红锥树干径向水分亏缺对饱和水汽压差的时滞响应
2024-11-11刘士玲杨保国郑路张培庞圣江陈琳闵惠琳舒韦维田祖为黄荣
摘 要:【目的】探索作为树干水分变化状况指标的树木水分亏缺(TWD)变化特征及其与饱和水汽压差(VPD)的关系,揭示径向变化对环境水分胁迫的响应特征,为合理指导红锥人工林经营提供科学依据和数据支持。【方法】利用径向变化记录仪记录不同分化等级红锥树干径向变化过程,同步监测环境因子,分析TWD对VPD的响应特征。【结果】不同分化等级红锥的TWD波动趋势相似,TWD随着VPD的变化而变化,但并不同步。小时尺度上,TWD和VPD显著正相关(r=0.488,r=0.489,r =0.512,P<0.01),划分失水期和复水期后,两个时期的TWD和VPD也显著正相关,且失水期的相关性增强(r=0.623,r=0.667,r=0.593,P<0.01)。日尺度上,无论是否划分失水期和复水期,二者均显著正相关,但相关性减弱。前后1天内的时滞响应分析结果显示,无论分化等级,TWD与VPD显著正相关(P<0.01);划分失水期和复水期后,在任何时间段考虑时滞响应时也均达到显著正相关,失水期的相关性强于复水期的相关性。未划分失水期和复水期时,不同分化等级红锥TWD对VPD响应滞后时间均为3 h。划分失水期和复水期后,失水期TWD对VPD响应滞后时间缩短,复水期TWD对VPD响应滞后时间延长。优势木、中等木和被压木复水期的滞后时间比失水期分别长3、4和5 h。VPD大幅度波动条件(VPDL)下,随着VPD的反复升降,TWD表现为失水-复水交替出现,变化的总体趋势是上升的;VPD小幅度波动条件(VPDs)下,TWD变化趋势较平缓,两种条件下TWD对VPD的响应均存在时滞。无论分化等级,VPDL条件下TWD与VPD之间的相关性(r=0.550,r=0.553,r=0.584,P<0.01)均强于VPDs下的相关性(r=0.256,r=0.260,r=0.281,P<0.01)。VPDL条件下,考虑滞后时间后,无论是否划分失水期和复水期,不同分化等级红锥TWD与VPD之间的相关性均增强。VPDs条件下,复水期的相关性低于忽略时滞效应时的相关性。【结论】TWD对VPD的响应存在时滞效应,在失水期和复水期,TWD对VPD的响应特征不同,且在不同分化等级间存在差异;失水期和复水期的划分有助于研究TWD与VPD的相关性。
关键词:径向变化动态;径向变化记录仪;红锥;时滞效应;林木分化等级
中图分类号:S718.43 文献标志码:A 文章编号:1673-923X(2024)09-0094-11
基金项目:中央级公益性科研院所基本科研业务费专项(CAFYBB2021MA002);广西自然科学基金(2019GXNSFBA245067)。
Time lag of stem water deficit in response to vapor pressure deficit of Castanopsis hystrix
LIU Shiling1,2,3, YANG Baoguo1, ZHENG Lu1,2,3, ZHANG Pei1, PANG Shengjiang1, CHEN Lin1,2,3, MIN Huilin1,2,3, SHU Weiwei1,2,3, TIAN Zuwei1, HUANG Rong1
(1. Experimental Center of Tropical Forestry, Chinese Academy of Forestry, Chongzuo 532600, Guangxi, China; 2. Guangxi Youyiguan Forest Ecosystem National Observation and Research Station, Chongzuo 532600, Guangxi, China; 3. Youyiguan Forest Ecosystem Observation and Research Station of Guangxi, Chongzuo 532600, Guangxi, China)
Abstract:【Objective】Tree water deficit-induced stem shrinkage (TWD) is an indicator for tree water status. By studying the variation characteristics of TWD and its relationship with VPD, and revealing the response characteristics of radial change to environmental water stress, the scientific basis and data support for reasonable guidance of Castanopsis hystrix plantation management was provided.【Method】Dendrometer was used to continuously record the radial growth change process of C. hystrix, and the environmental factors were measured simultaneously. The main goal of this study was to analyze the response characteristics of TWD to VPD.【Result】The TWD fluctuation trend of different differentiation classes was similar, and TWD varied with VPD, but not synchronously. On the hourly scale, TWD and VPD were significantly positively correlated (r = 0.488, r = 0.489, r = 0.512, P<0.01), and TWD and VPD were also significantly positively correlated after the water loss period and water recover period were divided. The correlation of water loss period was enhanced (r = 0.623, r = 0.667, r = 0.593, P<0.01). On the daily scale, no matter whether water loss period and water recovery period were divided, the two were significantly positively correlated, but the correlation was weakened. The results of comparison of pearson’s correlation coefficients between TWD and VPD under the condition of considering the time delay effect within 1-day showed that TWD and VPD were significantly positively correlated (P<0.01) regardless of differentiation classes. After dividing the water loss period and the water recovery period, the correlation of the water loss period was stronger than that of the water recovery period. When the water loss period and the water recovery period were not divided, the variation of TWD lagged behind that of VPD to a certain extent, by about 3 h. After the water loss period and the water recovery period were divided, the time lag of TWD to VPD in the water loss period was shortened, and time lag of TWD to VPD in the water recovery period was lengthened. The time lag of dominant, intermediate, and suppressed trees during water recovery period was 3 h, 4 h and 5 h, respectively, longer than that during the water loss period. Under VPD large fluctuation condition (VPDL), with repeatedly rising and falling of VPD, TWD appeared as water loss and recovery alternately. Under VPD small fluctuation condition (VPDs), TWD changed slowly. The time lag of TWD to VPD was existed under both conditions. The correlation between TWD and VPD under VPDL was stronger (r = 0.550, r = 0.553, r = 0.584, P<0.01) than that under VPDs (r = 0.256, r = 0.260, r = 0.281, P<0.01), regardless of differentiation classes. Under VPDL, after considering the time lag, the correlation between TWD and VPD of different differentiation classes was enhanced regardless of whether the water loss period and the water recovery period were divided. Under VPDs, the correlation of water recovery period was lower than that when the time lag effect was ignored.【Conclusion】TWD has a time-lag effect in response to VPD. TWD has different response characteristics to VPD in the water loss period and water recovery period, and these relationships have differences among different tree differentiation classes. The division of water loss period and water recovery period is helpful to study the correlation between TWD and VPD.
Keywords: radial variation dynamics; dendrometer; Castanopsis hystrix; time-lag effect; tree differentiation classes
水是植物生长发育必不可少的要素,生长季节适量的水分供应是保证树木健壮生长的关键[1]。饱和水汽压差(VPD)是全球水资源和植物水分关系的一个主要决定因素[2]。21世纪以来,所有气候带的大气干旱化(以VPD为代表)都有所增加[3]。高VPD增加了土壤的水分损失率,使地表干燥,并导致更频繁和严重的干旱事件和植物水分胁迫[4]。水力结构功能维持是植物在干旱条件下存活的基础,水势是体现植物水分状况的直接指标[5],但水势指标测定的难点在于对高大树木树冠叶子的获取。近年来研究发现,树干径向变化是树木水分状况变化的一个潜在指标[6],对了解树木生长和生理特性应对气候变化具有重要意义。为阐明树木水分状况的动态变化特征及其与VPD的关系,对树干径向进行连续监测与分析是至关重要的。
树干径向变化的高分辨率时间监测可以提供不同时间尺度下的径向生长模式分析,以及揭示由水分变化引起的可逆径向变化。Zweifel[7]将由树干水分状况引起的可逆径向变化称为树木水分亏缺(tree water deficit-induced stem shrinkage,TWD),并假设树木在树干径向可逆变化期间零增长。通过研究53个站点收集的21个树种的年际径向生长数据发现,2018年欧洲热浪导致了广泛的树干收缩,针叶树和阔叶树径向水分亏缺不同[8]。对7种温带树种的径向生长研究发现,径向生长受高VPD的严重限制,而低VPD条件下即使在中等干燥的土壤中也能生长[9]。VPD对径向变化的相对影响仍然不清楚,并且可能根据物种和环境条件而变化。研究表明,从土壤-植物-大气连续体的整体来看,植物对环境水分的响应存在时滞现象[10]。对兴安落叶松Larix gmelinii的TWD与VPD关系研究发现,二者存在明显的滞后效应[11]。另有研究发现,不同分化等级个体间对气候敏感性的差异比物种间的差异更大[12],树的形态特征(如树高等)与树干径向的变化具有密切关系[13]。然而,以往研究鲜有考虑不同分化等级个体TWD对VPD的敏感性差异,特别是不同时期(失水期和复水期)和不同水分条件下TWD与VPD的响应特征。
红锥Castanopsis hystrix是我国亚热带地区的重要速生阔叶用材树种[14],是广东省种植面积最大(5.33万hm2以上)的珍贵树种,且种植面积仍在逐年增加[15]。红锥已被广东、广西、福建等省区列为重点发展的珍贵树种,也是广西建设全国木材战略核心储备基地和全国亚热带珍贵树种基地的主要造林珍贵树种[16-17]。该研究以我国桂西南热带林业实验中心2001年种植的红锥人工林为研究对象,通过对树干径向动态变化及相关的气象资料进行连续监测,分析不同时期和不同水分条件下TWD与VPD的相关性,以揭示不同分化等级间的时滞效应差异,确定径向变化对环境水分胁迫的响应特征,为合理指导红锥人工林经营提供科学依据和数据支持。
1 研究区概况与研究方法
1.1 研究区概况
研究区位于广西凭祥市友谊关森林生态系统国家定位观测研究站(21°57′~22°16′N,106°41′~106°59′E),属亚热带季风气候。据1965—2015年凭祥市气象局的观测资料,年均气温20.5~21.7 ℃,年降水量1 200~1 500 mm,全年降水量的78%发生在5—9月,年均相对湿度80%~84%。地带性土壤为红壤,pH值为4.5~7.5,土层厚度≥60 cm,腐殖质层厚度5~10 cm。
1.2 树木选择和生长监测
红锥人工林为2001年种植,现存林分密度为750株·hm-2。2017年底,设置30 m×30 m的红锥样地。采取定性和定量相结合的方法,在林分内选取优势木、中等木和被压木各4株[18](表1),在胸高1.3 m处安装树干径向生长变化记录仪(Ecomatik,Germany,精度±2 μm),连续监测径向变化动态。数据记录时间为每30 min记录1次,本研究涵盖的数据为2022年4—9月的监测值。
1.3 环境因子观测
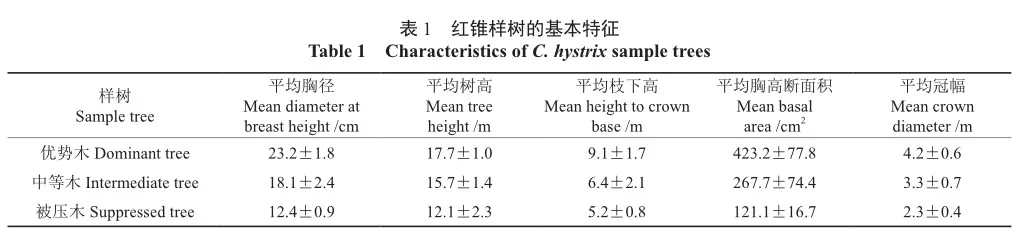
生长监测期间的气象资料如平均气温(Tave,℃)、降水量(P,mm)、10 cm深土壤体积含水率(SWC10)和10 cm深土壤温度(TS10)等从广西友谊关森林生态系统定位观测研究站获得。该站依据国家气象站标准和要求设有标准气象观测场(25 m×25 m),气象观测场距红锥试验林<1.5 km。数据记录时间为每1 min记录1次,饱和水汽压差(VPD,kPa)根据Murray等[19]的方法计算。
研究期间降水总量为845.8 mm,主要集中在5—9月;日均气温为11.4~34.9 ℃,平均为25.4 ℃;10 cm深土壤体积含水量为0.163~0.382 m3·m-3,平均为0.227 m3·m-3;饱和水汽压差为0~3.2 kPa,平均为0.76 kPa(图1)。
1.4 数据处理


将整个研究期观测到的时间序列数据分为失水期(TWDi)和复水期(TWDd),前者为TWD增加的时段,后者为TWD减小的时段。所有气象数据和径向数据换算为1 h的值。如果TWD值在1 h后下降,则记录为TWDd,否则记录为TWDi。
本研究采用Pearson相关分析和错位对比法,分析TWD与VPD变化的错位相关性。以步长为1 h,进行小时时间窗口下TWD与VPD逐行错位分析,分析提前或滞后24 h的时滞效应,两列数据错位后相关性最高且达到显著性水平时,所对应的时间即提前/滞后时间。采用SPSS19.0和SigmaPlot10.0软件对数据进行统计分析和图形绘制。
2 结果与分析
2.1 TWD和VPD的变化特征
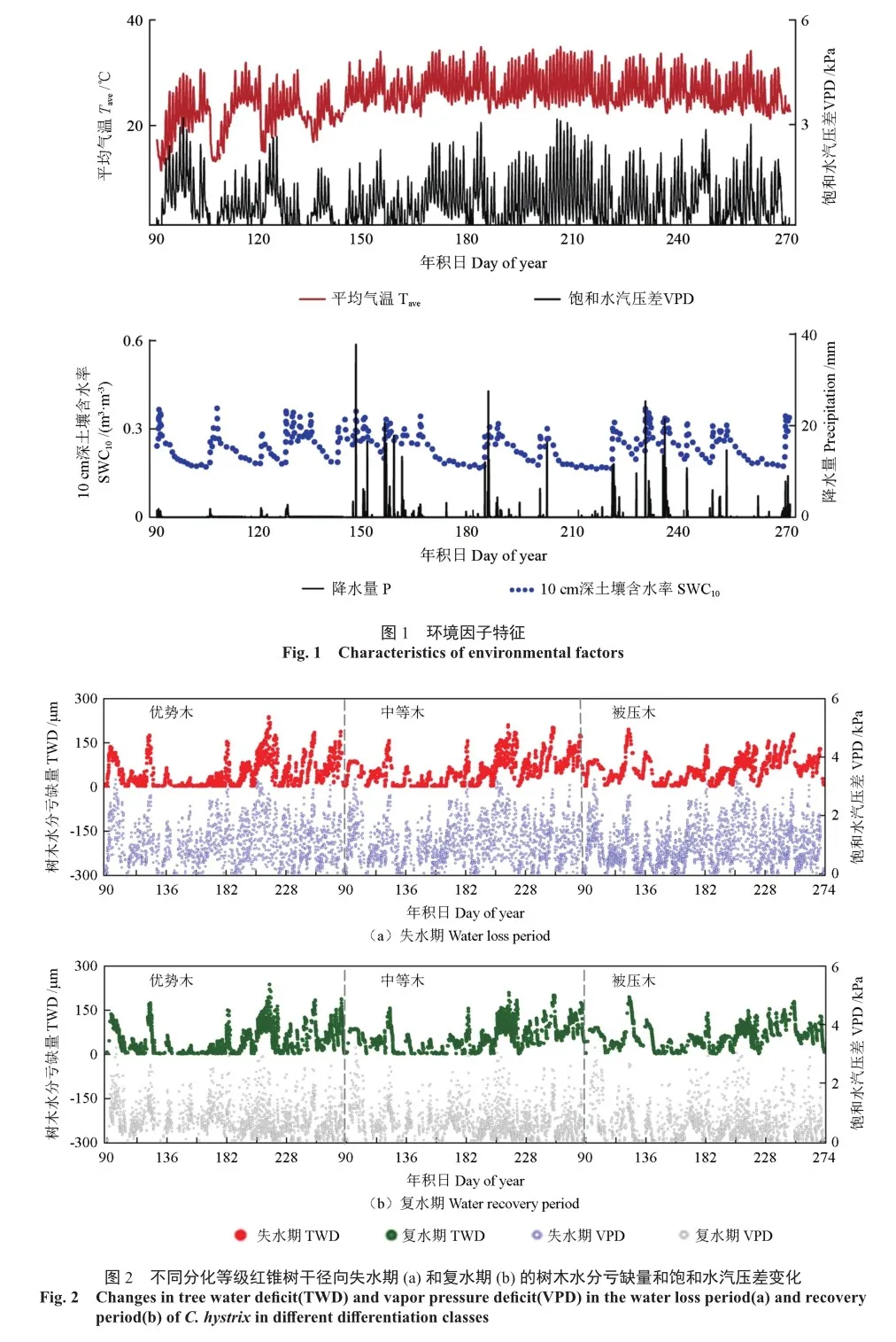
从整体表现来看,不同分化等级红锥的TWD波动趋势相似,除被压木外,TWD在失水期变化快,持续时间短;在复水期变化缓慢,持续时间长,TWDi和TWDd交替出现(图2,表2)。TWDi的变化时长和累积变化量表现为被压木>中等木>优势木,被压木的TWDd变化时长和累积变化量低于其他两个分化等级,优势木的TWDi发生时长最短,复水频率更高(表2)。不同月份间比较,5—6月份TWDi和TWDd大于0的发生次数少,其变化持续时间较短,振幅较小,TWDi和TWDd的累积变化量低于其他月份(图2)。失水期和复水期的VPD变化也表现不同。在失水期,VPD变化较大(图2a);在复水期,VPD变化较小(图2b)。不同月份间比较,5—6月份VPD均值小于其他月份(图2)。在整个观测期内,TWD随着VPD的变化而变化,但并不同步。

2.2 TWD对VPD变化的即时及滞后效应响应
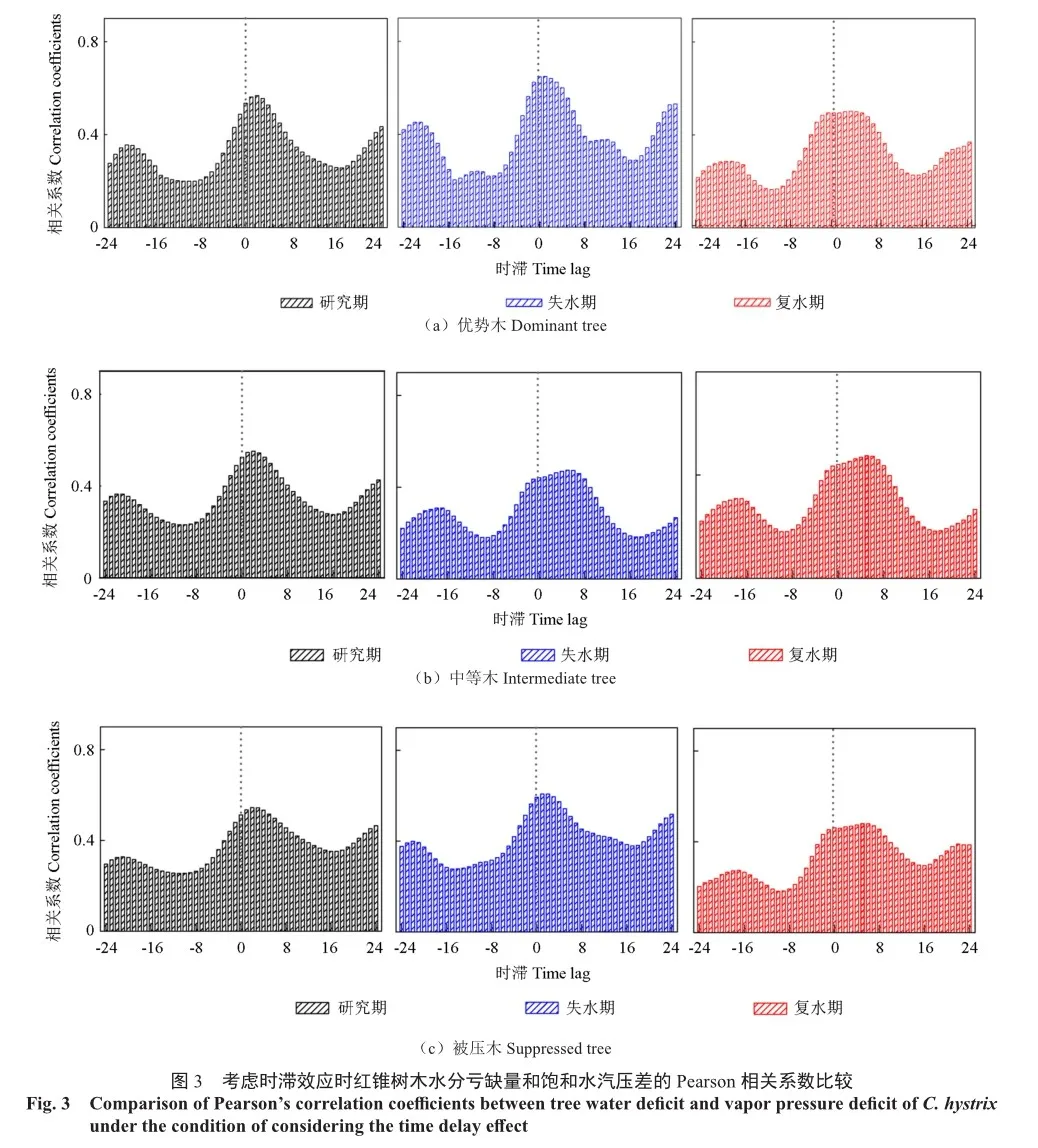
无论分化等级,在小时尺度上,未划分失水期和复水期时,TWD和VPD显著正相关(r=0.488,r=0.489,r =0.512,P<0.01)(表3)。划分失水期和复水期后,两个时期的TWD和VPD仍显著正相关,与未划分失水期和复水期相比,失水期TWD与VPD的相关性增强(r=0.623,r=0.667,r=0.593,P<0.01),且强于复水期TWD与VPD的相关性(r=0.490,r=0.435,r=0.461,P<0.01)(表3)。日尺度上,无论是否划分失水期和复水期,二者仍显著正相关,但相关性减弱(表3)。

前后1天内的时滞响应分析结果显示,不同分化等级红锥TWD对VPD的响应较一致(图3,表4)。未划分失水期和复水期时,TWD与VPD显著正相关(P<0.01)。划分失水期和复水期后,在任何时间段考虑时滞响应时也均达到显著正相关,失水期TWD与VPD的相关性强于复水期TWD与VPD的相关性。
TWD对VPD响应的滞后时间在失水期和复水期存在差异。不同分化等级红锥复水期TWD对VPD的响应滞后时间均大于失水期,优势木、中等木和被压木的复水期TWD对VPD的响应滞后时间分别比失水期长3、4和5 h。整个研究期,不同分化等级红锥TWD对VPD响应的滞后时间均为3 h。与未划分失水期和复水期相比,复水期TWD对VPD的反应时滞延长,而失水期TWD对 VPD的反应时滞缩短。
2.3 径向收缩随VPD变化的两种趋势特征
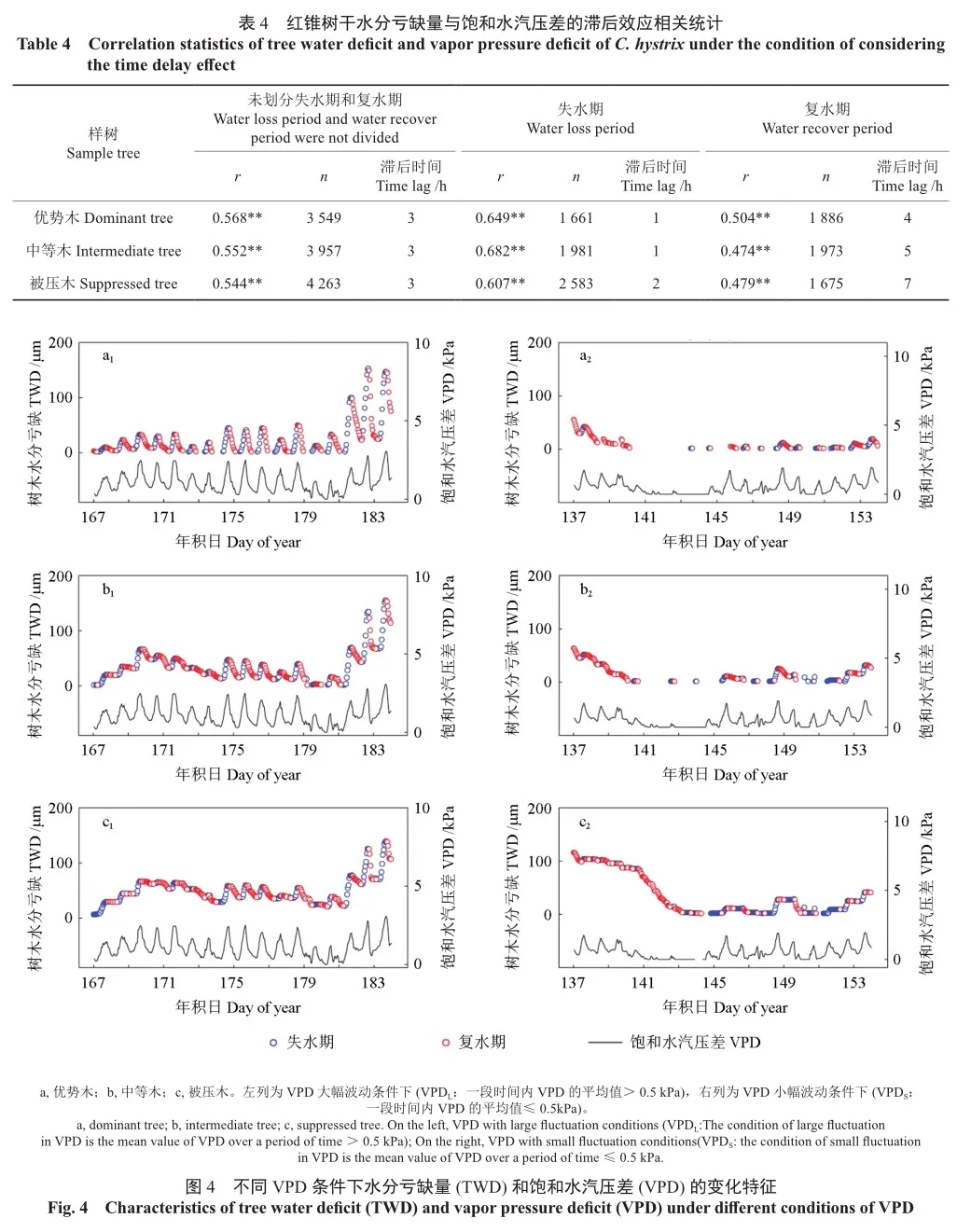
2个典型周期(VPD大/小幅度波动条件下)TWD随VPD的变化如图4所示,在不同条件下,TWD呈现出两种变化趋势。总体上,TWD随VPD的变化而变化,二者呈正相关,且TWD在变化趋势上滞后于VPD。
VPD大幅度波动条件(VPDL)下,TWD与VPD表现出相似的特征和变化趋势。随着VPD的反复升降,TWD表现为失水-复水交替出现,变化的总体趋势是上升的,但存在响应时滞。在VPD小幅度波动条件(VPDs)下,TWD变化趋势较平缓,TWD对VPD的响应也存在时滞。不同分化等级红锥TWD波动趋势较为一致,但VPDL条件下,优势木TWD变化表现为失水与复水交替最为明显,而在VPDs条件下,优势木TWD变化最为平缓。无论VPDL或VPDs条件下, TWDi和TWDd的变化时长和累积变化量均表现为被压木>中等木>优势木。
不同分化等级红锥TWD与VPD的相关性有所不同,表现为被压木>平均木>优势木。无论分化等级,VPDL条件下TWD与VPD之间的相关性(r=0.550,r=0.553,r=0.584,P<0.01)均强于VPDs下的相关性(r=0.256,r=0.260,r=0.281,P<0.01)(表5)。VPDL条件下,考虑滞后时间后,不同分化等级红锥TWD与VPD之间的相关性均增强(r=0.620,r=0.688,r=0.675,P<0.01);划分失水期和复水期后,两个时期的相关性也均增强。VPDs条件下,考虑滞后时间后,不同分化等级红锥TWD与VPD之间的相关性增强(r=0.335,r=0.352,r=0.322,P<0.01),失水期二者的相关性也增强,但复水期二者的相关性低于忽略时滞效应时的相关性。
3 讨 论
以往研究报道了干旱对亚热带树种生长的显著影响[21],强调水分对树木生长起着至关重要的作用,水分胁迫会限制我国亚热带地区树木的生长。本研究中,不同分化等级红锥在生长季节经历了一定的水分胁迫,优势木的TWDi发生时长较短,复水频率更高,这是由于TWD与树干弹性树皮细胞(形成层、韧皮部和薄壁组织细胞)中水的移动和储存有关[22-23]。一方面,优势木具有较强的树干储水能力[24],其对环境因子改变的缓冲能力较强;另一方面,优势木可以利用深层土壤稳定的水源,吸水效率高[25],其快速的水分运输可以减轻蒸腾损失引起的树干水分失衡,从而导致较短的水分亏缺和较早的复水。5—6月份的TWDi和TWDd的累积变化量低于其他月份。5—6月降水量相对较大,月均土壤体积含水量(0.248、0.231 m3/m3)高于其他月份(0.210、0.218、0.229、0.221 m3/m3)。年尺度下的红锥树干径向生长速率研究表明,优势木、中等木径向生长最大速度分别出现在5月末、6月中旬[26],结合5—6月份的TWD变化,表明这两个月红锥生长受水分胁迫的程度较轻。
TWD与VPD呈正相关,这与以往关于希腊杜松Juniperus excelsa、辽东栎Quercus wutaishanica、华北落叶松Larix principis-rupprechtii和欧洲云杉Picea abies径向变化对VPD的响应研究结果一致[27-29],表明高VPD将降低大气水势,导致树干流量和蒸腾失水增加,干旱胁迫加剧[30-31]。考虑时滞效应时,二者的相关性更强。TWD反映了树干的贮水量,与蒸腾作用密切相关。径向收缩的时间和幅度提供了关于蒸腾和土壤水分状况随时间的相对变化的信息[32]。从土壤-植物-大气连续体整体的角度来看,植物对环境水分的反应存在时滞[10]。时滞效应是由于叶片蒸腾作用受树木内外因子的影响而发生调节的结果。树木的蒸腾作用受VPD的控制[33],树冠不同高度和方向的叶片气孔蒸腾量不同。这些差异将表现为树干TWD的变化,导致TWD和VPD不同步的现象。本研究中,优势木TWD对VPD的敏感性略低,说明优势木受干旱的影响程度较小,这与刘家霖等[34]的研究结果相似,即受竞争胁迫的兴安落叶松中等木、被压木对高温、低空气湿度的天气更敏感。本研究中,在失水期TWD对VPD的响应时滞缩短,在复水期响应时滞延长,这表明在水分补充阶段和水分损失阶段树木对水分变化的响应不同,可能是因为在水分供应不足的情况下,失水期的径向变化与VPD之间的相关性比在复水期由空气和土壤水分增加引起的径向变化与VPD之间的相关性更强。本研究中,优势木、中等木和被压木的TWDd滞后时间比TWDi分别长3、4和5 h,优势木TWD对VPD的时滞效应更短,造成此现象的原因可能是不同分化等级林木的储水能力和水分运输能力存在差异[24-25]。

深入挖掘高时间分辨率的径向变化数据和与之匹配的环境因子数据,可进一步细化相关研究结论,聚焦不同条件下个体特征的响应机制。本研究中,TWD在不同条件下对VPD的响应模式存在差异,在VPDs条件下,TWD对VPD变化的敏感性较低。对VPDL和VPDs条件下TWD和VPD的相关关系研究表明,考虑时滞效应增加了TWD和VPDL之间的相关性,表明随着VPD的增加,树木将受到更多的水分胁迫。VPDs条件下,在TWDd中,考虑时滞效应减弱了TWD与VPD的相关性,这与兴安落叶松研究中在TWDd中考虑时滞效应增强了TWD与VPD的相关性的结果不同[11],究其原因可能与研究区域有关,其研究区域是大兴安岭地区,年均气温为-4.4 ℃,年均相对湿度为65.9%,年降水量约为473.5 mm,远低于本研究区域的年均气温(20.5~21.7 ℃)、年均相对湿度(80%~84%)和年降水量(1 200~1 600 mm)。VPD对TWD产生限制的阈值不是恒定的,不同气候带的VPD阈值存在差异,如Zweifel等[9]报道了温带树木生长的VPD阈值为0.4 kPa,而对潮湿热带森林的长期森林动态分析表明,热带树木生长的VPD阈值约0.9 kPa[35]。此外,Zweifel等[9]认为,当VPD足够低时,即使土壤适度干燥,树干仍可以进行径向生长;其次可能与树种特性有关,Raffelsbauer等[36]研究表明不同的树种具有不同的水回收能力。
以往径向生长与环境因子的关系研究常考虑多个环境因子,这虽然使研究更加系统,但削弱了研究的深度和针对性。本研究高分辨率的径向生长数据为分析树干径向水分亏缺与饱和水汽压差的关系提供了良好机会,但是单一生长季和点位的研究结果也使得本研究存在一定的局限性,今后的工作可以深入分析不同气候条件下(如干旱年和湿润年)二者相关性的差别,或设置多树种的监测,并以本研究的结果为基础继续探索不同树种对VPD的响应特征。此外,未来的研究应纳入其他因素,如功能性状、树干液流等,对于进一步理解该研究至关重要。
4 结 论
在失水期和复水期,TWD对VPD的响应特征不同,且在不同分化等级间存在差异;失水期和复水期的划分有助于研究TWD与VPD的相关性。TWD与VPD呈正相关,且存在时滞效应。在研究TWD与VPD的相关性时,需要考虑时滞的影响。
参考文献:
[1] 胡衍平,刘卫东,庞文胜,等.水分胁迫对山乌桕幼苗生长及生理生化的影响[J].中南林业科技大学学报,2023,43(8): 62-72. HU Y P, LIU W D, PANG W S, et al. Effects of water stress on the growth, physiological and biochemical characteristics of Sapium discolor seedlings[J]. Journal of Central South University of Forestry & Technology,2023,43(8):62-72.
[2] GROSSIORD C, BUCKLEY T N, CERNUSAK L A, et al. Plant responses to rising vapor pressure deficit[J]. New Phytologist, 2020,226(6):1550-1566.
[3] FANG Z X, ZHANG W M, BRANDT M, et al. Globally increasing atmospheric aridity over the 21st century[J]. Earth’s Future,2022,10(10):e2022EF003019.
[4] DAI A G. Increasing drought under global warming in observations and models[J]. Nature Climate Change,2013,3(1): 52-58.
[5] 张拥兵,王林,薛浩,等.不同干旱程度对枣树苗木水碳平衡和生物量分配的影响[J].经济林研究,2022,40(3):208-215. ZHANG Y B, WANG L, XUE H, et al. Effects of different degrees of drought on water-carbon balance and biomass allocation of jujube seedlings[J]. Non-wood Forest Research,2022,40(3): 208-215.
[6] SKELTON R. Stem diameter fluctuations provide a new window into plant water status and function[J]. Plant Physiology, 2020,183(4):1414-1415.
[7] ZWEIFEL R. Radial stem variations-a source of tree physiological information not fully exploited yet[J]. Plant, Cell & Environment,2016,39(2):231-232.
[8] SALOMóN R L, PETERS R L, ZWEIFEL R, et al. The 2018 European heatwave led to stem dehydration but not to consistent growth reductions in forests[J]. Nature Communications, 2022,13(1):28-38.
[9] ZWEIFEL R, STERCK F, BRAUN S, et al. Why trees grow at night[J]. New Phytologist,2021,231(6):2174-2185.
[10] COCOZZA C, TOGNETTI R, GIOVANNELLI A. Highresolution analytical approach to describe the sensitivity of treeenvironment dependences through stem radial variation[J]. Forests, 2018,9(3):134-151.
[11] TIAN Y, LIU X, ZHANG X, et al. Time lag of stem water deficit in response to increased vapor pressure deficit[J]. Forest Science, 2023,69(3):311-320.
[12] DELUIS M, NOVAK K, CUFAR K, et al. Size mediated climate-growth relationships in Pinus halepensis and Pinus pinea[J]. Trees,2009,23(5):1065-1073.
[13] 赵平.树木储存水对水力限制的补偿研究进展[J].应用生态学报,2010,21(6):1565-1572. ZHAO P. Compensation of tree water storage for hydraulic limitation: Research progress[J]. Chinese Journal of Applied Ecology,2010,21(6):1565-1572.
[14] 邹扬东,王春胜,尹海锋,等.混交比例对西南桦和红锥早期生长与竞争的影响[J].中南林业科技大学学报,2023, 43(11):93-99. ZOU Y D, WANG C S, YIN H F, et al. Effects of mixing ratio on early growth and competition of Betula alnoides and Castanopsis hystrix[J]. Journal of Central South University of Forestry & Technology,2023,43(11):93-99.
[15] 刘小金,徐大平.广东省珍贵树种资源分布特点、产业现状与发展建议[J].广东农业科学,2021,48(7):57-65. LIU X J, XU D P. Characteristics of resource distribution, industry status and development proposal of precious tree species in Guangdong[J]. Guangdong Agricultural Sciences,2021,48(7): 57-65.
[16] 吴国欣,何彦然,张伟,等.广西国家储备林建设现状及高质量发展策略[J].广西林业科学,2022,51(3):445-451. WU G X, HE Y R, ZHANG W, et al. Current situation and high-quality development strategies of national reserve forest construction in Guangxi[J]. Guangxi Forestry Science,2022,51(3): 445-451.
[17] 杨晓慧,杨会肖,徐放,等.不同育苗容器对红锥苗期生长及根系发育的影响[J].中南林业科技大学学报,2021,41(11): 16-26. YANG X H, YANG H X, XU F, et al. Effect of different seedling containers on the growth and root system development of Castanopsis hystrix[J]. Journal of Central South University of Forestry & Technology,2021,41(11):16-26
[18] 玉宝,乌吉斯古楞,王百田,等.兴安落叶松天然林树冠生长特性分析[J].林业科学,2010,46(5):41-48. YU B, WUJISIGULENG, WANG B T, et al. Analysis of growth characteristics in natural Larix gmelinii forests[J]. Scientia Silvae Sinicae,2010,46(5):41-48.
[19] MURRAY F W. On the computation of saturation vapor pressure[J]. Journal of Applied Meteorology,1967,6(1):203-204.
[20] ZWEIFEL R, HAENI M, BUCHMANN N, et al. “Are trees able to grow in periods of stem shrinkage?”[J]. New Phytologist, 2016,211(3):839-849.
[21] GHEYRET G, ZHANG H T, GUO Y P, et al. Radial growth response of trees to seasonal soil humidity in a subtropical forest[J]. Basic and Applied Ecology,2021,55(4):74-86.
[22] CHAN T, H?LTT? T, BERNINGER F, et al. Separating water-potential induced swelling and shrinking from measured radial stem variations reveals a cambial growth and osmotic concentration signal[J]. Plant, Cell & Environment,2015,39(2): 233-244.
[23] MENCUCCINI M, SALMON Y, MITCHELL P, et al. An empirical method that separates irreversible stem radial growth from bark water content changes in trees: theory and case studies[J]. Plant, Cell & Environment,2017,40(2):290-303.
[24] KUMAGAI T, AOKI S, OTSUKI K, et al. Impact of stem water storage on diurnal estimates of whole-tree transpiration and canopy conductance from sap flow measurements in Japanese cedar and Japanese cypress trees[J]. Hydrological Processes, 2009,23(16):2335-2344.
[25] 郭锦荣,白天军,邓文平,等.不同胸径日本柳杉树干液流及其蒸腾耗水差异[J].西南林业大学学报(自然科学),2019, 39(2):70-77. GUO J R, BAI T J, DENG W P, et al. Differences in sap flow and Transpiring water consumption of Cryptomeria japonica with different DBH[J]. Journal of Southwest Forestry University(Natural Sciences),2019,39(2):70-77.
[26] 刘士玲,杨保国,姚建峰,等.广西红椎树干径向生长研究[J].华南农业大学学报,2020,41(5):82-90. LIU S L, YANG B G, YAO J F, et al. Study on stem radial growth of Castanopsis hystrix in Guangxi[J]. Journal of South Agricultural University,2020,41(5):82-90.
[27] ZHANG Q, LYU L X, WANG Y. Patterns of daily stem growth in different tree species in a warm-temperate forest in northern China[J]. Dendrochronologia,2022,72:125934.
[28] OBERHUBER W, HAMMERLE A, KOFLER W. Tree water status and growth of saplings and mature Norway spruce(Picea abies) at a dry distribution limit[J]. Frontiers in Plant Science,2015,6:703-714.
[29] GüNEY A, ZWEIFEL R, TüRKAN S, et al. Drought responses and their effects on radial stem growth of two co-occurring conifer species in the Mediterranean mountain range[J]. Annals of Forest Science,2020,77(4):105.
[30] SáNCHEZ-COSTA E, POYATOS R, SABATé S. Contrasting growth and water use strategies in four co-occurring Mediterranean tree species revealed by concurrent measurements of sap flow and stem diameter variations[J]. Agricultural and Forest Meteorology,2015,207:24-37.
[31] LI W, SI J, YU T, et al. Response of Populus euphratica Oliv. sap flow to environmental variables for a desert riparian forest in the Heihe River Basin, Northwest China[J]. Journal of Arid Land,2016,8(4):591-603.
[32] KING G, FONTI P, NIEVERGELT D, et al. Climatic drivers of hourly to yearly tree radius variations along a 6 ℃ natural warming gradient[J]. Agricultural and Forest Meteorology,2013,168:36-46.
[33] EHRENBERGER W, RüGER S, FITZKE R, et al. Concomitant dendrometer and leaf patch pressure probe measurements reveal the effect of microclimate and soil moisture on diurnal stem water and leaf turgor variations in Young Oak trees[J]. Functional Plant Biology,2012,39(4):297-305.
[34] 刘家霖,满秀玲,胡悦.兴安落叶松天然林不同分化等级林木树干液流对综合环境因子的响应[J].林业科学研究,2016, 29(5):726-734. LIU J L, MAN X L, HU Y. Response of Tree sap flow of Larix gmelinii with various differentiation classes to multiple environmental factors[J]. Forest Research,2016,29(5):726-734.
[35] BAUMAN D, FORTUNEL C, DELHAYE G, et al. Tropical tree mortality has increased with rising atmospheric water stress[J]. Nature,2022,608(7923):528-533.
[36] RAFFELSBAUER V, SPANNL S, PE?A K, et al. Tree circumference changes and species-specific growth recovery after extreme dry events in a montane rainforest in southern Ecuador[J]. Frontiers in Plant Science,2019,10:342-351.
[本文编校:谢荣秀]
