内关、公孙协同作用的神经解剖学研究
2000-06-13林锓武陈以国
林锓武 陈以国
(上海医科大学华山医院中西医结合研究所,上海200040;1辽宁中医学院针灸系)
摘 要 运用CB睭RP神经示踪法对内关、公孙配伍机理从神经解剖学角度进行探讨。结果:内关、公孙配伍运用,其针刺信息可在脊髓内经中间内、外侧核的神经元纤维感传至相应脊髓节段,再分别通过与交感神经、副交感神经形成突触联系,形成在脊髓层次的协同增效关系;同时通过两穴在脊髓内相应神经元向孤束核的投射纤维产生突触联系,实现对胃等内脏传入信息在脊髓和孤束核水平的调节整合作用。
主题词 穴,内关/解剖组织学 穴,公孙/解剖组织学 神经解剖学内关为手厥阴心包经穴,公孙为足太阴脾经穴,此上下两肢、经脉不同、距离很远的两穴,在针灸临床上却常一起配伍运用,自古以来作为"八脉交会穴"的用穴之一,其机理何在?中医认为此两穴是通过心包经、脾经、阴维脉及冲脉交汇于胃、心、胸部位,来实现其治疗作用上的协同增效关系。历来对于内关、公孙的研究,多为中医传统理论探讨、临床应用观察及单穴实验研究,而对于两穴配伍相关性的研究,一直缺乏实验佐证。除了其它假说,目前大多认为经络腧穴实质与神经体液关系较大。所以,设想内关、公孙配伍能有效地治疗胃等内脏疾病,是否与两穴的神经纤维投射在中枢神经内的延伸与联系有关?两穴区的神经节段支配关系可能在中枢神经系统中有着一定的联系。本文应用霍乱毒素B亚单位(choleratoxinsubunitB)耦联辣根过氧化物酶(CB睭RP)法对大鼠"内关"、"公孙"单穴及两穴配伍后,在相应运动神经元的分布和树突构筑进行研究,以期对配穴的理论与临床实践提供一些神经解剖学基础。1 材料和方法本实验用中国协和医科大学合成的霍乱毒素B亚单位耦联辣根过氧化物酶(CB睭RP)作为神经解剖学探针;实验动物由中国医学科学院基础研究所动物繁育场提供的Wistar成年雌性健康大鼠,体重在200~250g之间,每组8只,并根据比较解剖学方法,选取大鼠"内关"、"公孙"穴位。
1.1 实验分组第1组:"内关"穴区注射CB睭RP;第2组:"内关"穴区注射CB睭RP后电针同侧"公孙";第3组:"公孙"穴区注射CB睭RP;第4组:"公孙"穴区注射CB睭RP后电针同侧"内关";第5组:胃壁注射CB睭RP;第6组:胃壁注射CB睭RP后电针右侧"内关"、"公孙"。
1.2 实验方法(1)第1组与第2组:先将0.3%CB睭RP5μl用微量注射器注射至右侧"内关"穴区(注射CB睭RP前先将动物麻醉),第2组待动物清醒后配伍电针右侧"公孙"穴,使用ZYZ1型多功能针灸仪,频率为2~100c/s,疏密波,强度调在低档2~4之间,以动物能耐受为限,每次电针20min,每日1次,存活72h后灌注固定,取同侧脊髓C3~S2节段。第3组与第4组:将0.3%CB睭RP5μl用微量注射器注射至右侧"公孙"穴区,第4组待动物清醒后配伍电针右侧"内关"穴20min,每日1次,存活84h后灌注固定,取同侧脊髓C3~S3节段。第5组与第6组:将0.3%CB睭RP10μl用微玻管套接微量注射器多点注射于胃的前壁、后壁、幽门和贲门的浆膜下与肌层之间,第6组待动物清醒后再行电针右侧"内关"、"公孙"穴20min,每日1次,存活48h后灌注固定,取延髓、腹腔神经节和C4~L3节段背根神经节。(2)固定、取材及CB睭RP酶组化过程[1]:①固定标本:麻醉动物(戊巴比妥钠30mg/kg腹腔注射)后,将灌注针经由左心室插入至主动脉并固定,剪开右心耳。按顺序灌注下列液体(须将气泡排尽):a.21℃生理盐水150ml速灌,灌注压为100cmH2O。b.4℃的固定液500ml,先快速灌注1/3后,再慢速灌注,总灌注时间为60min。②取材及切片:小心剥离需要的神经组织(延髓、脊髓、背根神经节、腹腔神经节),投入30%的蔗糖PB缓冲液过夜。以冠状或水平切面冰冻切片,片厚40μm,接片于0.1MPB缓冲液中。③CB睭RP酶组化过程:将切片以双蒸水洗6遍后,以四甲基联苯胺(TMB)法[2]显色,光学显微镜明、暗视野观察,观察后在暗视野下照相。
2 实验结果第1组:脊髓前角中CB睭RP标记细胞的节段性分布,在C6~8节段Rexed分层属于第Ⅸ层中发现阳性标记运动神经元和神经终末纤维。每张切片可见1~5个阳性标记细胞,其树突野可达第Ⅶ层。
第2组:脊髓背角中CB睭RP标记细胞的节段性分布及其特点,a.经配伍电针同侧"公孙"穴后,"内关"穴区阳性标记运动神经元分布节段扩大,可在C4~8节段的第Ⅸ层中出现;其树突野扩大,向第Ⅶ、Ⅵ、Ⅴ层延伸,其中C8节段树突达第Ⅶ层,并向其中间内侧核、中间外侧核特异性延伸。b.在纵切面切片中见到其树突野向纵向、横向延伸和联系明显加强,并可在阳性标记运动神经元周围见到大量神经终末标记(见图1,图2)。
图1 显示"内关"穴区配伍电针后在C6节段阳性标记Mn之树突野延伸至第Ⅶ、Ⅵ、Ⅴ层 10×6.3
图2 显示"内关"穴区配伍电针后在纵切面切片中可见阳性标记Mn显著增多及其树突野明显地向纵向横向联系和可见周围的神经终末标记 16×6.3
第3组:脊髓前角中CB睭RP标记细胞的节段分布,在L4~6节段第Ⅸ层中出现阳性标记的运动神经元和神经终末纤维。每张切片可见1~7个阳性标记细胞,其树突野可达第Ⅶ层。
第4组:脊髓前角中CB睭RP标记细胞节段性分布及其特点,a.经配伍电针同侧"内关"穴后,"公孙"穴区阳性标记运动神经元节段分布扩大至L4~S3节段,于其第Ⅸ层出现阳性标记细胞和神经终末纤维,其树突向第Ⅶ、Ⅵ、Ⅴ层特异性延伸。b.在中间内侧核、中间外侧核出现阳性神经纤维和终末标记。c.纵切面中可见到阳性标记运动神经元增多和树突野明显地向纵向、横向延伸及相互联系。d.S2,3节段阳性标记运动神经元树突特异性延伸至中间外侧核(骶副交感核,见图3~图5)。
第5组:a.在孤束核内发现阳性标记细胞及大量神经终末纤维,为多角型细胞,两侧基本对称。b.腹腔神经节出现阳性标记神经元,以大、中型为主,呈梭型、多角型,散在分布;可见到阳性标记神经纤维。c.在C4~L3节段背根神经节中可出现阳性标记细胞,以T5~T9节段最多。第6组:a.相同的存活期内,配伍电针后在孤束核的阳性标记细胞和神经终末纤维均不及第5组,其阳性标记细胞数明显减少(P<0.01,见表1)。b.配伍电针后,腹腔神经节内阳性标记细胞明显少于单纯注射CB睭RP组(P<0.05,见表1),并可见阳性标记神经纤维减少。c.配伍电针后,仅在4只大鼠T8,T9节段背根神经节出现少许阳性标记细胞。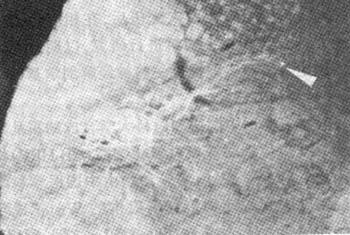
图3 显示"公孙"穴区配伍电针后在L6节段阳性标记Mn树突野延伸至第Ⅶ、Ⅵ、Ⅴ层 10×6.3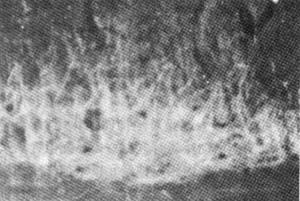
图4 显示"公孙"穴区配伍电针后在纵切面切片中可见阳性标记Mn显著增多及其树突野明显地向纵向横向联系和可见周围的神经终末标记 16×6.3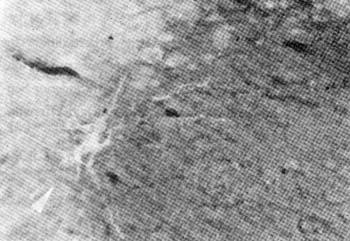
图5 "公孙"穴区配伍电针后在S2,S3节段阳性标记Mn树突特异性延伸至中间外侧核(骶副交感核) 10×6.3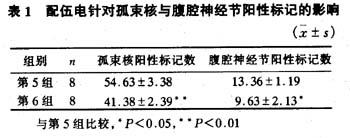
讨论
3.1 内关与交感神经系统的神经解剖学联系本实验采用对比的方法设立"内关"穴区注射CB睭RP组和"内关"穴区注射CB睭RP后配合电针"公孙"穴组。结果,第1组与第2组"内关"注射CB睭RP都能在脊髓C6~C8节段发现阳性运动细胞,但第2组配伍电针"公孙"穴后,不但脊髓前角运动神经元阳性标记细胞的节段性分布扩大,运动神经元阳性标记细胞出现在第Ⅸ层,而且,其树突野特异性延伸至与内脏活动有关的第Ⅶ(中间外侧核、中间内侧核)、Ⅵ、Ⅴ层。此结果提示针刺可增加CB睭RP阳性细胞标记作用,不但存在于相同的神经节段,也存在着跨节段的增强效应。配伍"公孙"电针后能明显加强"内关"穴区运动神经元与交感神经系统的联系,这一效应可能是通过中间内、外侧核内神经元间的相互影响来实现的。
3.2 公孙与副交感神经系统的神经解剖学联系本实验首次应用CB睭RP对大鼠"公孙"穴区进行神经解剖学角度的探讨,结果当配伍电针"内关"穴时,"公孙"穴区逆行阳性标记细胞的节段性分布扩大,其中阳性标记运动神经元的树突野向第Ⅶ、Ⅵ、Ⅴ层特异性延伸,在中间内侧核出现神经纤维,特别是S2,S3节段阳性标记运动神经元树突特异性延伸至中间外侧核(骶副交感核),提示配伍电针"内关"后,对"公孙"穴区相应的运动神经元与副交感神经系统之间的联系有正向调节作用。
3.3 关于第Ⅸ层阳性标记运动神经元周围的神经终末纤维本实验发现第Ⅸ层阳性标记细胞周围出现神经终末纤维,这与Sterling和Kuypers在猫下位颈节的后根纤维终入全部9层中,其中有相当数量的后根纤维终止于运动神经元的实验结果相同[3,4]。因此我们可以进一步推测,当针刺内关、公孙穴时,针感可能经由传入纤维到达第Ⅸ层而与前角中的运动神经元形成单突触或多突触联系,促使运动神经元树突特异性地延伸,而与相应的神经元纤维产生联系,其中一部分纤维将冲动向上传导,形成受针对象(大脑皮层)的得气感,一部分纤维将冲动向下传导形成施针者针下的得气感[5],另有一部分纤维特异性地与第Ⅶ(中间外侧核、中间内侧核)、Ⅵ、Ⅴ等与内脏支配有关的板层发生突触联系,形成两穴配伍在脊髓层次的整合,从而实现对胃等内脏器官功能的调节作用。
3.4 关于内关、公孙两穴配伍对胃等内脏的调节作用自Ellison和Clark[6]成功地将HRP示踪技术应用于内脏神经以来,国内外已有不少有关胃等内脏的神经支配报道,并在包括孤束核在内的不同节段追踪到HRP阳性标记细胞。孤束核(Sol)是延髓内重要的内脏感觉性核团,为一般内脏感觉传导通路上的第一级中继站。随着神经解剖学和神经生理学研究方法的不断发展,人们对孤束核的认识日趋深入,近年来HRP法以及还原银染技术对有关孤束核的大量研究证实了孤束核不但与脊髓、脑干具有传入、传出神经联系,而且与高级中枢(前脑、小脑)也具有复杂的往返联系[7]。Lowey认为孤束核向脊髓的投射主要起自孤束核的中间内侧核、腹外侧核、联合核[8]。而孟氏用HRP法证实了脊髓灰质Ⅲ~Ⅶ、Ⅹ板层向孤束核内侧部直接投射[9]。本实验可见到"内关"、"公孙"两穴配伍电针后,两穴的阳性标记运动神经元树突野延伸至Ⅶ、Ⅵ、Ⅴ板层,并特异性地与中间外侧核、中间内侧核联系。因此推测内关、公孙与胃等内脏之间可能存在着内关、公孙→躯体传入神经→第Ⅸ板层→第Ⅶ、Ⅵ、Ⅴ板层,中间外侧核、中间内侧核→孤束核→交感神经、副交感神经→胃等内脏的调节关系。
胃壁注射CB睭RP是对胃的病理性刺激,其传入应比生理状态下增加,而配伍电针内关、公孙后,可见孤束核内阳性标记细胞显著减少,正说明了内关、公孙配伍可对病理状态下胃等内脏的中枢传入起显著地抑制性作用;而当胃等内脏功能活动减弱时,内关、公孙配伍针刺又可以兴奋孤束核内相应神经元以增加交感和副交感神经对内脏的传出冲动,从而实现对胃等内脏功能的双向调节作用。内关和公孙之间通过脊髓的中间内、外侧核神经元纤维的相互影响,实现特异的增加效应,这可能是两穴配伍运用能增强临床效果的机制所在,而孤束核则起着实现两穴对胃等内脏调节的中枢整合作用。
4 参考文献
1MesulamM.M.TetramethylBenzidineforHorseradishPeroxidaseNeurohistochemistry:ANoncarcinogenicBlueReactionProductwithSuperiorSensitivityforVisualizingNeuralAfferentsandEfferents,J.Histochem.Cytochem.1978;26:106
2 鞠躬,万选才,董新文编著.神经解剖学方法.北京:人民卫生出版社,1985:76
3 T.L.皮尔著.第二军医大学主译.临床神经解剖学基础北京:人民卫生出版社,1980;77
4 REXED,B.SomeAspectsoftheCytoarchitectionicsandSynaptologyoftheSpinalCord.BrainRes.1964;11:58
5 HongchienH.A.,KaoT.,Tan,E.C.,etal.MuscleSensoryNeuronsintheSpinalGangliaintheRatasDeterminedbytheRetrogradeTransportofHorseradishPeroxi瞕ase.Anat.Rec.1979;193:557
6 EllisonJ.P.,ClarkG.M.RetrogradeAxonalTrans瞤ortofHorseradishPeroxidaseinPeripheralAuto瞡omicNerves.J.Comp.Neurol.1975;161:103
7 RicardoJ.A.,KohE.T.AnatomicalevidenceofDi瞨ectProjectionsfromtheNucleusoftheSolitaryTracttotheHypothalamus,Amygdala,andOtherForebrainStructuresintheRat.BrainRes.1978;153:1
8 LoweyA.D.,BurtonH.NucleioftheSolitaryTract:EfferentProfectionstotheLowerBrainStemanelSpinalCordoftheCat.JCompNeurol.1978;181:421
9 孟卓.大鼠脊髓灰质向孤束核的投射HRP法研究.神经解剖学杂志,1986;15(3):227(收稿日期:19981202,成平发稿)
