红光、蓝光处理下马铃薯转录组特异表达基因密码子的偏好性分析
2024-09-26许建民程柳洋仇学文













摘要:以红光、蓝光处理下的马铃薯转录组数据为基础,以白光为对照,各筛选出1 206条、691条特异表达基因。通过CodonW、CUSP对筛选出的基因编码序列(CDS)进行密码子使用偏好性分析。结果表明,在红光、蓝光处理下,马铃薯对基因密码子的使用偏好性都较弱,都偏好使用A或T(U)结尾的密码子。在2种光质处理下,特异表达基因序列密码子的GC总量都与同义密码子在第3位的胞嘧啶和鸟嘌呤含量(GC3s)有极显著相关性。在2种光质处理下,高频密码子[相对同义密码子使用度(RSCU)>1.00]均为30个,且高频密码子类型完全一致。从ENc-plot、PR2-plot和中性绘图结果可以看出,2种光质处理下的特异表达基因主要受到自然选择的影响。本研究还分别从蓝光、红光处理的特异表达基因中筛选26个、28个最优密码子。从马铃薯在不同光质处理下特异表达基因与其他生物密码子使用频率的比值及聚类分析结果可以看出,番茄、烟草均可作为红光、蓝光处理下马铃薯特异表达基因的异源表达受体。
关键词:红蓝光;马铃薯;转录组;特异表达基因;密码子;偏好性
中图分类号:S532.01文献标识码:A文章编号:1000-4440(2024)07-1199-13Analysis of codon usage bias in potato transcriptionally specific expressed genes under red and blue light treatmentsXU Jianmin1,CHENG Liuyang2,QIU Xuewen2
(1.Jiangsu Vocational College of Agriculture and Forestry, Jurong 212400, China;2.Agricultural Technology Extension Center of Dongtai City, Dongtai 224200, China;3.Jiangxi Agricultural University, Nanchang 330000, China)Abstract:In this study, 1 206 and 691 specifically expressed genes were screened based on the transcriptome data of potato under red light treatment and blue light treatment, with white light as control, respectively. The coding sequences (CDS) of the selected genes were analyzed by CodonW and CUSP for codon usage bias. The results showed that the codon usage bias of potato genes under both red and blue light treatments was weak, with both preferring codons ending in A or T (U). The total number of sequence codons GC under two light treatments was highly significantly correlated with GC3s. Codons with relative synonymous codon usage (RSCU) greater than 1.00 were high-frequency codons. The number of high-frequency codons under two light treatments was 30, and the types of high-frequency codons were identical. Based on the results of ENc-plot, PR2-plot and neutral plots, it was found that the specifically expressed genes under two light treatments were mainly affected by natural selection. In this study, the 26 and 28 best codons were also screened from genes specifically expressed under blue and red light treatments, respectively. The ratio of codon usage frequency of specifically expressed genes under different light treatments in potato to that in other organisms and the clustering analysis showed that both tomato and tobacco could be used as receptors for heterologous expression of potato specifically expressed genes under red and blue light treatments.
Key words:red and blue light;potato;transcriptome;specifically expressed genes;codon;preference
在自然界中,生物体内的基因表达主要通过转录和翻译2个步骤实现,其中转录是将DNA信息转变为RNA信息,翻译则是将RNA信息转变为蛋白质[1],上述过程离不开密码子的参与。在生物体内有20种氨基酸参与蛋白质的组成,其中部分氨基酸只对应1个密码子,但是大多数氨基酸对应多个密码子,这种1个氨基酸对应的多个密码子称为同义密码子[2]。现有研究发现,生物体在生存和繁衍的过程中,如果密码子的使用是完全随机的,则表明该生物的基因未受到任何偏倚或选择压力[3]。冉然等[4]研究认为,对某一生物或某个基因组而言,若某个密码子频繁出现,则可称该密码子为优化密码子或最优密码子;如果某个密码子极少出现或几乎不出现,则可称其为非优化密码子或稀有密码子。这种密码子出现频率存在明显使用偏好性的现象称为密码子偏好性[5]。
马铃薯(Solanum tuberosum)属于茄科、茄属多年生双子叶植物。光质是马铃薯生长环节中必不可缺的环境因素。目前有很多研究发现,不同光质对马铃薯生长发育有极显著的影响,例如林佳婷等[6]研究发现,按不同比例搭配的红蓝光质可以显著影响马铃薯块茎的形成与淀粉的积累;杨梦平[7]研究发现,纯蓝光和红蓝光的1∶1组合可以有效促进马铃薯组培苗光合色素的合成;韩逸飞等[8]研究发现,在红蓝光照射下的马铃薯对不同矿质元素的吸收与积累也存在显著差异。目前关于大多数红蓝光处理对马铃薯生长发育的影响的研究集中在生理与基因功能验证层面[9-10]。本研究拟以马铃薯品种转心乌在不同光质处理下的叶片转录组数据为基础,筛选出在红蓝光处理下的特异表达基因,并对原始序列进行密码子偏好性分析,将它们与其他物种进行比较,以期为后续马铃薯的基因功能验证及异源表达提供一定理论依据。
1材料与方法
1.1试验材料与处理
cbe774e2c4ef6a7fa00217890bde7d27试验所用马铃薯品种为转心乌,用MS培养基进行体外培养。试验在江苏农林职业技术学院进行。从离体马铃薯植株中无菌采集茎段(长10~15 mm,单叶)后垂直置于MS培养基中,在MS培养基中加入30 g/L蔗糖和7.5 g/L琼脂。在白光荧光灯下预培养3 d后,将各处理组置于蓝灯、红灯下培养30 d。用光谱辐射计(SPIC-200, Everfine Co.,杭州)检测发光二极管(LED)灯的不同光谱特性(蓝光峰值为440 nm,红光峰值为630 nm)。每种光的光合光子通量密度保持在(50±5) μmol/(m2·s)左右,光处理在温度(25±2) ℃、相对湿度75%、光-暗周期12 h-12 h的孵育室中进行。每个处理用10瓶苗,每瓶装10株苗,每个试验设3个重复。采集2个光处理组、1个对照组的所有样品后,立即在液氮中冷冻,保存在-80 ℃。
1.2序列处理
对采集后的样本进行转录组检测。参照刘财国等[11]、Mortazavi等[12]的方法[通过每千个碱基的转录每百万映射读取的片段数(FPKM)计算基因相对表达量],将结果中FPKM≥1的基因定义为高表达基因,而FPKM<1的基因定为低表达或不表达基因。提取出高表达基因的编码序列(CDS)并进行筛选,剔除序列长度小于300 bp的基因、重复基因、不以ATG作为起始密码子的基因、不以TAA/TAG/TGA作为终止密码子的基因、在序列中间存在终止密码子的基因等。
1.3密码子使用偏好性参数分析
整理筛选好的序列后,分别用CodonW、CUSP进行分析,获得在不同光质处理下表达基因序列的相对同义密码子使用度(RSCU)、同义密码子在第3位的胸腺嘧啶含量(T3s,%)、同义密码子在第3位的胞嘧啶含量(C3s,%)、同义密码子在第3位的腺嘌呤含量(A3s,%)、同义密码子在第3位的鸟嘌呤含量(G3s,%)、同义密码子在第3位的胞嘧啶和鸟嘌呤含量(GC3s,%)、密码子在不同位置上的胞嘧啶与鸟嘌呤含量(GC,%)、同义氨基酸数(Lsym)、氨基酸序列的长度(Laa)、平均亲水性值(Gravy值)、芳香族氨基酸频率(Aromo值)、密码子适应指数(CAI)、密码子偏好指数(CBI)、最优密码子使用频率(Fop)、有效密码子数(ENc)、同义密码子的相对使用频率(RFSC)等参数。用SPSS对上述参数进行相关性分析,用R语言绘制相关性分析结果图。
1.4密码子的ENc-plot分析、PR2-plot分析及中性绘图分析以ENc为纵坐标、密码子在第3位的胞嘧啶和鸟嘌呤含量(GC3)为横坐标绘制散点图,得到ENc-plot图。理论ENc采用如下公式计算:ENc=2+GC3+29/[GC32+ (1-GC3)2],以理论ENc为纵坐标、GC3为横坐标绘制标准曲线。对各密码子第3位的A、T、C、G进行定量分析,再以G3/(G3+C3)为横坐标、A3/(A3+T3)为纵坐标(G3、C3、A3、T3分别为密码子第3位的鸟嘌呤、胞嘧啶、腺嘌呤、胸腺嘧啶的含量)绘制散点图,得到PR2-plot图。再绘制X=0.5、Y=0.5 2条参考线,用于分析各密码子碱基的使用差异。以GC12为纵坐标、GC3为横坐标绘制散点图,得到中性图。再绘制Y=x函数曲线,作为参考线,用于后续分析。用Excel对相关点进行拟合,得到相关系数、回归系数与拟合函数。作图均用Origin。
1.5最优密码子的确定
最优密码子的确定采用Wright[13]的方法。首先以ENc为依据,由高到低进行排序,分别取两端10%的基因建立高低表达基因库。再以RSCU>1.00为条件,在高表达基因库中筛选高频密码子,并用高表达基因库密码子的RSCU减去低表达基因库密码子的RSCU,计算出△RSCU,再以△RSCU≥0.08为条件,筛选出最优密码子。
1.6其他物种密码子使用频率的对比
将用方法1.2处理后的序列上传至cusp网站(http://emboss.toulouse.inra.fr/cgi-bin/emboss/cusp),计算出不同光质处理下马铃薯特异表达基因的相对使用频率(RFSC),再在Kazusa DNA研究所网站下载模式物种拟南芥(Arabidopsis thaliana)、大肠杆菌(Escherichia coli )、水稻(Oryza sativa)、番茄(Solanum lycopersicum)、烟草(Nicotiana tabacum)及酵母(Saccharomyces cerevisiae)的密码子相对使用频率,并对这些物种密码子的相对使用频率与马铃薯进行比较及聚类分析,从而进一步分析不同光质处理下马铃薯基因密码子的使用偏好性模式。
2结果与分析
2.1序列筛选及密码子使用偏好性参数的分析
对测序公司提供的转录组数据进行筛选发现,在蓝光处理下,在马铃薯中特异表达的基因有1 206条,而在红光处理下,在马铃薯中特异表达的基因有691条。对这些基因序列进行密码子使用偏好性参数分析,发现在红光、蓝光处理下C3s、CAI、CBI、Fop、GC、Aromo值这6项参数的平均值相等,且蓝光处理下T3s、A3s的平均值大于红光处理下T3s、A3s的平均值(表1),表明在蓝光处理下,马铃薯更偏向于使用以T、A结尾的同义密码子。由表1可以看出,在蓝光处理下,T3s、G3s、Fop、ENc、GC3s、GC、Gravy值等参数的最小值小于红光处理下的最小值,C3s、Laa、Aromo值等参数的最小值与红光处理下的最小值相等,其余参数的最小值均大于红光处理下的最小值。此外,在蓝光处理下,ENc、Aromo值的最大值与红光处理下的最大值相等,其余参数的最大值均大于红光处理下的最大值。通过对变异系数的分析可以看出数据的离散程度。由表1还可以看出,在红光处理下,除CAI、CBI、ENc、GC3s、Gravy值外,其余参数的离散程度均略大于蓝光处理下的离散程度。在密码子使用方面,2种光质处理下的马铃薯特异表达基因的GC碱基使用总量较为接近,密码子第3位GC碱基的使用总量也均小于35%,表明2种光质都偏向使用以A、T结尾的密码子。
2.2不同光质处理下马铃薯转录组密码子使用偏好性参数相关性分析对红光、蓝光处理下马铃薯转录组中特异表达基因的密码子使用偏好性相关参数进行分析。如图1所示,在2种光质处理下,T3s与GC1间均无显著相关性,与CAI呈极显著正相关,与其他参数间均表现为极显著负相关;在蓝光处理下,C3s与G3s有显著正相关性,与GC1无显著相关性;在红光处理下,C3s与G3s无显著相关性,与GC1呈显著正相关;在2种光质处理下,A3s与CAI、G3s、CBI、Fop、ENc、GC1、GC2、GC3等参数之间的相关性趋势相同,均呈极显著负相关;在蓝光处理下,CAI与GC2呈极显著正相关,而在红光处理下的CAI与GC2则无显著相关性。
2.3不同光质处理下马铃薯转录组密码子相对使用度分析对不同光质处理下马铃薯转录组特异表达基因的同义密码子相对使用度进行分析。如表2所示,在蓝光处理下,RSCU值大于1.00的密码子有30个,而在红光处理下,RSCU值大于1.00的密码子同样有30个,且在蓝光、红光处理下RSCU值大于1.00的密码子种类完全相同。在上述30个密码子中,没有以C结尾的密码子,以A结尾的密码子有12个,占比为40%,以G结尾的密码子有3个,占比为10%,以U结尾的密码子有15个,占比为50%。上述研究结果表明,在蓝光、红光2种光质处理下,在马铃薯转录组中特异表达的基因均偏好使用以A或U(T)结尾的密码子。
2.4中性绘图分析
对筛选出来的序列进行分析后,进行中性绘图。如图2、图3所示,在蓝光处理下,GC12的取值范围为0.236~0.723,GC3的取值范围为0.086~0.789;在红光处理下,GC12的取值范围为0.252~0.600,GC3的取值范围为0.131~0.633。从图2还可以看出,蓝光处理下的马铃薯特异表达基因密码子大多数落在对角线的上方,少部分基因密码子落在对角线下方,表明在不同位置上,G、C的使用主要受到自然选择的影响,GC12与GC3之间的相关系数为0.272 93,相关性不显著,数据拟合后的回归系数为0.175 46,也说明在蓝光处理下的密码子GC12与GC3主要受到自然选择的影响。从图3还可以看出,红光处理下的马铃薯特异表达基因密码子大多数落在对角线的上方,同样各个位置的碱基使用受到自然选择的影响,但在对角线下方的基因密码子数量占比有所提升。进行线性拟合后发现,GC12与GC3之间的相关系数为0.368 12,相关性不显著,回归系数为0.235 55,同样表明红光处理下的密码子GC12与GC3主要受到自然选择的影响。
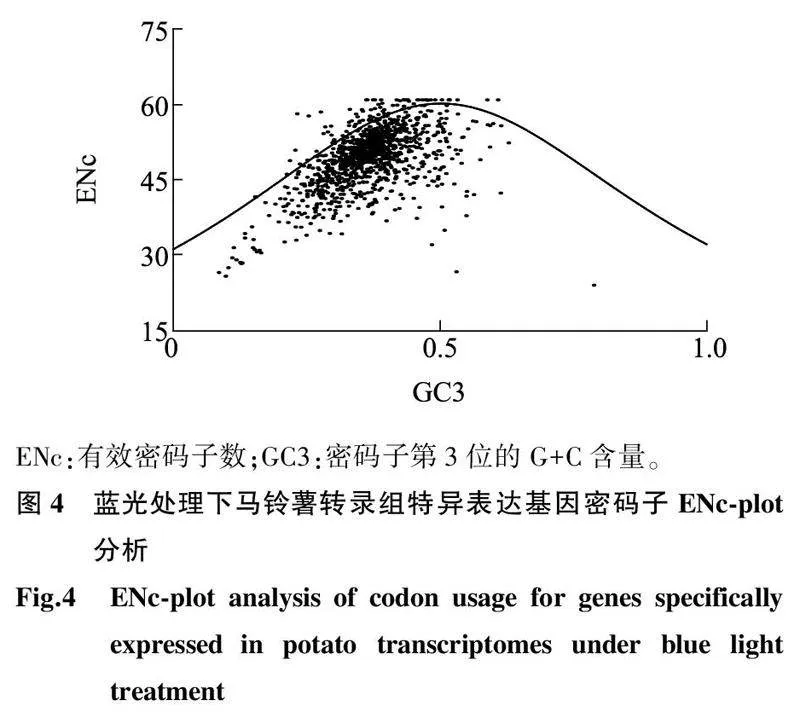
2.5ENc-plot分析
对筛选出来的基因进行ENc-plot分析,根据各基因与ENc标准曲线的相对位置,可以判断影响第3位碱基的因素。从图4可以看出,大多数基因密码子落在标准曲线的下方,表明这些基因密码子的使用选择主要受到自然选择的影响,而少部分基因密码子落在标准曲线的上方,表明这部分基因主要受到基因突变的影响。在图5中,同样也是大多数基因密码子落在标准曲线下方,部分基因密码子在标准曲线的上方,表明红光处理下的马铃薯特异表达基因密码子碱基选择同样主要受到自然选择的影响。
2.6PR2-plot分析
从图6、图7可以看出,在蓝光、红光处理下,马铃薯特异表达基因密码子主要集中在PR2-plot图的右下角区域,表明在2种光处理下,密码子第3位碱基对碱基A的使用频率低于对碱基T的使用频率,对碱基C的使用频率低于对碱基G的使用频率。此外,在靠近中心位置也有部分基因分布,表明这部分基因的密码子组成不仅受到自然选择的影响,同时也受到基因突变的影响。
2.7最优密码子分析
最优密码子的定义是同时具备高频率、高表达特征的密码子[14]。从表3可以看出,蓝光处理下的最优密码子有26个,红光处理下的最优密码子有28个。红光、蓝光处理下共有的最优密码子有25个,其中AGG为蓝光处理下特有的最优密码子,UUG、CUU、AUU为红光处理下特有的最优密码子。从密码子组成来看,蓝光处理下的最优密码子中,有13个以A结尾,1个以G结尾,12个以U结尾;在红光处理下的最优密码子中,有13个以A结尾,1个以G结尾,14个以U结尾。在2种处理下的最优密码子中,均无以C结尾的密码子,均只有1个以G结尾的密码子,表明在2种光质处理下,马铃薯转录组中特异表达基因的最优密码子都偏好以A或U(T)结尾。
2.8马铃薯与其他物种密码子使用偏好性的比较
同义密码子的相对使用频率也可作为衡量密码子使用偏好性的一个重要指标。当2个物种的密码子相对使用频率比值为0.50~2.00时,说明这2个物种对该密码子的使用特性较为一致;当2个物种的密码子相对使用频率比值≥2.00或≤0.50时,则认为这2个物种的密码子使用特性差异较大[15]。由图8可以看出,在蓝光处理下,马铃薯中特异表达基因的密码子相对使用频率与拟南芥、烟草、番茄相比无较大差异,而与大肠杆菌、酵母、水稻和大豆相比,分别有16个、5个、19个和1个相对使用频率差异较大的密码子。由图9可以看出,在红光处理下,马铃薯中特异表达基因的密码子相对使用频率与拟南芥、烟草、番茄相比也无较大差异,而与大肠杆菌、酵母、水稻和大豆相比,分别有16个、6个、19个和1个相对使用频率差异较大的密码子。上述结果表明,在2种光质处理下的马铃薯特异表达基因在受体选择方面具有一定的同质性,均可优先选择拟南芥、烟草和番茄。
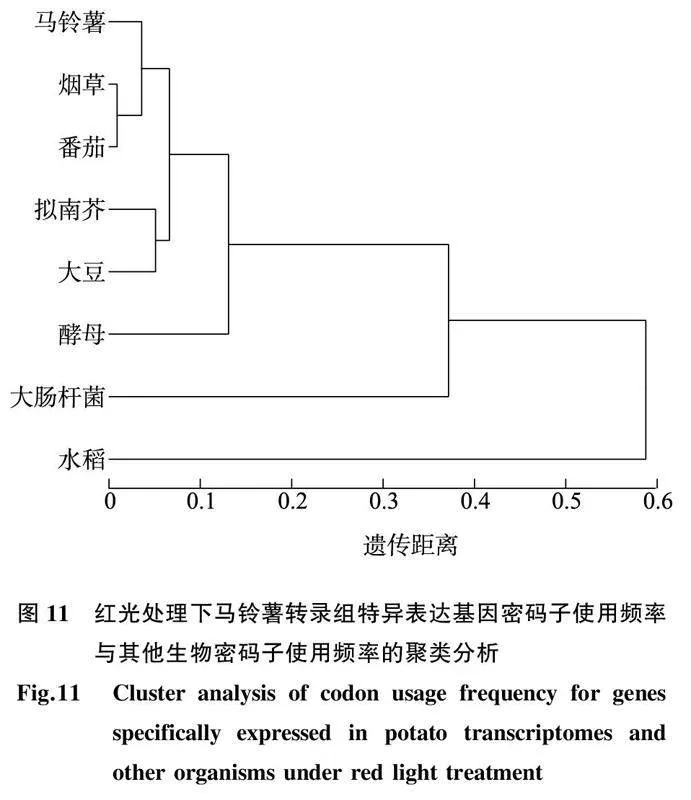
对不同光质处理下在马铃薯中特异表达基因的密码子使用频率与8种其他生物的密码子使用频率进行聚类分析。如图10所示,在蓝光处理下,当对马铃薯中特异表达基因进行异源表达试验受体选择时,可以优先选择烟草、番茄,之后可依次选择拟南芥、大豆和酵母等,这与通过比值计算得出的结果基本吻合。如图11所示,在红光处理下,马铃薯中特异表达基因在进行异源表达试验受体选择时,同样可以优先选择烟草、番茄,其次是拟南芥、大豆。在2种光质处理下,马铃薯中特异表达基因在进行异源表达试验受体选择时,均不推荐选择水稻、大肠杆菌,这与通过比值计算得出的结果基本吻合。图8蓝光处理下马铃薯转录组基因与其他生物密码子使用偏好性(相对使用频率比值)的比较
3讨论
密码子在基因表达过程中起着重要作用[16-20],研究生物体内密码子的使用模式与使用特性对于揭示植物基因的转录翻译效率及相对应的功能表达具有极其重要的意义[21]。现有研究发现,不同物种在密码子的使用过程中存在偏好性现象[22]。生物体在使用密码子时,通常会受到多种因素的影响,主要包括在进化过程中的自然选择及受环境影响导致的基因突变压力,继而导致基因结构、功能表达发生相应改变[13]。
在本研究中,蓝光处理下的马铃薯特异表达基因数量为1 206条,约为红光处理下的1.7倍。冯琛等[23]研究发现,蓝光处理下的草莓转录组特异表达基因数量明显多于红光处理,约为红光处理的2倍,这与本研究结果基本一致;但是,草莓在红光、蓝光处理下的特异表达基因数量分别为56条、114条,与本研究结果差异较大,这或许与物种不同或其他生长环境因子不同有关。对红光、蓝光处理下的马铃薯转录组基因密码子使用偏好性参数进行分析,发现两者都偏好使用以A或T(U)结尾的密码子,这与对白刺[24]、橄榄[25]等的研究结果类似,且在2种光质处理下,密码子的总C+G含量约为30%,均明显小于50%,这些现象也符合大多数双子叶植物对密码子使用偏好性的特征[26]。从密码子使用偏好性参数相关性分析结果可以看出,在蓝光处理下,马铃薯中特异表达drj7wK+4vDckNR9rWc43yKEQqh1zS6kEnvK2iw6vitY=基因密码子第3位碱基G与C之间有显著相关性,而在红光处理下则无显著相关性。本研究还发现,在蓝光处理下,G3s与C3s的变异系数均小于红光处理,表明蓝光处理下的特异表达基因数量多且稳定。除此以外,在2种光质处理下的特异表达基因的第3位碱基的占比与其他偏好性参数呈现极显著正相关,这或许是由于对于密码子碱基的选择而言,通常第3位碱基较为保守,自然选择对其造成的影响较小[27],这与刘洋等[28]对向日葵的研究结果基本一致。
同义密码子相对使用度(RSCU值)是衡量密码子使用偏好性最重要、最广泛的参数[29]。通过分析RSCU值的大小,可以判断相应密码子的实际使用频率与期望使用频率之间的比值,从而分析该密码子的使用偏好性[30]。在本研究中,红光、蓝光处理下的马铃薯特异表达基因的高频密码子种类完全一致,都偏好使用以A或T(U)结尾的密码子,这或许与马铃薯属于双子叶植物有关,这与前人的相关研究结果[31-32]基本一致。对最优密码子进行研究发现,在红光、蓝光处理下,共有25个相同的最优密码子,蓝光处理下的最优密码子略少于红光处理下的最优密码子,前人在红光、蓝光处理下草莓密码子的使用偏好性研究中也发现类似现象[23],认为该现象与2种处理下特异表达基因的数量差距较大有关。从最优密码子的选择性上也可以看出,在红光、蓝光处理下,马铃薯中的特异表达基因均偏好以A或T(U)结尾。
从ENC-plot分析结果可知,在2种光质处理下,马铃薯特异表达基因只有少数分布于标准曲线上方,表明这部分基因在密码子使用中主要受到碱基突变的影响;大部分基因在标准曲线的下方,表明这部分基因在密码子使用中主要受到自然选择的影响。在蓝光处理下受到碱基突变影响的基因数量略多于红光处理下的基因数量,这或许是由于在蓝光处理下的特异表达基因多于红光处理下的特异表达基因,在冯琛等[23]的研究中也发现了相关现象。但是从占比来看,在红光处理下,密码子使用受碱基突变影响的基因占比略大于蓝光处理下的相应占比。从PR2-plot分析结果可以看出,密码子第3位碱基存在明显的偏好性,在2种光质处理下,T的使用频率明显大于A,G的使用频率明显大于C。现有研究结果表明,如果密码子使用偏好性完全由碱基突变决定,则A和T、C和G的使用频率应该相等[33],而在本研究中,密码子的使用呈现了明显的偏好性,表明在2种光质处理下的马铃薯特异表达基因密码子选择主要受到自然选择的影响,部分密码子受碱基突变的影响,与红光处理相比,蓝光处理下的基因更多集中在坐标(0.5,0.5)附近,表明蓝光处理下受碱基突变影响的密码子数量略多于红光处理。从中性绘图结果也可以看出,在2种光质处理下,马铃薯中特异表达基因的密码子使用主要受到自然选择的影响,少部分受到碱基突变的影响,蓝光处理下受到碱基突变影响的基因数量略多于红光处理下的。比较两者的回归系数可以发现,自然选择对蓝光处理下密码子碱基的选择要强于红光处理下的,这也进一步验证了以上猜想。
有部分研究者认为,不同物种的密码子相对使用频率比值可以为目标基因的异源生物转化与表达提供参考[34]。晁岳恩等[35]研究发现,在目的基因进行异源表达时,通常要研究受体生物的密码子使用偏好性,进而对目的基因进行改造与修饰,从而提高目的基因在受体中的表达效率与成功率。在本研究中,蓝光处理下的马铃薯特异表达基因可优先考虑以番茄、烟草作为基因表达受体,在红光处理下则同样可以优先考虑以番茄、烟草作为基因异源表达的首要选择,这或许与番茄、烟草、马铃薯同为茄科作物有关。何柳等[36]在进行金银花基因的异源表达时发现,拟南芥比大肠杆菌、酵母更适合作为基因异源表达的底盘细胞。在本研究中,对拟南芥、酵母和大肠杆菌进行密码子使用频率比较研究发现,在进行受体选择时,拟南芥优于酵母,酵母又优于大肠杆菌,这与前人的研究结果基本一致,这或许是由于拟南芥、马铃薯同属于双子叶植物,酵母、马铃薯同属于真核生物,而大肠杆菌属于原核生物。水稻最不适合作为马铃薯异源表达的受体,这或许与水稻属于单子叶植物而马铃薯属于双子叶植物有关。本研究结果可为马铃薯特异表达基因的异源表达提供一定的理论基础,但具体表达效率还受其他因素影响[37],需深入研究。
参考文献:
[1]谢平. 遗传密码子的起源——从能量转化到信息化[J]. 生物多样性,2017,25(1):94-106.
[2]NOVOA E M, PAVON-ETERNOD M, PAN T, et al. A role for tRNA modifications in genome structure and codon usage[J]. Cell,2012,149(1):202-213.
[3]任桂萍,董璎莹,党云琨. 密码子中的密码:密码子偏好性与基因表达的精细调控[J]. 中国科学:生命科学, 2019,49(7):839-847.
[4]冉然,张晓明,宛涛,等. 密码子偏好性的相关研究及方法分析[J]. 草原与草业,2022,34(3):5-10,43.
[5]HERSHBERG R, PETROV D A. Selection on codon bias[J]. Annual Review of Genetics,2008,42(1):287-299.
[6]林佳婷,陈松,徐志刚. 不同红蓝光比例对马铃薯块茎形成与产量的影响[J]. 南京农业大学学报,2022,46(3):447-453.
[7]杨梦平. 不同光质LED光源对马铃薯组培苗光合色素含量的影响[J]. 黑龙江农业科学,2022(6):27-30.
[8]韩逸飞,顾天行,黄梦圆,等. 红蓝LED光源对马铃薯组培苗形态生长、光合色素含量及矿质元素积累的影响[J]. 中国马铃薯,2021,35(4):289-299.
[9]樊颖伦,孙艳茹,卞春松,等. 不同光质对紫色马铃薯薯块花青素含量的影响[J]. 聊城大学学报(自然科学版),2013,26(4):76-79.
[10]季彦林,王旺田,王蒂,等. 不同光质对马铃薯块茎糖苷生物碱积累的诱导效应[J]. 江苏农业学报,2010,26(1):40-45.
[11]刘财国,于文涛,王鹏杰,等. 基于转录组学的茶树品种福建水仙器官特异性分析[J/OL]. 分子植物育种,2021:1-16[2023-06-01]. http://kns.cnki.net/kcms/detail/46.1068.S.20210925.0055.002.html.
[12]MORTAZAVI A, WILLIAMS B C, MCCUE K, et al. Mapping and quantifying mammalian transcriptomes by RNA-Seq[J]. Nature Methods,2008,5(7):621-628.
[13]WRIGHT F. The ‘effective number of codons’ used in a gene[J]. Gene,1990,87(1):23-29.
[14]PARVATHY S T, UDAYASURIYAN V, BHADANA V. Codon usage bias[J]. Molecular Biology Reports,2022,49(1):539-565.
[15]仪登霞,杨丽梅,方智远,等. 结球甘蓝核基因组密码子使用偏爱性分析[J]. 园艺学报,2013,40(10):1927-1934.
[16]赵振宁,孙浩田,宋雨茹,等. 山楂属植物叶绿体基因组特征与密码子偏好性分析[J]. 江苏农业学报,2023,39(2):504-517.
[17]王飞,辛雅萱,董章宏,等. 无刺龙舌兰叶绿体基因组特征及密码子偏好性分析[J]. 南方农业学报,2022,53(4):1030-1039.
[18]刘潮,韩利红,盛巧,等. 8种植物类甜蛋白家族基因进化及密码子特征分析[J]. 江苏农业科学,2022,50(10):44-51.
[19]王占军,吴子琦,王朝霞,等. 3个茶树品种WOX基因家族的进化及密码子偏好性比较[J]. 南京林业大学学报(自然科学版),2022,46(2):71-80.
[20]刘潮,韩利红,吴丽芳,等. 辣椒基因组密码子使用偏性分析[J]. 江苏农业科学,2022,50(5):16-22.
[21]IRIARTE A, LAMOLLE G, MUSTO H. Codon usage bias: an endless tale[J]. Journal of Molecular Evolution, 2021,89(9/10):589-593.
[22]ARELLA D, DILUCCA M, GIANSANTI A. Codon usage bias and environmental adaptation in microbial organisms[J]. Molecular Genetics and Genomics,2021,296(3):751-762.
[23]冯琛,汤浩茹,江雷雨,等. 红蓝光对草莓转录组特异表达基因密码子使用偏好性的影响[J]. 浙江农业学报, 2017,29(4):566-574.
[24]张得芳,夏涛. 基于转录组的唐古特白刺密码子偏好性分析[J]. 干旱区资源与环境,2021,35(12):163-170.
[25]赖瑞联,冯新,陈瑾,等. 橄榄转录组密码子使用偏好性及其影响因素[J]. 核农学报,2019,33(1):31-38.
[26]MURRAY E E, LOTZER J, EBERLE M. Codon usage in plant genes[J]. Nucleic Acids Research,1989,17(2):477-498.
[27]LI X, WANG X C, GONG P T, et al. Analysis of codon usage patterns in Giardia duodenalis based on transcriptome data from Giardia DB[J]. Genes,2021,12(8):1169.
[28]刘洋,路妍,景岚. 向日葵全基因组NBS抗病基因密码子使用偏好性分析[J]. 江苏农业科学,2021,49(10):43-47.
[29]SHARP P M, LI W H. The codon adaptation index-a measure of directional synonymous codon usage bias, and its potential applications[J]. Nucleic Acids Research,1987,15(3):1281-1295.
[30]吴宪明,吴松锋,任大明,等. 密码子偏性的分析方法及相关研究进展[J]. 遗传,2007,29(4):420-426.
[31]王茹,罗永坚,刘相泉,等. 水杉叶绿体基因组同义密码子使用偏好性[J]. 湖北民族大学学报(自然科学版),2022,40(2):121-128.
[32]肖明昆,严炜,熊贤坤,等. 云南樟叶绿体基因组密码子偏好性分析[J]. 中南林业科技大学学报, 2022,42(6):127-134.
[33]PLOTKIN J B, KUDLA G. Synonymous but not the same: the causes and consequences of codon bias[J]. Nature Reviews Genetics,2011,12(1):32-42.
[34]孙继峰,韩太利,徐立功,等. 萝卜ICE1密码子使用偏性分析[J]. 核农学报,2018,32(3):477-485.
[35]晁岳恩,吴政卿,杨会民,等. 11种植物psbA基因的密码子偏好性及聚类分析[J]. 核农学报,2011,25(5):927-932.
[36]何柳,朱英杰,孙超. 金银花中表达基因密码子偏好性分析[J]. 世界科学技术-中医药现代化, 2013,15(3):360-366.
[37]赖瑞联, 林玉玲, 钟春水, 等. 龙眼生长素受体基因TIR1密码子偏好性分析[J]. 园艺学报,2016,43(4):771-780.(责任编辑:徐艳)
