辣椒种质亲缘关系的数量分类学研究
2009-10-13陈学军方荣周坤华等
陈学军 方 荣 周坤华等
摘 要:基于29个表型性状对辣椒属(Capsicum)5个栽培种的60份种质进行了聚类分析,结果显示:60份材料两两不同种质间遗传距离在1.376~7.321之间,平均为4.112;以遗传距离4.412为阀值,60份种质被分为4大组,即C. annuum+C. chinense组、C. baccatum组、C. frutescens组和C. pubescens组,表明C. annuum与C. chinense亲缘关系最近,而与C. pubescens最远; C. annuum5个变种在4.013阀值处分为2组,长椒(var. longum)、灯笼椒(var. grossum)和樱桃椒(var. cerasiforme)在同一组。本文还就C. annuum变种的演化进行了探讨。
关键词:辣椒属;遗传差异;数量分类学;亲缘关系
在辣椒属(Capsicum)的5个栽培种中,一年生辣椒(C. annuum)是栽培最广泛、类型最丰富的种,包括长椒(var. longum)、灯笼椒(var. grossum)、圆锥椒(var. conoides)、樱桃椒(var. cerasiforme)和簇生椒(var. fascicutatum)5个变种,我国栽培辣椒绝大多数都属于C. annuum[1]。为探明辣椒属种间及种内遗传变异,国内外学者基于形态学标记[2]、细胞学标记[3, 4]、生化标记[5, 6])和DNA分子标记[1, 7-9]对辣椒属系统发育进行了大量研究,但基于表型数据的数量分类学研究却少有报道。为此,本文应用数量分类学方法,对辣椒属5个栽培种的60份不同类型材料进行了亲缘关系分析,旨在为辣椒种质资源的有效利用和杂交品种的选育提供参考。
1 材料与方法
1.1供试材料
在辣椒属5个栽培种中,选取有代表性的材料共60份。其中C. annuum种质51份,分属于5个变种。C. chinense和 C. frutescens种质各2份,包括1份中国海南C. frutescens种质。其他2个种(C. baccatum和C. pubescens)各1份,种间材料(C. annuum×C. chinense)3份。材料编号、名称、来源及种类列于表1。试验在江西省农业科学院蔬菜花卉研究所试验园进行,上述材料2008年3月2日大棚播种育苗,4月20日大棚定植。采用随机区组试验设计,重复3次,每份材料每小区随机选取5株进行定株、定时观测记录,对于数量性状取其平均值进行统计分析。
1.2表型性状的选取与编码处理
根据5个栽培种主要性状的差异,选取29个有分类价值的性状进行编码处理,具体参照陈学军等的方法进行[10](表2)。
1.3 数据分析
采用SPSS软件进行数据分析处理。数据的标准化使用STD(标准差标准化)命令,采用欧氏(Euclidean)距离系数计算遗传距离,聚类方法采用Between-groups linkage法。
2 结果与分析
2.1遗传距离
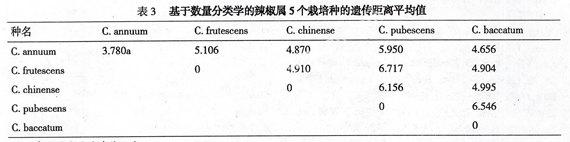
以60份辣椒材料和44个分解性状的数据为原始矩阵,共获得1770个两两不同种质间的遗传距离系数,遗传距离最大者为7.321(种质37与41),最小者为1.376(种质26与27),平均为4.112;材料26与27均为长椒(var. longum)中的线椒种质;41为圆锥椒(var. conoides)种质,37则为C. pubescens种质。
从表3看到,C. annuum 种内遗传距离平均为3.780,而5个栽培种种间遗传距离都大于4.870,可见辣椒属栽培种种间遗传变异大于C. annuum种内遗传变异。此外,3个种间杂交组合29、30和31与母本(C. annuum)的遗传距离平均为3.699,与父本(C. chinense)的遗传距离平均为3.791,说明种间杂种(F1)的基因型界于双亲之间。



2.2聚类分析
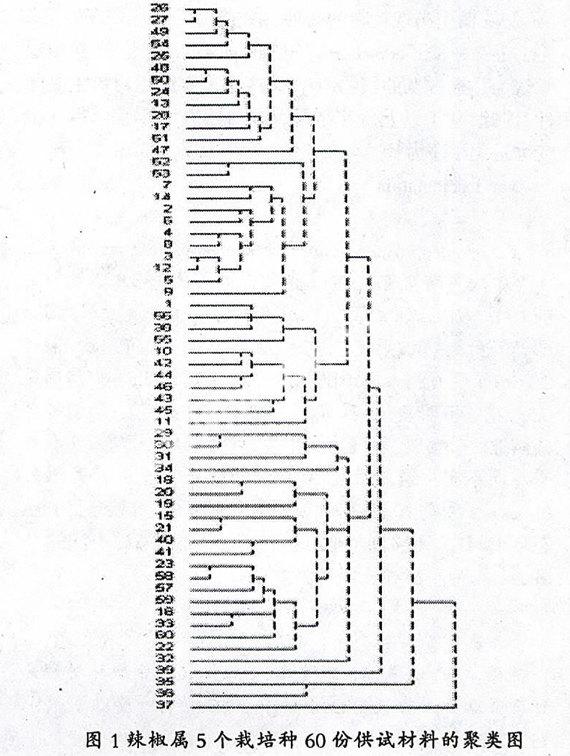
以4.412的遗传距离系数为阈值,所有供试材料被分为4大组群,即C. annuum+C. chinense组、C. baccatum组、C. frutescens组和C. pubescens组。在C. annuum+C. chinense组中,包含所有的51份C. annuum材料、3份C. annuum与C. chinense种间杂种及2份C. chinense种质,种质35和36则构成C. frutescens组,种质39和37分别聚为C. pubescens组和C. baccatum组(图1)。
在4.013的遗传距离系数处,C. annuum+C. chinense组进一步分为5个亚组,第一亚组由长椒、灯笼椒和樱桃椒种质组成,在该亚组中,25份长椒种质在3.551的遗传距离处聚为一类,但也有3份长椒种质10、11和55在3.309处与灯笼椒、樱桃椒种质聚为一类,种质10果实牛角形,老熟果黄色,肉厚无辣味,11和55均为短粗牛角果、肉厚。第二亚组包括C. chinense种质34与3个种间杂种29、30、31。第三亚组有簇生椒种质4份和圆锥椒种质11份,该亚组种质果实均朝上生长。第四亚组只有1份C. chinense种质32,该材料果实单生,果面皱缩不平,与种质34表型有一定差异。
3 讨论
本研究基于29个表型性状,利用数量分类学方法对辣椒属5个栽培种共60份材料进行了聚类分析,结果显示:材料间遗传距离变幅较大,从1.376~7.321,说明辣椒属5个栽培种在进化过程中保持了较高的遗传变异。在5个栽培种中,C. annuum与C. chinense亲缘关系最近,而与C. pubescens最远,这与陈学军等[1]基于RAPD分子标记的聚类结果是一致的。
在供试的51份C. annuum材料中,灯笼椒(var. grossum)种质与长椒(var. longum)、樱桃椒(var. cerasiforme)首先聚为一类,而圆锥椒(var. conoides)则与簇生椒(var. fascicutatum)相聚,这说明在C. annuum的5个变种中,灯笼椒种质、长椒和樱桃椒三者亲缘关系较近,而与圆锥椒和簇生椒亲缘关系稍远。核型研究表明,var. conoides和var. longum为2A核型,是较原始的类型;而var. cerasiforme和var. grossum是2B核型,是由var. conoides派生出现较晚的变种,并认为其进化顺序为 var. conoidess → var. cerasiforme → var. grossum [11-14]。辛辣椒通常认为是最古老的植物[15],而辣椒果实簇生对单生是隐性,因此,基于以上研究结果,作者认为,一年生辣椒5个变种进化顺序为:
var. fascicutatum
↑
ar. conoides→var. longum→var. cerasiforme→var. grossum
中国海南C. frutescens种质在遗传距离3.372处与美洲C. frutescens种质聚为一类。该种质植株高大、叶阔卵形,花冠绿白色,果实单生,果顶直立向上,纺锤状、果长2.5cm,果径0.8 cm,果肉厚0.05 cm,果柄长2.5 cm,单果重0.5g,花萼不明显,晚熟,味辣,在海南为多年生;但引种至南昌地区栽培时,表现为一年生,且对长日照敏感,在春夏季花而不实,越夏后在10月份才能正常座果,美洲C. frutescens种质也有此特性。我国海南地区的气候条件和生态环境与中、南美洲类似,这可能是这2个来源不同的C. frutescens种质都对长日照敏感的原因。
参考文献
1. 陈学军,陈劲枫,耿红,娄群峰.辣椒属5个栽培种部分种质亲缘关系的RAPD分析[J] .园艺学报,2006,33(4):751~756
2. Egawa Y, Tanaka M. Cytogenetical study of the interspecific hybrid between Capsicum annuum and C. baccatum[J]. Japan Journal of Breeding, 1986, 36: 16~21.
3. Pickersgill B. The genus Capsicum: a multidisciplinary approach to the taxonomy of cultivated and wild plants[J]. Biol Zent Bl., 1988, 107: 381~389.
4. Pickersgill B. Cytogenetics and evolution of Capsicum[A]. In: Tsuchiya T, Gupta P K, eds. Chromosome engineering in plants: genetics, breeding, evolution. Part B[C]. Amsterdam: Elsevier, 1991, 139~160.
5. Mcleod M J, Guttman S I, Eshbaugh W H, et a1. An electrophoretic study of evolution in Capsicum (Solanaceae) [J]. Evolution, 1983, 37(3): 562~574.
6. Lcaiza-Figueroa F K, Ritland J A , Laborde Cancino, et a1. Pattern of genetic variation of the genus Capsicum (Solanaceae) in Mexico[J]. Pl Syst Evol, 1989, 165: 159~188.
7. Prince J P, Lackney V K, Angeles C, et a1. A survey of DNA polymorphism within the genus Capsicum and the fingerprinting of pepper cultivars[J]. Genome, 1995, 38: 224~231.
8. Rodriguez J M, Berke T, Engle L, et a1. Variation among and within Capsicum species revealed by RAPD markers[J]. Theor Appl Genet, 1999, 99: 147~156.
9. 张璐,杨若林,刘文轩等.辣椒部分栽培种遗传相似性的RAPD分析[J] .上海大学学报(自然科学版),2003.9(5):433~437.
10. 陈学军,程志芳陈劲枫,等.辣椒种质遗传多样性的 RAPD和 ISSR及其表型数据分析[J] .西北植物学报,2007,27(4):0662~0670.
11. 李光涛,梁涛. 4种(变种)辣椒的核型研究[J] .广西植物,1993,13(2):155~158.
12. 李光涛,梁涛.两种辣椒的核型研究[J] .西南农业大学学报,1992,14(1):82~85.
13. 李林初,沈敏健,李懋学.辣椒的核型研究[J] .园艺学报,1984,1(2):1l9~121.(略)
