zhanjiang
上述结果表明:在20~30 mm体长段中,胃含物中食物种类及食量均稀少,位于其他体长组之末。随着体长的增长,其胃含物中食物种类及食量基本呈增长趋势,而在50~60 mm与(60~70) mm体长段中,藻类及全部动物性食物出现率较为接近,可认为在这两个体长段中,文昌鱼食性的稳定性及对饵料的喜好程度比较一致。动物性食物开始出现在(30~40)mm的体长组中,并且最小个体为♂性体长32.8 mm,同时亦是观察中发现的最小成熟个体。枝角类(出现率 6.4%)与砂盘虫(出现率 2.1%)仅出现在(50~60)mm体长段中,但出现次数太少,可认为是偶然性食物。
3 讨论
张玺等[31]曾介绍过厦门文昌鱼体内占全部食料的3/4以上的是圆筛硅藻、舟形硅藻、小环藻和直链藻。除了硅藻外,还有些小型的浮游动物。在本研究中,硇洲岛海域文昌鱼胃含物中出现直链藻仅有 4次(出现率5.0%,出现量级为R),并非文昌鱼的主要食料,出现这种情况估计是采捕季节及两地海域文昌鱼栖息地饵料种类不同所致。
动物性食物出现在(30~40) mm的体长组中,并且最少个体为♂性体长32.8 mm,同时也是本研究中发现的最小成熟个体。对比蔡英亚[32]等观察到北部湾文昌鱼最小成熟个体体长为 29.5 mm,估计文昌鱼是到了性腺成熟后才开始产生相应的酶,以便消化桡足类和瓣鳃类幼虫等动物性食物。而文昌鱼酸性磷酸酶方面的酶[33,34]已有报道,在消化方面的酶尚需深入的研究。
有研究认为,文昌鱼为滤食性动物[21,35],本研究观察到幼小个体文昌鱼以植物性(硅藻类)饵料为主,成熟个体不仅摄食大量的硅藻类,而且也摄食少量的瓣鳃类和桡足类及有机碎屑等食物。但它们的优势食物种类均取决于其生存环境中的微小藻类及其他微小生物种类组成和数量的变动。同时,食物中生物的种类和组成随着季节的变化有所变化。该研究结果与茂名文昌鱼的食性分析相似[23]。研究结果表明,文昌鱼摄食藻类、微生物等而自身转化为其他动物的较高营养层次食物,在海洋食物网中具有一定的重要作用。
在食性研究中,较多地采用饵料生物的重量百分比来进行分析,但在文昌鱼的食性分析中,要获得食物的重量难度很大,并且食物在胃内处于消化状态,被消化的程度差别很大,在胃内能保持完整的生物个体极少。而饵料生物的出现频率较易获得,但由于饵料生物的个体大小差异较大,出现频率也很难全面地反映每种饵料生物的重要性。在最新的文昌鱼食性研究中已经采取了脂肪酸和稳定同位素进行定量分析[22],能够较准确地研究文昌鱼营养层次及食物来源,可以作为今后进一步研究的方向。
[1]张士璀,吴贤汉.从文昌鱼个体发生谈脊椎动物起源[J].海洋科学,1995,19(4):15-21.
[2]张士璀,袁金铎等.文昌鱼——研究脊柱动物起源和进化的模式动物[J].生命科学,2001,13(5):214-218.
[3]Gee H.Evolutionary biology-The amphioxus unleashed[J].Nature,2008,453(7198):999-1000.
[4]张士璀,郭斌,梁宇君.我国文昌鱼研究50年[J].生命科学,2008,20(1):64-68.
[5]王义权,方少华.文昌鱼分类学研究及展望[J].动物学研究,2005,26(6):666-672.
[6]闫路娜,左惠凯,曹玉萍.文昌鱼秦皇岛、青岛和厦门地理种群形态特征的分化[J].动物学研究,2005,26(3):311-316.
[7]孙玉苗,祝茜.威海海域文昌鱼的形态特征与分类学地位探讨[J].海洋科学,2008,32(9):18-24.
[8]Chen Y,Cheung S G,Kong R Y,et al.Morphological and molecular comparisons of dominant amphioxus populations in the China Seas[J].Marine Biology,2007,153(2):189-198.
[9]吴贤汉.温度和盐度对青岛文昌鱼胚胎发育的影响[J].海洋科学,1998,22(4):66-68.
[10]毛炳宇,李艳等.碱性磷酸酶在文昌鱼胚胎和幼体中表达图式的研究[J].发育与生殖生物学报(英文版),2000,9(1):21-26.
[11]方永强,翁幼竹,戴燕玉等.厦门文昌鱼人工繁殖和幼虫发育及其变态的研究[J].海洋学报,2005,27(4):102-107.
[12]王义权,张秋金,吕小梅等.文昌鱼的实验室繁育及子二代获得[J].动物学研究,2006,27(6):631-634.
[13]周仁杰,方琦.厦门文昌鱼人工繁育批量生产技术研究[J].台湾海峡,2007,26(1):121-128.
[14]范宁宁,李萍,张士璀.文昌鱼在中国的人工繁育[J].鲁东大学学报:自然科学版,2008,24(2):168-171.
[15]马明辉,张志南,冯志权等.滦河口青岛文昌鱼分布与栖息地底质特征[J].海洋环境科学,2005,24(2):39-42.
[16]杨建威,任一平,徐宾铎.青岛自然保护区文昌鱼的资源调查与生长研究[J].水利渔业,2008,28(3):112-114.
[17]石松林,黄晓明,李祺福,等.文昌鱼鳃组织原代培养[J].海洋科学,2009,33(3):44-48.
[18]Holland N D,Venkatesh T V,Holland L Z,et al.Amphink2-tin,an amphioxus homeobox gene expressed in myocardial progenitors:insights into evolution of the vertebrate heart[J].Developmental Biology,2003,255(1):128-137.
[19]Holland L Z,Holland N D,Gilland E.Amphioxus and the evolution of head segmentation[J].Integrative and Comparative Biology,2008,48(5):630-646.
[20]Lin X J.Systematic and ecological studies of amphioxus from the coastal areas of Taiwan,Kinmen and Matsu[D].Chinese Taipei:Taiwan University,2001.
[21]Nielsen S E,Bone Q,Bond P,et al.On particle filtration by amphioxus (Branchiostoma lanceolatum)[J].Journal of the Marine Biological Association of the United Kindoms,2007,87(4):983-989.
[22]Chen Y,Cheung S G,Shin P K.The diet of amphioxus in subtropical Hong Kong as indicated by fatty acid and stable isotopic analyses[J].Journal of the Marine Biological Association of the United Kindoms,2008,88(7Sp.Iss.SI):1487-1491.
[23]陈文河,卢伙胜,冯波.茂名文昌鱼形态及生态特征的初步研究[J].大连水产学院学报,2008,23(2):110-115.
[24]冯波,卢伙胜,陈文河.广东茂名大放鸡海区文昌鱼分布与底质关系[J].热带海洋学报,2006,25(2):18-23.
[25]宣立强,卢伙胜,何秀玲,等.茂名大放鸡海域文昌鱼分布与种群特性研究[J].中国生态农业学报.2007,15(2):147-150.
[26]卢伙胜,沈春燕,冯波.茂名大放鸡海域文昌鱼、蛇尾分布及其与底质粒度特征的关系[J].中国海洋大学学报,2008,38(2):297-302.
[27]冯波,卢伙胜,廖传翼,等.硇洲岛海域文昌鱼生长与繁殖特性[J].生态学杂志,2008,27(8):1327-1331.
[28]白秀娟,卢伙胜,廖春风.文昌鱼对几种环境因子及污染物的耐受极限初步研究[J].广东海洋大学学报,2007,27(1):92-94.
[29]郭昱嵩,王中铎,刘楚吾,等.湛江海域文昌鱼(Branchiostoma)线粒体 Cyt b基因的全序列分析[J].广东海洋大学学报,2008,28(1):6-10.
[30]卢伙胜,陈春亮,颜云榕.文昌鱼专用采集器[P].中国,ZL200420045790.7.
[31]张玺,张凤瀛,吴宝铃,等.中国经济动物志[M].北京:科学出版社,1963.
[32]蔡英亚,刘志刚,张志强,等.北部湾的文昌鱼[J].热带海洋学报,1986,5(2):42-50.
[33]颜思旭,陈素丽,蔡红玉,等.文昌鱼碱性磷酸酶的动力学初步研究[J].厦门大学学报(自然科学版),1980,19(3):64-71.
[34]陈素丽,龚少静,曾伟章.文昌鱼酸性磷酸酶的分离提纯及其性质的初步研究[J].厦门大学学报(自然科学版),1985,24(1):84-90.
[35]金德祥.文昌鱼[M].福州:福建人民出版社,1957.
