提高猪体细胞克隆胚胎发育率的研究进展
2024-12-31张才用陈指龙谢浩彭翠婷晏超赵玉兰齐霖周磊唐中林


摘要:体细胞克隆技术可使终末分化的体细胞获得完整全能性,进而生产出完整个体。猪体细胞克隆技术在种质资源的保护与利用及制备基因编辑猪等方面具有广泛的应用前景。然而,猪体细胞克隆的胚胎发育率和移植存活率限制着该技术在实际生产中的推广应用。随着微量细胞测序技术的发展,克隆胚胎发育异常机制不断被揭示,诸多研究人员尝试通过改良核移植操作程序或纠正克隆胚胎的异常重编程来提高克隆成功率,取得了一定效果。综述了近年来围绕卵母细胞、供体细胞和克隆胚胎3个方面来提高猪体细胞克隆胚胎发育率的相关研究,以期为提高猪体细胞克隆效率提供参考,有助于推动体细胞克隆技术在猪生产中的应用。
关键词:猪;体细胞;核移植;胚胎;发育doi:10.13304/j.nykjdb.2024.0209
中图分类号:S828 文献标志码:A 文章编号:10080864(2024)12020109
体细胞克隆又称体细胞核移植(somatic cellnuclear transfer,SCNT),是一种可使移入去核卵母细胞的终末分化体细胞获得全能性,进而生产出与供体细胞相同基因型个体的技术。自1996年克隆羊“Dolly”问世以来[1],通过SCNT技术获得克隆后代的哺乳动物已超过20多种,特别是我国科学家率先完成体细胞克隆猴的研究,实现了历史性突破[2]。克隆猪的历史可追溯至1989 年,Prather等[3]将4-细胞胚胎的细胞核移入去核卵母细胞,获得的重构胚胎移植至受体母猪体内继续发育诞生一头克隆仔猪。由于猪SCNT技术发展缓慢,直到2000年才获得第一头体细胞克隆猪[4]。SCNT发展至今,在保护珍贵或濒危猪种、快速繁育优良种猪和加快生猪遗传改良等方面发挥着重要的作用[5]。此外,SCNT 与基因编辑技术相结合,生产的基因辑编猪已应用于异种器官移植、再生医学和抗病育种等领域[6]。然而,猪体细胞克隆效率仍然很低,克隆猪平均出生率仅1%左右,新生克隆猪死亡率高达30%以上[78],严重制约着该技术在实际生产中的应用。SCNT主要包括卵母细胞体外成熟培养、去核、供体细胞培养、融合、重构胚胎培养和胚胎移植等操作,每一环节对克隆胚胎的发育都至关重要。随着高通量测序技术的发展,SCNT胚胎重编程机制逐步被揭示,诸多学者尝试改良SCNT操作程序或纠正异常表观遗传修饰来提高SCNT胚胎发育率,进一步提升克隆效率。本文围绕供体细胞、卵母细胞和克隆胚胎3个方面综述了提高猪SCNT胚胎发育率的研究,为提高猪体细胞克隆效率提供参考。
1 体细胞克隆胚胎发育异常原因
SCNT通过重编程体细胞基因组来产生全能基因组,重构胚胎能够发生类似于正常受精胚胎合子基因组激活(zygotic genome activation,ZGA)的过程。SCNT胚胎与正常受精胚胎存在遗传和表观遗传方面的差异。研究报道,SCNT发生的遗传异常是由于供体细胞和卵母细胞之间的细胞周期不同步或体外操作失误造成染色体畸变,如SCNT胚胎早期卵裂阶段经常出现的异常染色体分离[9]。表观遗传修饰异常被认为是SCNT胚胎发育的主要障碍,主要包括DNA高度甲基化、组蛋白的异常修饰以及X染色体失活异常。此外,卵母细胞成熟质量同样是影响体细胞克隆胚胎发育率的重要因素。
1.1 卵母细胞体外成熟质量差
卵母细胞体外成熟(in vitro maturation,IVM)培养是SCNT的关键步骤,其成熟质量对克隆胚胎发育至关重要。卵母细胞IVM培养的关键在于用IVM培养基模拟卵泡液微环境,使未成熟的卵母细胞同时达到细胞核和细胞质的成熟。但由于SCNT所需的大量卵母细胞来自于未性成熟的青年母猪卵巢,卵母细胞体外成熟效率普遍低于体内正常发育[10],因此,揭示卵母细胞成熟的分子机制与影响因素对改善卵母细胞的成熟质量至关重要。
Ye等[11]通过单细胞多组学分析发现,人体外与体内发育成熟的卵母细胞相比,存在507个差异表达基因,体内成熟的卵母细胞中富集的差异基因与细胞周期过程有关,而体外成熟的卵母细胞中富集的差异基因与线粒体呼吸生物合成有关。卵母细胞体外成熟培养过程相关信号调控通路受到影响将导致体外培养成熟效率和质量下降。体内与体外成熟卵母细胞的主要差异是线粒体功能,体外成熟的有限条件可导致线粒体形态改变以及编码与线粒体功能相关的基因表达改变,功能失调的线粒体抗氧化应激能力减弱,进而影响卵母细胞的成熟[12]。
1.2 供体细胞DNA 高度甲基化
DNA 甲基化是最早发现的表观修饰遗传标记,指DNA分子在DNA甲基转移酶的作用下将甲基选择性地添加到特定碱基上的过程。其中5-甲基胞嘧啶(5-methylcytosine,5mC)是最常见的DNA甲基化标记。供体细胞基因组DNA的高度甲基化是阻碍SCNT胚胎发育的主要因素之一,不同供体的细胞DNA甲基化存在明显差异,使用DNA甲基化水平低的供体细胞获得的SCNT胚胎具有更强的发育潜力[13]。罗婵[14]研究发现,水牛胎儿的4个成纤维细胞系中,克隆效率高的供体细胞DNA甲基化水平较低,赵鑫[15]的研究进一步证实了此结果。此外,Zhai等[16]研究发现,用猪骨髓间充质干细胞(bone marrow mesenchymal stemcell,BMSC)作为供体细胞的SCNT(BMSC nucleartransfer,BMSC-NT)胚胎比猪胎儿成纤维细胞(porcine fetal fibroblast,PFF) 的SCNT (PFFnuclear-transfer,FF-NT)胚胎具有更低的DNA 甲基化和5mC水平,更有利于SCNT胚胎的发育。
SCNT胚胎的DNA甲基化主要来源于供体细胞的细胞核,合子基因组激活后,供体细胞核基因的不完全去甲基化致使胚胎发育相关基因表达紊乱,进而导致胚胎发育阻滞[17]。供体细胞基因组DNA 高度甲基化是引起SCNT 胚胎发育过程中DNA甲基化水平动态变化的直接原因。Liu等[18]通过胚胎活检系统和单细胞简化亚硫酸氢盐测序(reduced representation bisulfite sequencing,RRBSseq)分析了SCNT胚胎的DNA甲基化水平发现,在2-细胞和4-细胞阶段,SCNT样本的甲基化水平普遍高于对应时期的体外受精胚胎(in vitrofertilization,IVF)[19],而SCNT与IVF囊胚的甲基化水平相似。此外,Canovas等[20]通过RNA测序并结合RRBS-seq对猪体内和体外胚胎分析发现,通过辅助生殖技术获得的胚胎甲基化水平显著高于体内发育胚胎。综上表明,体细胞克隆胚胎供体细胞基因组DNA甲基化的擦除和重新建立对其后续SCNT胚胎发育至关重要,与受精胚胎相似的DNA甲基化模式更有利于克隆胚胎的发育。
1.3 克隆胚胎表观修饰异常
克隆胚胎表观修饰异常是导致SCNT胚胎发育异常的最主要原因,包括组蛋白异常修饰和X染色体失活异常等。哺乳动物基因组组蛋白的N端存在多种修饰,其中最常见的是组蛋白甲基化和乙酰化修饰。组蛋白甲基化是调控早期胚胎发育的重要遗传标记,在胚胎发育的各个阶段均发挥着重要作用。正常猪体外受精胚胎组蛋白H3第4和第27位赖氨酸的三甲基化位点(H3K4me3和H3K27me3)参与调控ZGA 的发生和退出,而SCNT胚胎H3K4me3和H3K27me3大量富集则会导致异常的ZGA[21]。SCNT 胚胎H3K27me3 和H3K9me3的不完全去甲基化使胚胎发育相关基因的表达下降,将导致胚胎发育阻滞[17]。有研究分别通过RRBS-seq及单细胞测序等手段发现小鼠SCNT胚胎Kdm4d和Kdm5b(2种特异性去甲基化酶)的表达水平在胚胎发育中发挥重要作用,Kdm5b与胚胎4-细胞到8-细胞阶段的发育速率明显相关[18],Kdm4d可调控SCNT胚胎在2-细胞阶段基因组的正常激活[22]。
组蛋白乙酰化对基因转录激活具有重要的调控作用,与SCNT胚胎的体外成熟与发育能力密切相关。检测SCNT胚胎发现,异常的组蛋白乙酰化会导致胚胎2-细胞期的基因组异常激活[23]。与猪体外受精胚胎相比,SCNT胚胎1-细胞与2-细胞期组蛋白H3第18位和组蛋白H4第8位赖氨酸位点乙酰化(H3K18ac、H4K8ac)水平均显著降低[24]。与猪受精胚胎相比,猪SCNT胚胎在基因组激活后H3K27ac出现异常[25]。综上说明,组蛋白甲基化和乙酰化的动态修饰模式对于维持正常的胚胎发育至关重要。
此外,X染色体失活异常也是导致SCNT胚胎发育异常的重要表观遗传修饰障碍。SCNT胚胎X染色体失活异常不仅与印记基因Xist 的调控密切相关,也与DNA甲基化以及组蛋白修饰相关。张宁等[26]研究发现,发育异常的SCNT胚胎Xist 基因的DNA 甲基化水平普遍高于正常胚胎,且其DNA甲基化水平与囊胚率之间存在显著负相关。研究表明,Xist 基因的调控与母源组蛋白甲基化有关,H3K27me3可抑制母源Xist 基因在X染色体失活中发挥的作用,导致SCNT胚胎发育阻滞,而胚胎中X染色体失活通常伴随着H3K9甲基化的发生[2728]。另外,Xist 基因异常活化可导致胚胎致死或者受体母猪流产,而敲除供体细胞的Xist 基因或利用非编码RNA干扰胚胎中Xist 基因的异常表达均可显著提高猪SCNT胚胎发育率[2930]。
2 提高猪体细胞克隆胚胎发育率的方法
2.1 选择合适的供体细胞
在猪SCNT研究中,多种类型体细胞被用作核移植的供体细胞,至少有10种细胞已被证实了具有重编程能力,目前应用较广泛的是猪胎儿成纤维细胞和猪肾成纤维细胞[31]。不同类型供体细胞的克隆效率存在差异。Wei等[32]比较了用于SCNT的PFF、新生猪成纤维细胞和成年猪成纤维细胞的重编程能力,结果表明,PFF 和新生猪成纤维细胞SCNT胚胎的卵裂率、囊胚率和妊娠率均高于成年猪成纤维细胞。此外,Jiao等[33]比较了供体细胞大小对SCNT胚胎发育的影响,结果表明,小于13 μm体细胞的卵裂率显著高于13~18 μm,囊胚率和囊胚细胞数同样显著高于13 μm以上的体细胞,说明直径小于13 μm的PFF更有利于SCNT胚胎发育。
干细胞(stem cell,SC)是具有自我复制能力、可分化成多种功能细胞的多潜能细胞,在现代生物育种(干细胞育种)中展现出显著的应用优势,作为供体细胞可显著提高SCNT重编程效率。Lee等[34]比较了精原干细胞(spermatogonial stem cell,SSC)和PFFs的SCNT效率,发现SSC作为供体细胞的SCNT胚胎具有更高的囊胚率,是生产猪SCNT胚胎的理想供体细胞类型,用脱氨胆碱处理核供体SSC可进一步提高克隆胚胎的生产效率。另有研究发现,BMSC-NT胚胎具有更活跃的表观遗传标记和更少的抑制性表观遗传标记,胚胎卵裂率和囊胚率接近IVF胚胎,且显著高于FF-NT胚胎;进一步用RRBS-seq 发现,在BMSC-NT 胚胎中,NANOG 和POU5f1 基因启动子区域DNA 甲基化水平低于FF-NT[16]。另外,BMSC的RNA N6-甲基腺苷(m6A)修饰水平比猪胚胎成纤维细胞高,更有利于SCNT胚胎发育[35]。以上说明,供体细胞的初始表观遗传状态差异对SCNT胚胎重编程有一定影响,具有开放染色质状态的干细胞更有利于供体细胞核重编程,可促进SCNT胚胎的发育。
2.2 优化卵母细胞体外成熟体系、去核及激活方式
优质卵母细胞是SCNT的基础,IVM培养液是影响卵母细胞质量的关键因素。诸多研究表明,IVM培养液中添加促成熟因子可有效改善卵母细胞的成熟质量(表1)。
卵母细胞的去核是否完全是影响核移植效率关键因素之一。卵母细胞去核方法有多种,包括盲摘法、化学药物诱导去核法、DNA染色荧光去核法等,其中大部方法步骤繁琐且需要借助显微操作仪才能完成。相比以上方法,手工克隆技术(handmade cloning,HMC)不需要借助显微操作仪等昂贵设备,借助显微切割刀去除极体下面的少量胞质即可达到去核目的,无需用Hoechst 33342染色以及紫外光检测,避免了紫外线照射对胚胎发育产生的不利影响[48]。任子利等[49]研究表明,用脱羰秋水酰碱辅助去核,可提高去核率和克隆胚胎囊胚率。HMC得到的胚胎无透明带,在胚胎体外培养液(invitro culture,IVC)容易暴露于有毒物质中,且需要2个卵母细胞重构1个胚胎,需要大量的卵母细胞,同时会带来线粒体异质性问题。因此,Lee等[50]建立了一种改良手工克隆技术,用1枚卵母细胞构建1个重构胚来解决卵母细胞去核过程中卵母细胞损耗和线粒体异质性的问题。此外,得益于人工智能的快速发展,一种无标记机器人精准去核技术可减少SCNT去核过程中的细胞质丢失,去核成功率达95%,来自机器人去核卵母细胞的克隆胚胎卵裂率是盲摘法的2倍,并显著提高了克隆效率[51]。
供体细胞与去核卵母细胞融合后,需要进一步激活重构胚才能使其正常发育。当前应用于猪SCNT重构胚激活方法可分为物理激活、化学激活和联合激活3大类。目前应用最为普遍的是联合激活,与单纯化学激活和物理激活相比,联合激活对重构胚的激活效率更高。石俊松等[52]比较了2 种电激活方式(同步融合激活法和延迟激活法),发现延迟激活虽然会降低重构胚的融合率,但能够显著提高克隆胚胎的卵裂率和囊胚率。
2.3 添加小分子化合物促进重构胚发育
诸多研究人员使用表观遗传修饰相关小分子化合物用于处理SCNT 供体细胞,以改善异常DNA 甲基化提高胚胎发育率。研究发现,0.25~0.50 μmol·L−1的5-氮杂-2’脱氧胞苷(5-Aza-CdR)处理PFF 72 h,可提高猪克隆胚胎囊胚发育率[53];用5 μmol·L−1 非核苷类 DNA 甲基转移酶(DNAmethyltransferase,DNMT)抑制剂RG108 处理PFF72 h可改善其核重编程,降低PFF基因组的DNA甲基化的时间依赖性和印记基因 h19的甲基化水平,改善猪SCNT胚胎发育,提高SCNT效率[54]。谢颖瑜等[55]研究表明,0.1 mmol·L−1的S-腺苷高半胱氨酸处理猪成纤维细胞24 h,可降低DNMT转录水平和整体DNA甲基化水平,具有诱导成纤维细胞分化、改善克隆效率的潜能。
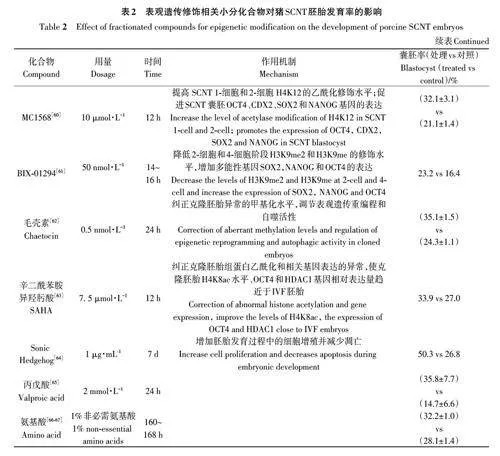
越来越多的研究表明,在IVC中添加表观遗传修饰小分化合物培养克隆胚胎,可纠正表观重编程错误,调控胚胎发育相关基因的表达,从而提高猪SCNT胚胎发育率(表2)。
3 展 望
近年来,随着高通量多组学测序技术(如DNA测序、转录谱、ChiIP-Seq等)在全基因组水平上的快速发展和广泛应用,克隆胚胎发育过程中的重编程障碍机制逐渐被发现。利用DNA甲基转移酶抑制剂、组蛋白去乙酰化酶抑制剂等改善表观重编程的策略,在提高克隆胚胎发育的研究中取得了较大进展,但克隆动物的出生率和健仔率依然很低。
在未来的研究中,提高SCNT 胚胎的发育率仍然是体细胞克隆研究的重点方向。随着研究人员对克隆胚胎发育机制的研究逐步加深,更多调控SCNT胚胎发育的特异性靶点逐渐被发现,有待开发出更多、更有效的特异性表观遗传修饰相关小分子化合物、基因特异性干扰RNA及其组合方法,联合最新测序技术和最新基因编辑技术,快速解析表观遗传重编程机制,克服SCNT胚胎发育过程中的重编程障碍,从而大幅提高克隆胚胎的发育率。当前微量细胞测序技术已应用于解析SCNT异常机制,小鼠SCNT胚胎发育率得到很大改善,囊胚率达80%以上[23]。猪SCNT可以借鉴相应技术,进一步改善猪体细胞克隆胚胎的发育率。
参考文献
[1] WILMUT I, SCHNIEKE A E, MCWHIR J, et al .. Viable offspring derived from fetal and adult mammalian cells [J].Nature, 1997,385: 810-813.
[2] LIU Z, CAI Y J, WANG Y, et al .. Cloning of macaque monkeys by somatic cell nuclear transfer [J]. Cell, 2018,172(4):881-887.
[3] PRATHER R S, SIMS M M, FIRST N L. Nuclear transplantation in early pig embryos [J]. Biol. Reprod., 1989,41(3):414-418.
[4] POLEJAEVA I A, CHEN S H, VAUGHT T D, et al .. Cloned pigs produced by nuclear transfer from adult somatic cells [J].Nature, 2000,407:86-90.
[5] RYU J, PRATHER R S, LEE K. Use of gene-editing technology to introduce targeted modifications in pigs [J/OL]. J.Anim. Sci. Biotechnol., 2018, 9:5 [2024-02-20]. https://doi.org/10.1186/s40104-017-0228-7.
[6] SPIZZO T, DENNER J, GAZDA L, et al .. First update of the International xenotransplantation association consensus statement on conditions for undertaking clinical trials of porcine islet products in type 1 diabetes--chapter 2a: source pigs--preventing xenozoonoses [J]. Xenotransplantation, 2016,23(1):25-31.
[7] ZHANG X L, GAO S R, LIU X Y. Advance in the role of epigenetic reprogramming in somatic cell nuclear transfermediated embryonic development [J/OL]. Stem Cells Int., 2021,2021:6681337 [2024-02-20]. https://doi.org/10.1155/2021/6681337.
[8] SCHMIDT M, WINTHER K D, SECHER J O, et al .. Postmortem findings in cloned and transgenic piglets dead before weaning [J].Theriogenology,2015,84(6):1014-1023.
[9] ATSUO O, SHOGO M, KIMIKO I. 25th anniversary of cloning by somatic-cell nuclear transfer: epigenetic abnormalities associated with somatic cell nuclear transfer [J]. Reproduction(Camb. Engl.), 2021,162(1):45-58.
[10] TELFER E E, ANDERSEN C Y. In vitro growth and maturation of primordial follicles and immature oocytes [J].Fert. Ster., 2021,115(5):1116-1125.
[11] YE M, YANG Z Y, ZHANG Y,et al .. Single-cell multiomic analysis of in vivo and in vitro matured human oocytes [J]. Hum.Reprod., 2020,35(4):886-900.
[12] KIRILLOVA A, SMITZ J, SUKHIKH G T, et al .. The role of mitochondria in oocyte maturation [J/OL]. Cells, 2021, 10(9):92484 [2024-02-20]. https://doi.org/10.3390/cells10092484.
[13] 邢青华,邹超霞,魏耀昶,等.不同年龄水牛成纤维细胞DNA甲基化与组蛋白乙酰化水平分析[J].中国畜牧杂志,2021,57(8):102-106.
[14] 罗婵. 不同个体水牛体细胞克隆效率差异的分子机制研究[D].南宁:广西大学,2018.
LUO C. Studies related to the mechanism of different somatic cell cloning efficiencies in different buffalo individuals [D].Nanning: Guangxi University, 2018.
[15] 赵鑫.水牛体细胞继代克隆及卵胞质对供体细胞核重编程作用的初步研究[D].南宁:广西大学,2019.
ZHAO X. A preliminary study of buffalo serial somatic cell nuclear transfer and the effect of oocyte cytoplasm on reprogramming of donor cell [D]. Nanning: Guangxi University,2019.
[16] ZHAI Y, LI W, ZHANG Z, et al .. Epigenetic states of donor cells significantly affect the development of somatic cell nuclear transfer (SCNT) embryos in pigs [J]. Mol. Reprod.Dev., 2018,85(1):26-37.
[17] 曹嘉程,薛丽娥,蓝群,等.表观遗传修饰影响哺乳动物体细胞核移植效率的研究进展[J]. 中国畜牧杂志,2023,59(3):7-15.
CAO J C, XUE L E, LAN Q, et al .. Research progress on the effect of epigenetic modification affecting the efficiency of mammalian somatic cell nuclear transfer [J]. Chin. J. Anim.Sci., 2023,59(3):7-15.
[18] LIU W Q, LIU X Y, WANG C F, et al .. Identification of key factors conquering developmental arrest of somatic cell cloned embryos by combining embryo biopsy and single-cell sequencing [J/OL]. Cell Discov., 2016, 2:16010 [2024-02-20].https://doi.org/10.1038/celldisc.2016.10.
[19] MATOBA S, WANG H H, JIANG L, et al .. Loss of H3K27me3 imprinting in somatic cell nuclear transfer embryos disrupts post-implantation development [J]. Cell Stem Cell, 2018,23(3):343-354.
[20] CANOVAS S, IVANOVA E, ROMAR R, et al .. DNA methylation and gene expression changes derived from assisted reproductive technologies can be decreased by reproductive fluids [J/OL]. Elife, 2017, 6:23670 [2024-02-20]. https://doi.org/10.7554/eLife.23670.
[21] BU G, ZHU W, LIU X, et al .. Coordination of zygotic genome activation entry and exit by H3K4me3 and H3K27me3 in porcine early embryos [J]. Genome Res., 2022, 32(8): 1487-1501.
[22] HIROSE M, HADA M, KAMIMURA S, et al .. Aberrant imprinting in mouse trophoblast stem cells established from somatic cell nuclear transfer-derived embryos [J]. Epigenetics,2018,13(7):693-703.
[23] YANG G, ZHANG L F, LIU W Q, et al .. Dux-mediated corrections of aberrant H3K9ac during 2-cell genome activation optimize efficiency of somatic cell nuclear transfer [J]. Cell Stem Cell, 2021,28(1):150-163.
[24] YAMANAKA K, SUGIMURA S, WAKAI T, et al .. Acetylation level of histone H3 in early embryonic stages affects subsequent development of miniature pig somatic cell nuclear transfer embryos [J]. J. Reprod. Dev., 2009,55(6):638-644.
[25] ZHOU N R, CAO Z B, WU R H, et al .. Dynamic changes of histone H3 lysine 27 acetylation in pre-implantational pig embryos derived from somatic cell nuclear transfer [J]. Anim.Reprod. Sci., 2014,148(3/4):153-163.
[26] 张宁,余波,石俊松,等.印记基因XIST和H19DNA甲基化水平与猪克隆胚胎发育效率关联分析[J].华南农业大学学报,2021,42(3):9-16.
ZHANG N, YU B, SHI J S, et al .. Correlation between DNA methylation levels of imprinted genes XIST and H19 and developmental efficiency of porcine cloned embryo [J]. J. South China Agric. Univ., 2021,42(3):9-16.
[27] CHEN Z Y, ZHANG Y. Maternal H3K27me3-dependent autosomal and X chromosome imprinting [J]. Nat. Rev. Genet.,2020,21:555-571.
[28] FURLAN G, GALUPA R. Mechanisms of Choice in XChromosome Inactivation [J/OL]. Cells, 2022, 11(3):535 [2024-02-20]. https://doi.org/10.3390/cells11030535.
[29] RUAN D G, PENG J Y, WANG X S, et al .. XIST derepression in active X chromosome hinders pig somatic cell nuclear transfer [J].Stem Cell Rep., 2018,10(2):494-508.
[30] YANG X, WU X, YANG Y, et al .. Improvement of developmental competence of cloned male pig embryos by short hairpin ribonucleic acid (shRNA) vector-based but not small interfering RNA (siRNA) -mediated RNA interference (RNAi) of Xist expression [J]. J. Reprod. Dev., 2019, 65(6):533-539.
[31] LIU H B, LV P R, ZHU X X, et al .. In vitro development of porcine transgenic nuclear-transferred embryos derived from newborn Guangxi Bama mini-pig kidney fibroblasts [J]. Vitro Cell. Dev. Biol. Anim., 2014,50(9):811-821.
[32] WEI H J, QING Y B, PAN W R, et al.. Comparison of the efficiency of Banna miniature inbred pig somatic cell nuclear transfer among different donor cells [J/OL]. PLoS One, 2013, 8(2):e57728 [2024-02-20]. https://doi.org/10.1371/journal.pone.0057728.
[33] JIAO D, CHENG W, ZHANG X, et al .. Improving porcine SCNT efficiency by selecting donor cells size [J].Cell Cycle,2021,20(21):2264-2277.
[34] LEE J, LEE Y, LEE G S,et al .. Comparative study of the developmental competence of cloned pig embryos derived from spermatogonial stem cells and fetal fibroblasts [J]. Reprod.Domest. Anim., 2019,54(9):1258-1264.
[35] ZHANG M, ZHAI Y H, AN X L, et al .. DNA methylation regulates RNA m(6) A modification through transcription factor SP1 during the development of porcine somatic cell nuclear transfer embryos [J/OL]. Cell Prolif, 2023: e13581 [2024-02-20]. https://doi.org/10.1111/cpr.13581.
[36] 高悦悦,顾浩,陈矾,等.α-硫辛酸对猪卵母细胞体外成熟及孤雌胚胎发育的影响[J]. 中国畜牧杂志, 2023, 59(11):204-209.
[37] 刘子嶷,李凯扬,王鹏,等.添加IGF-1对体外培养不同直径猪卵泡中卵母细胞成熟率的影响[J]. 中国畜牧杂志,2022,58(11):189-194.
[38] NIE J Y, YAN K, SUI L M, et al .. Mogroside V improves porcine oocyte in vitro maturation and subsequent embryonic development [J]. Theriogenology, 2020,141:35-40.
[39] 张碧菡,赵宝宝,高静,等.甘草酸单铵盐对猪卵母细胞体外成熟及胚胎发育能力的影响[J].中国兽医学报,2023,43(11):2361-2367.
ZHANG B H, ZHAO B B, GAO J, et al .. Effects of monoammonium glycyrrhizinate on oocyte maturation and embryonic development in porcine [J]. Chin. J. Vet. Sci., 2023,43(11):2361-2367.
[40] YANG L, WANG Q K, CUI M S, et al .. Effect of melatonin on the in vitro maturation of porcine oocytes, development of parthenogenetically activated embryos, and expression of genes related to the oocyte developmental capability [J/OL]. Animals (Basel), 2020, 10(2):209 [2024-02-20]. https://doi.org/10.3390/ani10020209.
[41] 荆焕松,周荣,罗绿花,等.猪卵母细胞体外成熟过程中添加铜蓝蛋白对成熟率和孤雌胚胎发育能力的影响[J].广东畜牧兽医科技,2023,48(6):28-33.
JING H S, ZHOU R, LUO L H, et al .. Effects of ceruloplasmin on maturation rate and embryonic development of porcine oocytes cultured in vitro [J]. Guangdong J. Anim. Vet. Sci.,2023,48(6):28-33.
[42] 蓝昕蕊,赵宝宝,张碧菡,等.β-谷甾醇对猪卵母细胞体外成熟和胚胎发育的影响[J]. 畜牧兽医学报,2024,55(4):1629-1637.
LAN X R, ZHAO B B, ZHANG B H, et al .. Effects of β -Sitosterol on porcine oocyte maturation and embryonic development in vitro [J]. Acta Vet. Zootechnica Sin., 2024,55(4):1629-1637.
[43] HICKS E, MENTLER M, ARENA H A, et al .. Cyanidin improves oocyte maturation and the in vitro production of pig embryos [J].Vitro Cell. Dev. Biol. Anim., 2020,56(7):577-584.
[44] 许建春,汪浩鑫,自永宏,等.虾青素对猪卵母细胞体外成熟的影响[J].中国兽医学报,2022,42(12):2549-2554.
XU J C, WANG H X, ZI Y H,et al ..Effects of astaxanthin on maturation of porcine oocytes in vitro [J]. Chin. J. Vet. Sci.,2022,42(12):2549-2554.
[45] 孙婧陶,姜超前,刘佳慧,等.单宁酸对猪卵母细胞体外成熟及胚胎发育能力的影响[J].中国畜牧兽医,2021,48(10):3708-3714.
SUN J T, JIANG C Q, LIU J H, et al.. Effects of tannin acid on the in vitro maturation and embryonic development of porcine oocytes[J]. China Anim. Husb. Vet. Med., 2021,48(10):3708-3714.
[46] WANG X, ZHU X, LIANG X, et al .. Effects of resveratrol on in vitro maturation of porcine oocytes and subsequent early embryonic development following somatic cell nuclear transfer [J]. Reprod. Domest. Anim., 2019,54(9):1195-1205.
[47] ZHOU C, ZHANG X, SHIYANG X,et al .. Tea polyphenol protects against cisplatin-induced meiotic defects in porcine oocytes [J]. Aging, 2019,11(13):4706-4719.
[48] LIN Q Y, AIHARA M, SHIRAI A,et al .. Porcine embryo development and inactivation of microorganisms after ultraviolet-C irradiation at 228 nm [J]. Theriogenology, 2023,197:252-258.
[49] 任子利,赵彦玲,杨小淦,等.猪卵母细胞化学辅助手工去核及手工核移植胚胎发育能力因素的研究[J].畜牧兽医学报,2011,42(7):921-931.
REN Z L, ZHAO Y L, YANG X G,et al .. Studies on factors affecting chemical assisted handmade enucleation of porcine oocytes and developmental capacities of handmade somatic cells nuclear transfer embryos [J]. Acta Vet. Zootechnica Sin.,2011,42(7):921-931.
[50] LEE E J, JI K B, LEE J H, et al .. Application of the modified handmade cloning technique to pigs [J]. J. Anim. Sci. Technol.,2021,63(2):281-294.
[51] ZHAO Q, QIU J, FENG Z,et al .. Robotic label-free precise oocyte enucleation for improving developmental competence of cloned embryos [J]. IEEE Trans. Bio Med. Eng., 2021,68(8):2348-2359.
[52] 石俊松,罗绿花,周荣,等.延迟激活对猪克隆胚胎体外、体内发育效率的影响[J].中国生物工程杂志,2019,39(4):16-23.
SHI J S, LUO L H, ZHOU R, et al .. Delayed activation can improve in vitro and in vivo developmental capacity of pig cloned embryos [J]. China Biotechnol., 2019,39(4):16-23.
[53] 吕玲燕,陆杏蓉,孙俊铭,等. 5-Aza-CdR对德保黑猪手工克隆重构胚胎体外发育效果的影响[J]. 中国畜牧兽医,2018,45(11):3144-3152.
LYU L Y, LU X R, SUN J M, et al .. Effect of 5-Aza-CdR on the in vitro developmental potency of reconstructed embryos of Debao black pig handmade cloner [J]. China Anim. Husb. Vet.Med., 2018,45(11):3144-3152
[54] WU C F, ZHANG D F, ZHANG S, et al .. Optimizing treatment of DNA methyltransferase inhibitor RG108 on porcine fibroblasts for somatic cell nuclear transfer [J]. Reprod.Domest. Anim., 2019,54(12):1604-1611.
[55] 谢颖瑜,曹嘉程,宋红兵,等.SAH对猪成纤维细胞体外增殖及甲基化水平的影响[J].中国畜牧杂志,2022,58(8):87-93.
[56] JIN L, GUO Q, ZHU H Y, et al .. Quisinostat treatment improves histone acetylation and developmental competence of porcine somatic cell nuclear transfer embryos [J]. Mol. Reprod.Dev., 2017,84(4):340-346.
[57] 朱彦宾,龙川,冯冲,等.Scriptaid处理可提高近交系五指山猪克隆胚胎的发育能力和克隆效率[J].基因组学与应用生物学,2011,30(3):268-273.
ZHU Y B, LONG C, FENG C, et al .. Improving the developmental potency of cloned embryos and cloning efficiency in Wuzhishan inbred pig by scriptaid treatment [J].Genom. Appl. Biol., 2011,30(3):268-273.
[58] JIN L, ZHU H Y, GUO Q, et al .. Effect of histone acetylation modification with MGCD0103,a histone deacetylase inhibitor,on nuclear reprogramming and the developmental competence of porcine somatic cell nuclear transfer embryos [J]. Theriogenology,2017,87:298-305.
[59] 侯黎明.组蛋白去乙酰化酶抑制剂Oxamflatin对猪体细胞克隆胚胎体外发育率的影响[D].武汉:华中农业大学,2016.
HOU L M. The effect of histone deacetylase inhibitor Oxamflatin in vitro developmental efficiency of porcine somatic cell nuclear transfer embryos [D]. Wuhan: Huazhong Agricultural University, 2016.
[60] WANG H, CUI W, MENG C, et al .. MC1568 enhances histone acetylation during oocyte meiosis and improves development of somatic cell nuclear transfer embryos in pig [J]. Cell.Reprogramming, 2018,20(1):55-65.
[61] HUANG J, ZHANG H, YAO J, et al .. BIX-01294 increases pig cloning efficiency by improving epigenetic reprogramming of somatic cell nuclei [J]. Reproduction, 2016,151(1):39-49.
[62] JEONG P S, SIM B W, PARK S H, et al .. Chaetocin improves pig cloning efficiency by enhancing epigenetic reprogramming and autophagic activity [J/OL]. Int. J. Mol. Sci., 2020,21(14):E4836 [2024-02-20]. https://doi.org/10.3390/ijms21144836.
[63] 孙俊铭,崔奎青,李志鹏,等. SAHA处理对猪手工克隆胚胎体外发育潜能的影响[J].黑龙江畜牧兽医, 2018(5):104-108.
SUN J M, CUI K Q, LI Z P, et al .. Effects of SAHA on the developmental potential of porcine handmade cloned embryos in vitro [J]. Heilongjiang Anim. Sci. Vet. Med., 2018(5):104-108.
[64] NGUYEN N T, LIN D P C, SIRIBOON C, et al .. Sonic Hedgehog improves in vitro development of porcine parthenotes and handmade cloned embryos [J].Theriogenology, 2010,74(7):1149-1160.
[65] 罗绿花,周荣,麦然标,等.丙戊酸处理对猪体细胞克隆胚胎体外、体内发育效率的影响[J].黑龙江畜牧兽医, 2021(5):64-68.
LUO L H, ZHOU R, MAI R B, et al .. Effects of valproic acid treatment on developmental capacity of porcine cloned embroys in vitro and in vivo [J]. Heilongjiang Anim. Sci. Vet.Med., 2021(5):64-68.
[66] 吕玲燕,陈宝剑,吴永绍,等.氨基酸对猪手工克隆重构胚胎发育效果的影响[J].湖北农业科学,2018,57(6):108-110.
LYU L Y,CHEN B J,WU Y S,et al ..Effects of developmental potency of reconstructed embryos on pig handmade clone of amino acid [J]. Hubei Agric. Sci., 2018,57(6):108-110.
[67] LIU T, LI J, DOU H, et al .. Low-concentration essential amino acids in PZM-3 improve the developmental competence of porcine embryos produced by handmade cloning [J]. Cell.Reprogramming, 2020,22(6):282-290.
