国家珍稀濒危保护植物永瓣藤生存群落的区系多样性研究
2010-01-19谢国文李海生王发松郑毅胜蔡耿雄彭碧珠林媚珍
谢国文,李海生,王发松,郑毅胜,蔡耿雄,彭碧珠,林媚珍
(1. 广州大学 生命科学学院,广东 广州 510006;2.广东第二师范学院 生物系,广东 广州 510303;3.湖北民族学院 化学与环境工程学院, 湖北 恩施 445000;4.广州大学 地理科学学院,广东 广州 510006)
植物区系多样性作为生物多样性表征之一,可从多方面反映特定区域植物多样性的特征.确定特定区域或群落的植物区系成分、自然环境性质,有利于揭示植物区系地理分布格局与环境特别是环境变迁的关系,对建立植物的自然发生分类系统,认识古地理的变迁或区域地理的自然历史提供有价值的资料或依据,以及更好地指导区域植物资源的开发利用和保护具有重要意义.近10余年来,国内在植物区系系统[1-4]、研究方法[5-7]、具体地区的植物区系分析[8-24]等方面大批植物学工作者做了大量的工作,但从珍稀植物所在群落植物区系的性质与特征等方面的研究报道尚少见.本研究偿试初步探讨单个国家重点保护物种所生存群落的小尺度区系特征,以其抛砖引玉.
永瓣藤(Monimopetalumchinense)是卫矛科(Celastraceae)的一个中国特有单型属[25-27],由于它分类位置孤立,在研究卫矛科系统演化上有重要意义;而且资源稀少,分布特殊,受到人类活动的严重威胁;并具药用价值,被列为国家二级稀有濒危植物[28,29].本文依据野外标本采集、样地调查,对永瓣藤群落进行区系分析,研究植物区系成分及其多样性特征,为物种多样性保护以及永瓣藤的开发利用提供植物地理科学依据.

表1 调查样方的地理位置和群落类型
1 自然环境、群落特点和分布特征
1.1 自然环境
永瓣藤天然分布于我国的江西、湖北、安徽等省,是华东植物区系地理成分[30,31],分布范围为北纬28°30′~30°10′,东经114°05′~118°10′,其分布区气候属温暖湿润型,年均气温15~17.3℃,一月平均气温4~5℃,7月平均气温33~35℃,绝对最低气温-10℃,绝对最高气温40℃,≥10℃年活动积温为5 200~5 400℃,无霜期230~250 d,年均降水量1 700~2 000 mm,5~6月最多,冬季最少,空气相对湿度75%左右.分布区的土壤多为山地红黄壤和山地黄壤,pH值为5.6~6.5,有机质含量在2.9%~5.1%,成土母岩多为千枚岩,页岩,板岩或沙岩,但一般不存在于石灰岩和花岗岩等山地[31].
1.2 群落特点
永瓣藤大都缠绕于常绿阔叶林或落叶阔叶林的林木上,零星地分布在杉木林、马尾松林、毛竹林、杂灌林等四类不同生态类型的群落中.乔木层以杉木(Cunninghamialanceolata)、马尾松(Pinusmassoniana)、毛竹(Phyllostachyspubscens)、赤扬叶(Alniphyllumfortunei)、野鸭椿(Toxicodendronsuccedaneum)、甜槠栲(Castaneasclerophylla)、野漆树(Euscaphisjaponica)等树种为主;灌木层常见的树种一般有檵木(Loropetalumchinese)、油茶(Camelliaoleifera)、鹿角杜鹃(Rhododendronlatoucheae)、格药柃(Euryamuricata)、老鼠矢(Symplocosstellaris)、山胡椒(Linderaglauca)等;草本层如狗脊蕨(Woodwardiajaponica)、阔鳞鳞毛蕨(Dryopterischampioni)、阔叶山麦冬(Liriopemuscari)等;层外植物多为南五味子(Kadsuralongipedunculate)、爬山虎(Parthenocissustricuspidata)、大血藤(Sargentodoxacuneata)、南蛇藤(Celastrusorbiculatus)、鸡矢藤(Paederiascandens)、刺葡萄(Vitiscuneata)、三裂蛇葡萄(Ampelopsisdelavayana)、菝葜(Smilaxchina)等种类.永瓣藤是一种半常绿的半阴性的缠绕木质藤本,不具备直立乔木或灌木的种间竞争优势,在森林群落中为伴生物种,处于从属地位.
1.3 分布特征
永瓣藤一般在土层较深、腐殖质含量丰富、排水良好的酸性土壤环境生长最好,在湿润的沟谷、土层浅薄、岩石裸露的地方也能适应,但在长期积水之地或干旱的山嵴未见分布.其分布区域狭窄,零星分布于赣西北及鄂东南的九岭、幕阜山林区(西部亚区)和赣东北与皖南交界的山区(东部亚区),因鄱阳湖水域使其呈东西间断分布[32].本研究共调查了11个群落样地,其中有8个群落样地是在江西省内,包括还有3个群落样地分别是在安徽省的祁门县牯牛降风景区观音潭、安徽省的休宁县左龙和湖北省的通山县沙店乡盘龙山(见表1),原有记载的贵池、贵溪、宜丰、万载等县现未见有其种群踪迹,很可能已绝灭,说明永瓣藤的分布区正在逐渐缩小.调查还发现,永瓣藤种群数量、规模与林分郁闭度有一定的相关性.永瓣藤在森林群落中为伴生成分,林分郁闭度过大或过小都不利于其生存.郁闭度达0.8或以上的天然林中就几乎找不到它的踪迹,即使偶尔存在,也仅是小种群或较小种群,年龄结构间断,为明显的衰退种群.林分郁闭度过小(≤0.3),人为干扰过度,也不适应永瓣藤的定居和散布.在垂直分布上,永瓣藤记载分布的海拔范围为130~1 000 m.经考察论证,其海拔分布下限为100 m,上限为1 000 m,以海拔400~700 m之间的山谷、沟旁、路边、疏林和林缘、林窗等生境条件较好的地段较多见.在海拔300 m以下的红壤环境中,缠绕于保存较好的杂灌林或天然的针阔混交林内.
2 研究方法
2.1 野外调查
野外调查按生态学常规的群落样方法对永瓣藤整个分布区的分布点(县)进行了群落调查,并在每个分布点根据生境选择有代表性的地段设置样方,每个样方面积为10 m×10 m,调查、采集与鉴定每个样方内的维管植物.
2.2 编目与统计方法
植物分类编目:蕨类植物按秦仁昌1978年的中国蕨类植物分类系统;裸子植物按郑万钧《中国植物志》第7卷的分类系统;被子植物按恩格勒分类系统;并按吴征镒等[1-3]的分布区类型划分科、属、种分布区类型.

表2 永瓣藤植物群落科内属的组成
3 结果与分析
3.1 区系组成的多样性
3.1.1 科属种统计 经过对永瓣藤所在区域的十多个群落植物区系调查、采集和鉴定,发现共有维管植物80科138属182种,其中蕨类植物7科、7属、7种:芒萁(水龙骨目里白科Gleicheniaceae芒萁属Dicranopteris)、海金沙(海金沙科Lygodiaceae海金沙属Lygodium)、鳞毛蕨(鳞毛蕨科Dryopteridaceae鳞毛蕨属Dryopteris)、狗脊(乌毛蕨科Blechnaceae狗脊蕨属Woodwardia)、凤丫蕨(裸子蕨科Hemionitidaceae凤丫蕨属Coniogramme)、中华金星蕨(金星蕨科Thelypteridaceae金星蕨属Parathelypteris)、紫萁(紫萁科Osmundaceae紫萁属Osmunda),其它为种子植物,有175种,分别隶属于131属、73科.种子植物中有裸子植物3科、3属、3种,被子植物70科、128属、172种.
3.1.2 各科所含属数情况 在这80科植物中,各科所含属数差异不是很大,含5~10属的有6科,占总科数的7.5%,属数36个,占总属数的26.09%,包括蔷薇科(Rosaceae)(8属,下同)、蝶形花科(Papilionaceae)(5)、茜草科(Rubiaceae)(6)、菊科(Compositae)(5)、山茶科(Theaceae)(5)、樟科(Lauraceae)(7).含2~4个属的科有19科,占总科数的23.75%,属数47个,占总属数的34.06%.只含1个属的科有55科,占总科数的68.75%,属数55个,占总属数的39.85%(见表2).由此可见,属在科内的分布,集中于大科的现象不突出,大部分的种分布在含5属以下的科内.
3.1.3 各科所含种数情况 在80科植物中,含10种或10种以上的科有蔷薇科(Rosaceae)(14种,下同)、樟科(Lauraceae)(13)、山茶科(Theaceae)(11)、壳斗科(Fagaceae)(10)、占全部维管植物科的5%,种的26.37%.含5~9种的科还有茜草科(Rubiaceae)(6)、菊科(Compositae)(6)、蝶形花科(Papilionaceae)(5)、禾本科(Poaceae)(5)、葡萄科(Vitaceae)(6)5科,上述9科合计48属,76种,占全部维管植物科的11.25%,属的34.78%,种的41.76%.此外,含2~4种的科有21科,40属,56种,占全部维管植物科的26.25%,属的28.99%,种的30.77%.只含1个种的科有50科,50属,50科,占全部维管植物科的62.5%,属的36.23%,科的27.47%,由此可见,植物集中于几个大科的现象比较明显,优势科在区系组成中占有重要地位(见表3).
3.1.4 各属所含种数情况 在维管植物的138属中,含5~10种属是栎属(Lithocarpus)(5种,下同),悬钩子属(Rubus)(6),占总属数的1.45%,占总科数的6.04%.含2~4种的属有24属,总计为59科,占总属数的17.39%,占总科数的32.42%.只含1个种的属有112个,占总属数的81.16%,占总科数的61.54%(见表4).由此可见,种在属内的分布,集中于大属的现象不突出,大部分的种分布在仅含1个种的属内.

表3 永瓣藤植物群落科内种的组成

表4 永瓣藤植物群落属内种的组成
3.2 生态类型多样性
永瓣藤的群落分布多为常绿阔叶林或落叶阔叶林的林木,以杉木林、马尾林、毛竹林、杂灌林等四类不同生态类型的群落为主,灌木植物发达.
永瓣藤是一种半常绿的半阴性的缠绕木质藤本,不具备直立乔木或灌木的种间竞争优势,在森林群落中为伴生物种,处于从属地位.永瓣藤植物群落的区系主要是由灌木或半灌木植物组成,乔木,草本只有少数属,如黄连木属、三尖杉属、栎属、栲属、刚竹属等,而灌木与半灌木植物却占到80%以上,占绝对优势地位.灌木植物种类较多的科有蔷薇科(14种,下同)、樟科(13)、山茶科(11)、壳斗科(10),占全部维管植物种的26.37%.
3.3 种子植物科的分析
根据中国种子植物科的分布区类型系统[2,3],可将永瓣藤植物群落区系分为11种分布区类型,分别阐述如下(见表5):

表5 永瓣藤生存群落植物区系种子植物科的分布区类型
3.3.1 世界分布 该类型有23科,以温带分布的草本植物为主,其中含有较多的属数和种数的有茜草科(Rubiaceae)、蔷薇科(Rosaceae)、菊科(Compositae)、蝶形花科(Papilionaceae)、禾本科(Poaceae)、其它的还有桑科(Moraceae)、鼠李科(Rhamnaceae)、玄参科(Scrophulariaceae)、榆科(Ulmanceae)、防己科(Menispermaceae)、椴树科(Tiliaceae)、十字花科(Cruciferae)、茄科(Solanaceae)、藜科(Chenopodiaceae)、蓼科(Polygonaceae)、莎草科(Cyperaceae)、败酱科(Valerianaceae)、兰科(Orchidaceae)、石竹科(Caryophyllaceae)、毛茛科(Ranunculaceae)、柳叶菜科(Onagraceae)、旋花科(Convolvulaceae)、木犀科(Oleaceae).
3.3.2 泛热带分布 泛热带分布的科有21科,其中含有较多的属数和种数的有山茶科(Theaceae)、樟科(Lauraceae)、葡萄科(Vitaceae)、漆树科(Anacardiaceae),其它的还有卫矛科(Celastraceae)、大戟科(Euphorbiaceae)、萝藦科(Asclepiadaceae)、鸭跖草科(Commelinaceae)、紫金牛科(Myrsinaceae)、山矾科(Symplocaceae)、冬青科(Aquifoliaceae)、芸香科(Rutaceae)、金粟兰科(Chloranthuaceae)、菝葜科(Smilacaceae)、荨麻科(Urticaeae)、爵床科(Acanthaceae)、柿科(Ebenaceae)、薯蓣科(Dioscoreaceae)、夹竹桃科(Apocynaceae)、醉鱼草科(Buddlejaceae)、大风子科(Flacourtiaceae).
3.3.3 热带亚洲至热带美洲间断分布 该类型有6科,分别是安息香科(Styracaceae)、马鞭草科(Verbenaceae)、省沽油科(Staphyleaceae)、杜英科(Elaeocarpaceaee)、五加科(Araliaceae)、木通科(Lardizabalaceae).每科所含的属数和种数较少,只有1~3属(种).
3.3.4 旧世界热带分布 该类型只有1科,为八角枫科(Alangiaceae),只含1属,1科.
3.3.5 热带亚洲至热带大洋州分布 该类型有3科,分别是马钱科(Loganiaceae)、虎皮楠科(Daphniphyllum)、姜科(Zingiberaceae),每科所含的属数和种数只有1属,1种.
3.3.6 热带亚洲至热带非洲 该类型只有1科,杜鹃花科(Ericaceae),含2个属,4种.
3.3.7 热带亚洲分布 该类型有3科,分别是清风藤科(Sabiaceae)、大血藤科(Sargentodoxaceae)、金丝桃(Hypericaceae),每科只含1属,1种.
3.3.8 北温带分布 该类型有10科,其中含有较多的属数和种数的有壳斗科(Fagaceae)、绣球科(Hydrangeaceae),其他的还有小檗科(Berberidaceae)、金缕梅科(Hamamelidaceae)、山茱萸科(Cornaceae)、百合科(Liliaceae)、胡桃科(Juglandaceae)、杉科(Taxodiaceae)、松科(Pinaceae)、忍冬科(Caprifoliaceae).
3.3.9 东亚至北美间断分布 该分布区有2科,分别是木兰科(Magnoliaceae)、五味子科(Schizandraceae),每科只含1属,1种.
3.3.10 东亚分布 该分布区有2科,分别是猕猴桃科(Actinidiaceae),三尖杉科(Cephalotaxaceae),每科只含1属,1种.
3.3.11 南半球以外间断或星散分布 该分布区只有1科,鼠刺科(Escalloniaceae),含1属,2种.
以上阐述及表5可知,永瓣藤植物群落的种类比较丰富.种类较多的科是茜草科,蔷薇科、菊科、蝶形花科、禾本科,其次是山茶科、樟科、葡萄科、漆树科等,这些优势科多属亚热带植物区系的表征科.虽然永瓣藤植物群落科的区系亚热带性质占优势地位,但有一些北温带典型科(如壳斗科、绣球科等)也在其中广泛分布,反映了永瓣藤植物群落区系亚热带成分和北温带成分的交汇,处于亚热带与暖温带的过渡地区,过渡性特征明显.
永瓣藤所在区中含有许多古老的科.裸子植物有产于白垩纪的松科,被子植物有产于白垩纪的桑科,卫矛科,鼠李科,毛茛科,金缕梅科,胡桃科,防已科和杜鹃花科等.第三纪已建立的科有八角枫科,马鞭草科等.而永瓣藤虽然属于卫矛科,但其起源时间约为新第三纪中新世(或更晚),因此是被子植物一个起源比较年轻的属,并不是起源古老的地理成分[32].
3.4 种子植物属(种)的分析
根据吴征镒教授[1-3]关于中国种子植物属的分布区类型系统,可将永瓣藤植物群落区系分为13种分布区类型,分别阐述如下(见表6):
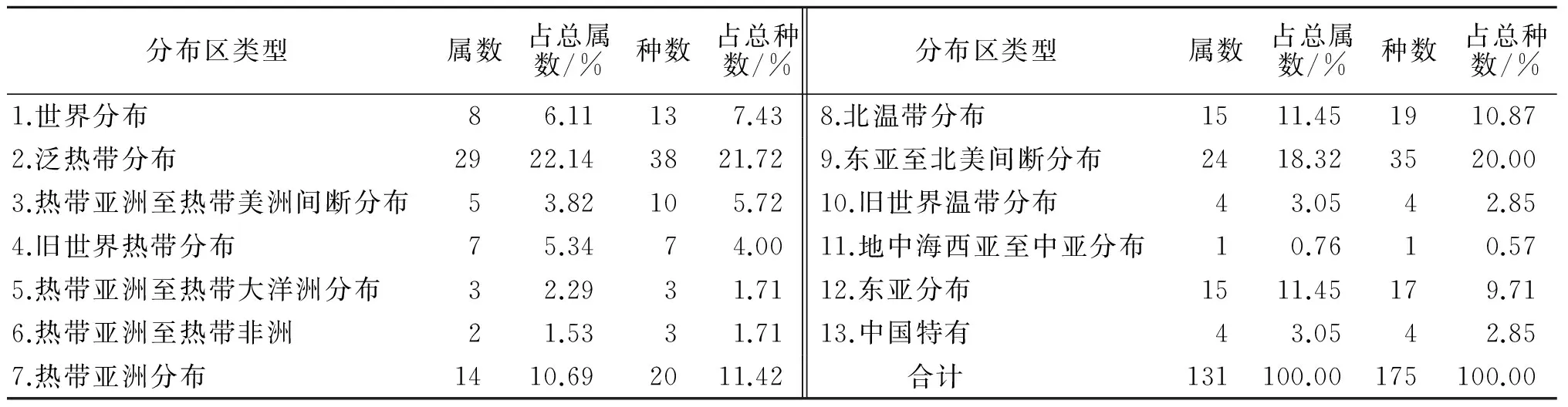
表6 永瓣藤生存群落植物区系种子植物属、种的分布区类型
3.4.1 世界分布 该类型共有8属13种,其中悬钩子属(Rubus)含有6种,其它的水杨梅属(Geum)、鼠李属(Rhamnus)、金丝桃属(Hypericum)、菟丝子属(Cuscuta),各含1种铁线莲属(Clematis),藜属(Chenopodium)苔草属(Carex)各含1科.
3.4.2 泛热带分布 该类型所含属数和种数最多,共有29属,占总属数的22.14%,38种,占总种数的21.71%.其中杜英属(Elaeocarpus)、菝葜属(Smilax)各含3种;紫珠属(Callicarpa)、冬青属(Ilex)、朴树属(Celtis)、薯蓣属(Dioscorea)、榕属(Ficus)各含2种;其它的栀子属(Gardenia)、牡荆属(Vitex)、厚皮香属(Ternstroemia)、檀属(Dalbergia)、崖豆藤属(Millettia)、桂樱属(Laurocerasus)、紫金牛属(Ardisia)、山矾属(Symplocos)、叶下珠属(Phyllanthus)、柞木属(Xylosma)、卫矛属(Euonymus)、花椒属(Zanthoxylum)、南蛇藤属(Celastrus)、金粟兰属(Chloranthus)、鹅绒藤属(Cynanchum)、假蓬属(Conyza)、下田菊属(Adenostemma)、醉鱼草属(Buddleja)、山黄麻属(Trema)、苎麻属(Boehmeria)、狗肝菜属(Dicliptira)、柿属(Diospyros),各含1种.
3.4.3 热带亚洲至热带美洲间断分布 该类型共有5属10科,其中柃木属(Eurya)含4种,木姜子属(Litsea)含3科,其它的楠木属(Phoebe)、泡花树属(Meliosma)、山姜属(Alpinia)各含1科.
3.4.4 旧世界热带分布 该类型共有7属7种,包括乌口属(Tarenna)、八角枫属(Alangium)、野桐属(Mallotus)、乌蔹莓属(Cayratia)、娃儿藤属(Tylophora)、水竹叶属(Murdannia)、扁担杆属(Grewia),各含1种.
3.4.5 热带亚洲至热带大洋州分布 该类型有3属3种,分别是樟属(Cinnamomum)、淡竹叶属(Lophatherum)、兰属(Cymbidium),各含1种.
3.4.6 热带亚洲至热带非洲 该类型仅有2属3种,分别是黄瑞木属(Adinandra),含2种,荩草属(Arthraxon)含1种.
3.4.7 热带亚洲分布 该类型共有14属20种,其中山胡椒属(Lindera)含4种,山茶属(Camellia)含3种,润楠属(Machilus)含2种,其它的构属(Broussonetia)、山茉莉属(Huodendron)、流苏子属(Coptosapelta)、鸡矢藤属(Paederia)、木荷属(Schima)、常山属(Dichroa)、秤钩风属(Diploclisia)、赤杨叶属(Alnipyllum)、南五味子属(Kadsura)、虎皮楠属(Daphniphyllaceae)、新木姜子属(Neolitsea)各含1种.
3.4.8 北温带分布 该类型共有15属19种.其中杜鹃花属(Rhododendron)含3种,紫菀属(Aster)、绣线菊属(Spiraea),各含2种,其它的茜草属(Rubia)、山梅花属(Philadelphus)、蛇葡萄属(Ampelopsis)、枸杞属(Lycium)、李属(Prunus)、栗属(Castanea)、黄精属(Polygonatum)、荚蒾属(Viburnum)、露珠草属(Circaea)、蔷薇属(Rosa)、松属(Pinus)、水青冈属(Fagus),各含1种.
3.4.9 东亚至北美间断分布 该类型所含的属数和种数仅次于泛热带分布类型,共有24属35种,占总属数的18.32%,占总科数的20%,其中石栎属(Lithocarpus)含5种,爬山虎属(Parthenocissus)栲属(Castanopsis),各含3种,勾儿茶属(Berchemia)、绣球花属(Hydrangea)、漆树属(Toxicodendron)、鼠刺属(Itea),各含2种,其它的檫木属(Sassafras)、蕲艾属(Crossotephium)、山蚂蝗属(Desmodium)、胡枝子属(Lespedeza)、络石属(Trachelospermum)、楤木属(Aralia)、木兰属(Magnolia)、金线草属(Antenoron)、木犀属(Osmanthus)、腹水草属(Veronicastrum)、紫藤属(Wisteria)、石楠属(Photinia)、葡萄属(Vitis)、十大功劳属(Mahonia)、南烛属(Lyonia)、六道木属(Abelia)、枫香属(Liquidambar),各含1种.
3.4.10 旧世界温带分布 该类型共有4属4科,分别是石竹属(Dianthus)、菊属(Chrysanthemum)、菘蓝属(Isatis)、梨属(Pyrus),各含1种.
3.4.11 地中海区、西亚至中亚分布 该类型仅有1属1种,为黄连木属(Pistacia).
3.4.12 东亚分布 该类型共有15属17科,其中山麦冬属(Liriope)、刚竹属(Phllostachys),各含2种;其它的三尖杉属(Cephalotaxus)、野鸦椿属(Euscaphis)、蓬莱葛属(Gardneria)、败酱属(Patrinia)、木通属(Akebia)、六月雪属(Serissa)、猕猴桃属(Actinidia)、泡桐属(Paulownia)、南天竹属(Nandina)、南酸枣属(Choerospondias)、继木属(Loropetalum)、青荚叶属(Helwingia)、四照花属(Dendrobenthamia),各含1种.
3.4.13 中国特有分布 该类型含有4属4种,分别是青钱柳属(Cyclocarya)、箬竹属(Indocalamus)、杉木属(Cunninghamia)、大血藤属(Sargentodoxa),各含1种.
综上所述,永瓣藤植物群落属的区系成分以热带成分占绝对优势,共有60属,占总属数的45.80%.在各种热带成分中以泛热带分布类型最多,共有29属,占总属数的22.14%.而各种属又以灌木为主,常见的有檵木属、茶属、杜鹃花属、柃木属、山矾属、山胡椒属等,乔木属次之,常见的有黄连木属、三尖杉属、栎属、栲属、刚竹属等.而温带分布的成分有43属,占总属数的32.82%,一些典型温带属如栗属、杜鹃花属、绣线菊属等也在其中广泛分布.
永瓣藤植物群落的区系中,种的分布也以热带成分占优势,在各种热带成分中以泛热带分布类型最多,共有38种,占总种数的21.17%,而温带分布以东亚北美间断(35)最丰富,占总种数的20%.由此可见,永瓣藤植物群落区系泛热带成分和北温带成分占优势,具有北亚热带向暖温带过渡的特性
4 结论
1)永瓣藤植物群落区系组成多样 经过对永瓣藤所在区域的11个群落植物区系调查、采集和鉴定,发现共有维管植物80科138属182种,其中蕨类植物7科、7属,其它为种子植物,有175种,分别隶属于131属,73科.以蔷薇科、樟科、山茶科、壳斗科等的种类较多.植物集中于几个大科的现象比较明显,优势科在区系组成中占有重要地位.
2)大型科、属较少,中、小型科、属较多 在永瓣藤植物群落的区系中,含10种或10种以上的科4个,占总科数的5.0%,含5~9种的科5个,占总科数的6.25%.含1~4种的科71,占总科数的88.25%.含5~10种的属2个,占总属数的1.45%含1~4种的属136属,占总属数的98.55%.
3)生态类型多样性 永瓣藤的群落分布多为常绿阔叶林或落叶阔叶林的林木,以杉木林、马尾林、毛竹林、杂灌林等四类不同生态类型的群落为主,灌木植物发达.
4)分布区类型多样 永瓣藤植物群落的区系科的分布区含有11个分布区类型,属的分布区含有13个分布区类型.在所有分布区类型中,泛热带分布区类型占首位.
5)泛热带成分占优势,具有北亚热带向暖温带过渡的特征 由上面的分析可知,永瓣藤植物群落种类最多的是茜草科,蔷薇科,菊科,蝶形花科,禾本科,其次是山茶科,樟科,葡萄科,漆树科等,这些优势科多属典型亚热带科.而属(种)的分布也是以泛热带分布类型最多,共有29属(38种),占总属(种)数的22.14%(21.17%).温带成分也很丰富,属、种所占比例与热带成分很相近,一些典型北温带科(如壳斗科,绣球科等)也在其中广泛分布,反映了永瓣藤植物群落区系具有北亚热带向暖温带的过渡的特性,这与永瓣藤分布于北亚热带地区相一致.
6)特有属、种较贫乏 永瓣藤植物群落的区系种类的多样性丰富,但缺乏特有属、种,仅含中国4个特有属、4个特有种(表6).特有属、种能较好反映所在地域植物区系在起源方面的特征[4,33].缺乏特有属、种,表明永瓣藤植物群落的区系的个性特征不明显.
[1] 吴征镒.中国种子植物属的分布区类型[J].云南植物研究,1991,增刊(IV):1-139.
[2] 吴征镒,周浙昆,李德铢,等.中国种子植物科的分布区类型[J].云南植物研究,2003,25(3):245-257.
[3] 吴征镒,周浙昆,孙航,等.种子植物分布区类型及其起源和分化[M].昆明:云南科技出版社,2006:1-566.
[4] 王荷生.植物区系地理[M].北京:科学出版社,1992.
[5] 左家哺,傅德志,彭代文.植物区系的数值分析[M].北京:中国科学技术出版社,1996.
[6] Zhang M L.Studies on geographical distribution pattern of the subgenus Pogonophace (Fabaceae: Astragalus) in China using GIS technique[J].Acta Bo,Sinica,2000,42(8):849-854.
[7] 朱华. 关于地区间植物区系亲缘关系研究方法问题讨论[J].武汉植物学研究,2005,23(4):399-400.
[8] 李锡文,李捷.横断山脉地区种子植物区系的初步研究[J].云南植物研究,1993,15(3):217-231.
[9] 应俊生.秦岭植物区系的性质、特点和起源[J].植物分类学报,1994,32(5):389-410.
[10] 吴素功,杨永平,费勇.青藏高原高寒地区种子植物区系的研究[J].云南植物研究,1995,17(3):233-247.
[11] 周浙昆,孙航,俞宏渊.西藏壳斗科的地理分布[J].云南植物研究,1995,17(2):144-152
[12] 廖文波,张宏达.广东种子植物区系的特有属研究[J].中山大学学报:自然科学版,1995,34(3):73-80.
[13] 廖文波,张宏达.广东种子植物区系的特有属研究[J].中山大学学报:自然科学版,1996,35(6):74-79.
[14] 汤庚国,李湘萍,谢继步,等.江苏湿地植物的区系特征及其保护与利用[J].南京林业大学学报,1997,21(4):47-52.
[15] Xing F W, Ng S C, Chau K C. Gymnosperms and angiosperms of Hong Kong[J]. Memoirs of the Hong Kong Natural History Socity, 2000, 23:21-136.
[16] 彭华,吴征镒.无量山半湿润常绿阔叶林的区系特征及保护生物学意义[J].云南植物研究,2001,23(3):278-286.
[17] 张宏达.海南植物区系的多样性[J].生态科学,2001,20(1):1-10.
[18] 沈泽昊,张新时,金义兴.三峡大老岭植物区系的垂直梯度分析[J].植物分类学报,2001,39(3):260-268.
[19] 孙航.北极-第三纪成分在喜马拉雅-横断山的发展及演化[J].云南植物研究,2002,24(6):671-688.
[20] Zhu H,Ma Y X,Yan L C,et al.The relationship between geography and climateinthe generic-level patterns of Chinese seed plants[J].Acta Phytotaxonomica,2007,45:134-166.
[21] 罗世家.宣恩洪家河流域植物区系分析[J].湖北民族学院学报:自然科学版,2000,18(4):26-28.
[22] 吴玉虎.澜沧江源区种子植物区系研究[J].武汉植物学研究,2009,27(3):277-289.
[23] 陈功锡,邓涛,张代贵,等.湖南德夯风景区峡谷特殊生境植物区系与生态适应性初探[J].西北植物学报,2009,29(7):1 470-1 478.
[24] 曾宪锋,庄雪影,唐光大,等.粤东代表山地铜鼓嶂的种子植物区系研究[J].武汉植物学研究,2009,27(4):381-386.
[25] Rehder A.Monimopetalum,a new genus of celastraceae[J].Jour of the Arnol Arboretum,1926,Ⅶ(4):233-234.
[26] 应俊生,张玉龙.中国种子植物特有属[M].北京:科学出版社,1994.
[27] 谢国文,丁宝章,王遂义.赣北云居山植物区系地理探讨[J].云南植物研究, 1991,13(4):391-401.
[28] 傅立国.中国植物红皮书—稀有濒危植物(第1册)[M]. 北京:科学出版社,1992.
[29] 谢国文. 江西稀有濒危植物资源及其保护[J].植物资源与环境,1994,3(1):52-55.
[30] 谢国文,周芝德.江西种子植物特有属的生物多样性及其保护[J].武汉植物学研究,1996,14(4):294-300.
[31] 谢国文,文林.永瓣藤的分布现状及其保护[J].生物多样性,1999,7(1):15-19.
[32] 谢国文,孙叶根.中国特有的永瓣藤属植物区系地理性质与特征[J].地理研究,1999,18(2):130-135.
[33] 王荷生. 华北植物区系地理[M].北京:科学出版社,1997.
