琯溪蜜柚汁胞ACTIN的分离鉴定和cDNA克隆
2010-01-08钟凤林郭志雄潘东明
钟凤林 郭志雄 潘东明
琯溪蜜柚汁胞ACTIN的分离鉴定和cDNA克隆
钟凤林1,2郭志雄1潘东明2
1.福建农林大学园艺学院 2.福建农林大学园艺产品贮藏保鲜研究所
对琯溪蜜柚粒化与正常汁胞双向电泳分析,获得一个在粒化中上调表达的差异蛋白质点,经过质谱鉴定、生物信息学分析,为肌动蛋白(Actin),以琯溪蜜柚粒化汁胞cDNA为模板,根据Actin氨基酸序列设计引物,进行全长的扩增,得到Actin开放阅读框包含1134碱基,编码377氨基酸。其核酸序列与黑杨派(Populus trichocarpa)、棉花(Gossypium hirsutum)、蓖麻(Ricinus communis)、圆叶锦葵(Malva pusilla) ACT的同源性在89%以上。
琯溪蜜柚 ACTIN 双向电泳 串联质谱 克隆
肌动蛋白(Actin)是真核生物细胞中普遍存在的一种重要的蛋白质,构成细胞骨架中的微丝(Microfilament)系统,参与真核生物细胞的许多重要的生命活动[1]。肌动蛋白是单一多肽链的球状蛋白质(G-actin),由375~377个氨基酸残基所组成,分子量为42kD。生物体中包含7个肌动蛋白基因,编码不同的肌动蛋白同工蛋白质。高等植物肌动蛋白在进化过程中保持高度的保守性和同源性,且具有组织和器官特异性的表达。研究肌动蛋白在拟南芥表皮毛中排列与分布时发现,肌动蛋白对细胞形态与生长极性尤为重要,在表皮毛发育过程中破坏肌动蛋白会引起表皮毛弯曲和分支[2]。Baluska等在研究拟南芥和Rye幼苗时发现,没有F-actin条件下细胞一样可以分裂,但植物会变矮,说明F-actin是细胞伸长所必需[3]。范小平等通过RNAi序列构建载体转化棉花,表明RNAi载体转基因植株棉纤维发育迟缓或受到抑制,但GhACT1只是属于影响纤维生长的多基因中的一个,而不是唯一决定棉纤维长度的基因[4]。
琯溪蜜柚 ((L.) Osbeck) 原产福建省平和县琯溪河畔,是我国蜜柚优良品种之一,有400多年的栽培历史。柑橘果实粒化是成熟期和采后贮藏期的生理病害。其主要表现为汁胞异常膨大,变硬,汁胞木质化,风味变淡,一般情况下为囊瓣近蒂端汁胞先发生粒化,后渐向果心发展,果心处长形汁胞最为严重[5-7]。通过差异蛋白质组学研究粒化与正常汁胞,对获得的差异蛋白质点进行分离、鉴定和cDNA克隆,为从转录水平了解粒化的分子机理奠定了基础。
1 方法
1.1 材料
2007年9月18日,琯溪蜜柚采至福建省平和县小溪村大坑果园,选择大小均匀,色泽一致、无病虫、无损伤的果实。当日从平和寄回实验室,掰开果实,液氮处理、保存、备用。
1.2 双向凝胶电泳
取琯溪蜜柚汁胞,总蛋白的提取、蛋白质含量测定及双向电泳等参照赖呈纯等(2009) 的方法[8]。
1.3 差异表达蛋白点质谱鉴定
差异蛋白点进行脱色、胶内胰酶酶切(in-gel digestion)和肽段提取,然后进行肽质量指纹谱(Peptide Mass Fingerprinting,PMF)分析。检索数据库为NCBInr;检索种属为:all,数据检索的方式为combined;最大允许漏切位点为1;酶为胰蛋白酶。质量误差范围设置:PMF 0.3 Da,MS/MS 0.4 Da;在数据库检索时胰酶自降解峰和污染物质的峰都手工剔除[9,10]。
1.4 琯溪蜜柚汁胞RNA提取及总RNA含量和浓度测定
取琯溪蜜柚汁胞样品,参照钟凤林等方法提取RNA[11]。
1.5 琯溪蜜柚汁胞中ACTIN cDNA ORF的克隆
1.5.1 cDNA合成及引物设计
按Invitrogen RT-PCR、3'和5'-RACE说明书中附加方案说明进行逆转录,合成cDNA。
根据ACTIN [Gossypium hirsutum] (gi|32186906)在蛋白的起始位置和结尾设计特异引物进行cDNA全长扩增。引物由上海博尚生物技术有限公司合成。
ACTR(Nco I): 5'- GCATA CCATGG TTAGAAGCAC TTCCTGTGGA CAATGGA
ACTF(Nde I): 5'- GCATA GTATAC ATG GCC GAT GCT GAG GAT ATT C
1.5.2 PCR扩增
反应程序为:94℃ 5.0min,94℃ 45s,63℃ 45s,72℃1min,35个循环,72℃延伸10 min。
1.5.3序列测定、序列分析
胶回收PCR产物,连接、转化、涂板、筛选和PCR鉴定。菌液送上海英骏公司进行测序。序列通过Expasy、NCBI网址和DNAMAN软件进行分析。
2 结果与分析
2.1 差异蛋白质点APX的获得
正常汁胞和粒化汁胞总蛋白的双向电泳差异比较后,把其中的50个差异蛋白质点经过胶内酶解、MAIDI-TOF-TOF MS以及MASCOT搜索,得到差异蛋白质点为Actin (图1为琯溪蜜柚粒化过程中的差异蛋白质Actin的局部放大示意图)。获得高品质肽质量指纹(PMF)(图2 A),然后根据PMF自动选取5个的峰形最好、强度最高的肽离子送入CID碰撞,得到的碎片离子再用TOF质谱分析获得肽碎片指纹(FFP: fragment fingerprint 图2 B)。使用MASCOT的离子搜索模式搜索蛋白质NCBInr绿色植物蛋白质数据库得到匹配的蛋白(图2 C、D)。Actin其与棉花()actin Score为393。

注:A为正常汁胞;B为粒化汁胞;↑表示表达量上升

注:A为PMF;B为FFP;C为检索值;D为匹配的结果
2.2 Actin cDNA ORF的克隆与分析
2.2.1 RNA的提取及质量分析
用Trizol法提取琯溪蜜柚粒化汁胞总RNA,电泳检测显示(图3,M为DL2000 Marker),分离得到的总RNA的质量较好,条带完整,紫外检测和分析,提取分离得到的总RNA其A260/ A280值为1.90,说明该样品纯度高,适合PCR扩增。
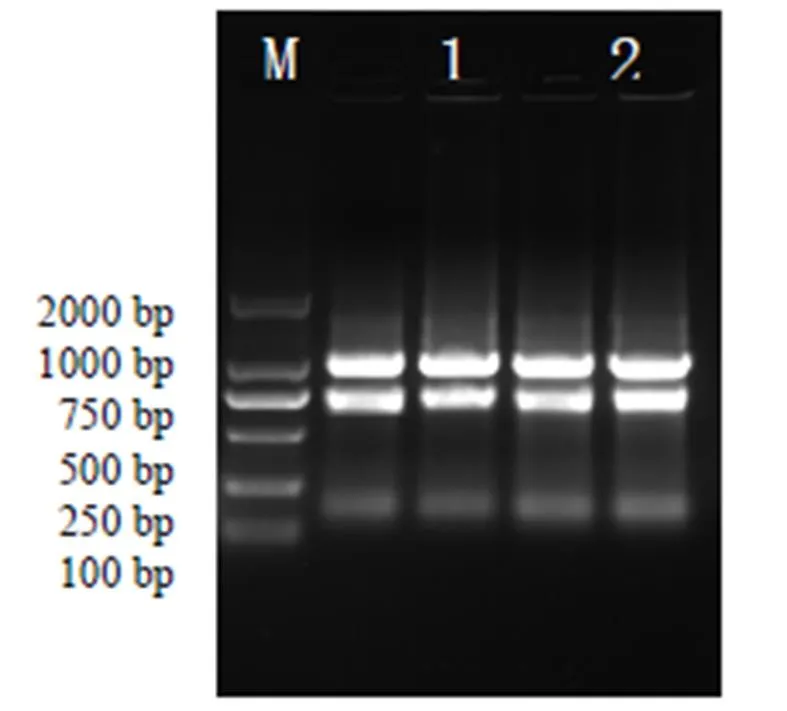
图3琯溪蜜柚汁胞RNA电泳结果
2.2.2 ACTIN ORF 扩增结果
将开放阅读框的两端加上酶切位点和保护碱基后设计引物,以带Oligo-dT的接头引物的cDNA为模板,进行PCR扩增ORF,电泳结果获得了大小约为1100bp的特异产物(图4,M为DL2000 Marker)。胶回收纯化该片段,连接、转化、涂板、筛选和PCR鉴定,送菌液测序。菌液提取质粒后进行测序。

图4 琯溪蜜柚汁胞的ORF电泳图

下划线、方框表示引物区
测序后得到的cDNA序列长为1156,开放阅读框(ORF)共有1134个碱基组成,编码377个氨基酸,ATG为起始密码子,TAA为终止密码子。通过expasy分析,氨基酸位点55~65具有Actins signature1信号,氨基酸位点106~118具有Actins and actin-related proteins signature信号,氨基酸位点358-366具有ACTINS signature2信号(图5)。
序列分析表明,琯溪蜜柚ACTIN cDNA的核苷酸序列以及推导的氨基酸序列与许多植物来源的ACT有很高的同源性。其核酸序列与黑杨派()的ACT1、ACT9、棉花()ACT1、ACT11同源性高达90%,与蓖麻()ACT、圆叶锦葵() ACT、棉花()ACT4、ACT8、ACT9的同源性89%。
2.2.3 Actin ORF推导的氨基酸与其它植物Actin氨基酸序列比对
由DNAMAN软件分析可知,Actin cDNA编码377个氨基酸,理论分子量为41.7014KD,理论等电点(pI)为5.45,属酸性或阴离子型。经由DNAMAN软件分析比对后得到与黑杨派(Populus trichocarpa)的ACT1、ACT9、棉花(Gossypium hirsutum)ACT1、ACT11等的氨基酸序列比对图(图6),可知:ACT 与推导的氨基酸的同源性都达到了99%以上。
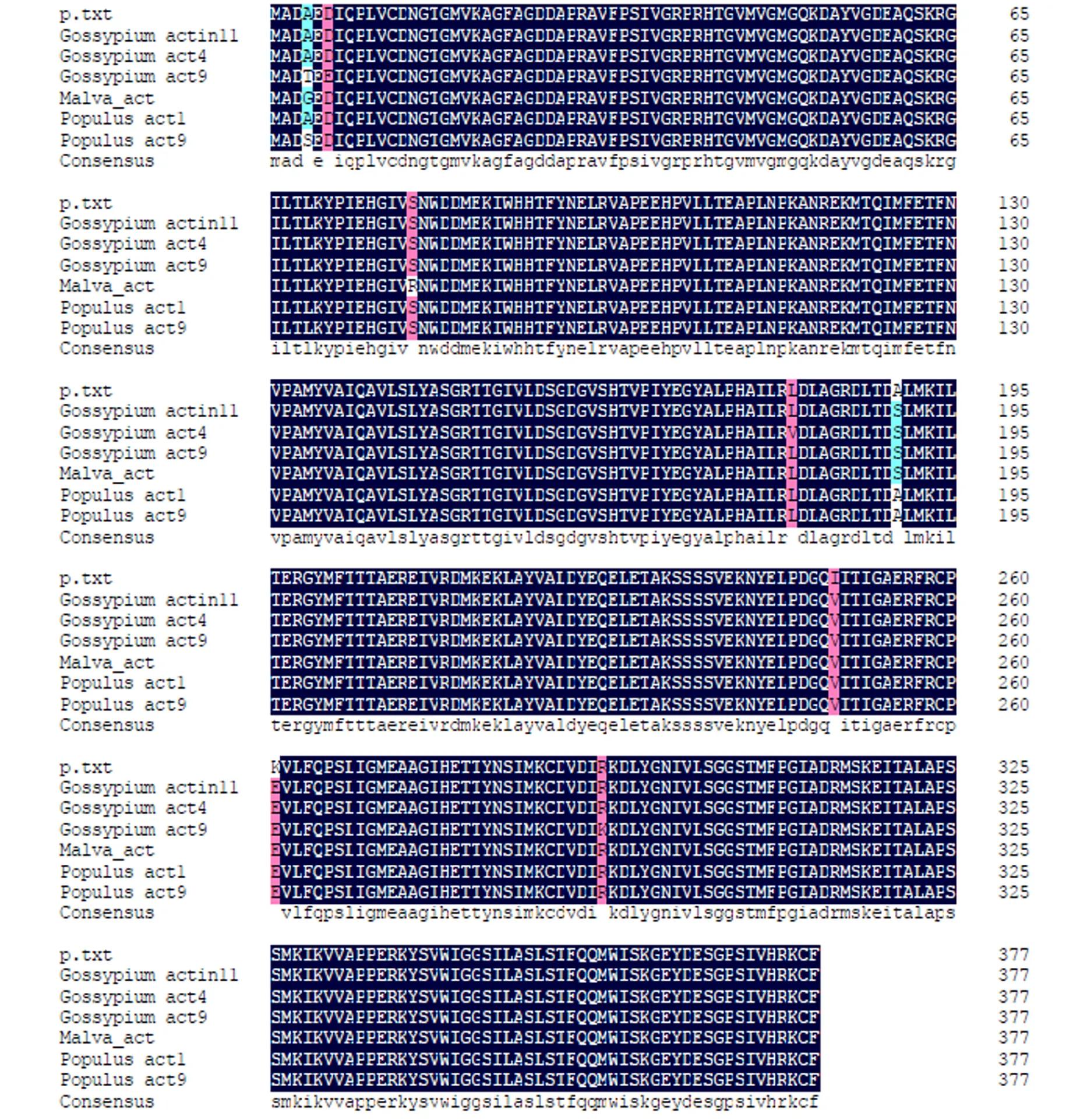
图6 Actin cDNA编码氨基酸序列与其它植物ACTIN的多重序列比对
3 讨论
在高等植物中,肌动蛋白在进化过程中保持高度的保守性和同源性,以基因家族形式存在,可分为营养体基因与生殖细胞基因,并且肌动蛋白的表达有组织、器官特异性和发育阶段特异性[12-14]。肌动蛋白骨架对植物高度特异化的细胞的形态发生和功能起着重要作用,同时在高等植物形态发生的细胞分裂和伸长过程中起着关键的调节作用[15,16]。Li等从cDNA文库中筛选出15个actin基因中,real-time PCR结果表明有5个肌动蛋白基因在纤维中特异表达,推断在GhACT1缺失时,其他4个基因仍正常行使功能,弥补和维持纤维生长,使纤维继续伸长,尤其在伸长期后期GhACT1的缺失对纤维伸长的影响甚微[17]。
本研究通过分离、鉴定和克隆,获得了Actin的cDNA全长,并表明在琯溪蜜柚汁胞发育阶段的粒化汁胞中Actin表达的变化,可能是Actin与肌球蛋白的相互作用,参与了细胞运动有关活动,使得琯溪蜜柚汁胞生长与延伸发生改变,猜测其对构建细胞骨架和适应多样胁迫环境变化有着重要作用。有关这方面的有待进一步进行。
[1] Staiger C J,Lloyd C W.The plant cytoskeleton[J].Curr Opin Cell Biol,1991,3:33-42.
[2] Huang S,An Y,Mcdowell J M,et al.TheACT11 actin gene is strongly expressed in tissues of the emerging inflorescence,pollen and developing ovules[J].Plant Mol Biol,1997, 33:125-139.
[3] Baluska F, Jasik J, Edelmann H G, et al. Latrunculin B-induced plant dwarfism: plant cell elongation is F-actin-dependent[J]. Developmental Biology, 2001, 231(1): 113-124.
[4] 范小平,范博红,李学宝,等.肌动蛋白基因在棉花纤维中的作用研究[J].华北农学报,2008,23(5):73-75.
[5] 潘东明,郑国华,陈桂信,等.琯溪蜜柚汁胞粒化原因分析[J].果树学报,1999,16(3):202-209.
[6] 钟凤林,郭志雄,潘东明,等.琯溪蜜柚汁胞APX1cDNA的克隆及原核表达[J].热带作物学报,2010,31(1):76-82.
[7] 钟凤林,郭志雄,李开拓,等.琯溪蜜柚汁胞在发育和粒化过程中APX活性变化及同工酶分析[J].热带亚热带植物学报,2010,(1):27-31.
[8] 赖呈纯,赖钟雄,何园,等.龙眼胚性培养物高分辨率蛋白质双向电泳技术[J].福建农林大学学报,2008,37(1):37-41.
[9] 周玮,刘晓慧,周新文,等.糖蛋白结构质谱解析的样品前处理[J].色谱,2007,25(5):623-627.
[10] 谢锦云,李小兰,陈平,等.温敏核不育水稻花药蛋白质组初步分析[J].中国生物化学与分子生物学报,2003,(19)
[11] 钟凤林,郭志雄,李开拓,等.琯溪蜜柚汁胞RNA提取[J].生物技术通报,2010,(2):109-112.
[12] Heslop-Harrison J, Heslop-Harrison Y. Intracellular motility, the actin cytoskeleton and germinability in the pollen of wheat (Triticum aestivum L.)[J]. Sexual Plant Reproduction, 1992, 5(4): 247-255.
[13] O'Connor P. Actin-dependent cell elongation in teleost retinal rods: requirement for actin filament assembly[J]. Journal of Cell Biology, 1981, 89(3): 517-524.
[14] Szymanski D B,Marks M D,Wick S M.Organized F-actin is essential for normal trichome morphogenesis in Arabidopsis[J].Plant Cell,1999,11:2331-2347
[15] Dong C H, Xia G X, Hong Y, et al. ADF proteins are involved in the control of flowering and regulate F-actin organization, cell expansion, and organ growth in Arabidopsis[J]. The Plant Cell 2001, 13(6): 1333-1346.
[16] 李岩, 徐是雄. 百合花粉及花粉管内微丝和微管的分布[J].植物学报, 1998, 40(010): 890-894.
[17] Li X B, Fan X P, Wang X L, et al. The cotton ACTIN1 gene is functionally expressed in fibers and participates in fiber elongation[J]. The Plant Cell, 2005, 17(3): 859-875.
