Protein succinylation, hepatic metabolism, and liver diseases
2024-05-08ShuangLiuRuiLiYaWenSunHaiLinHaiFangLi
Shuang Liu, Rui Li, Ya-Wen Sun, Hai Lin, Hai-Fang Li
Abstract Succinylation is a highly conserved post-translational modification that is processed via enzymatic and non-enzymatic mechanisms. Succinylation exhibits strong effects on protein stability, enzyme activity, and transcriptional regulation. Protein succinylation is extensively present in the liver, and increasing evidence has demonstrated that succinylation is closely related to hepatic metabolism. For instance, histone acetyltransferase 1 promotes liver glycolysis, and the sirtuin 5-induced desuccinylation is involved in the regulation of the hepatic urea cycle and lipid metabolism. Therefore, the effects of succinylation on hepatic glucose, amino acid, and lipid metabolism under the action of various enzymes will be discussed in this work. In addition, how succinylases regulate the progression of different liver diseases will be reviewed, including the desuccinylation activity of sirtuin 7, which is closely associated with fatty liver disease and hepatitis, and the actions of lysine acetyltransferase 2A and histone acetyltransferase 1 that act as succinyltransferases to regulate the succinylation of target genes that influence the development of hepatocellular carcinoma. In view of the diversity and significance of protein succinylation, targeting the succinylation pathway may serve as an attractive direction for the treatment of liver diseases.
Key Words: Protein succinylation; Hepatic metabolism; Fatty liver; Hepatitis; Hepatocellular carcinoma
INTRODUCTION
Introduction to protein succinylation
Post-translational modification is an important mechanism that affects protein function, integrating metabolism with physiological and pathological processes. Succinylation is an important post-translational modification of proteinsviaboth enzymatic and non-enzymatic manners[1].
Process of succinyl modification:Succinylation is the process by which a succinyl donor transfers a negatively charged four-carbon succinyl group to the amine of lysine residues by enzymatic or non-enzymatic means[2,3] (Figure 1). The succinyl group binding to the lysine residue has a relatively larger molecular weight (approximately 100.02 Da), which significantly changes the protein structure. Additionally, the charge carried by the lysine residues changes from +1 to -1, resulting in alterations to the physical and chemical properties as well as the functions of the proteins[1-4].
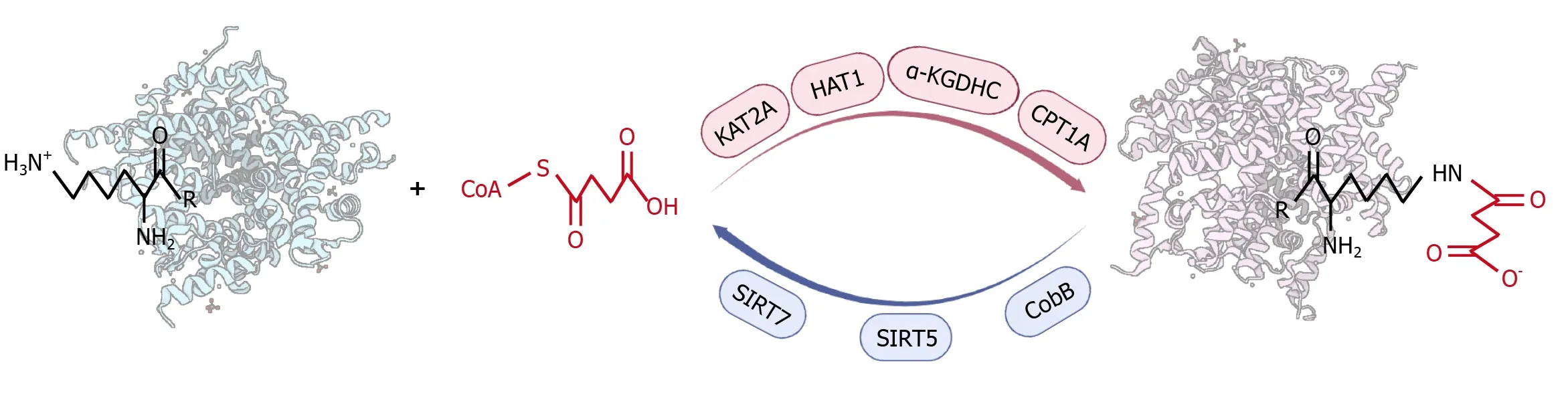
Figure 1 Mechanisms for succinylation. Succinylation is the process of transferring negatively charged four-carbon succinyl groups to amines of lysine residues through enzymatic and non-enzymatic manners using succinyl-CoA as a direct substrate. The succinylation degree can be promoted by succinyltransferases, such as lysine acetyltransferase 2A, histone acetyltransferase 1, α-ketoglutarate dehydrogenase complex, and carnitine palmitoyltransferase 1A. Meanwhile, desuccinylases, including CobB, sirtuin 5, and sirtuin 7 negatively regulate the extent of protein succinylation. KAT2A: Lysine acetyltransferase 2A; HAT1: Histone acetyltransferase 1; α-KGDHC: α-ketoglutarate dehydrogenase complex; SIRT5: Sirtuin 5; SIRT7: Sirtuin 7; CPT1A: Carnitine palmitoyltransferase 1A.
Succinyl modification is widespread in both the cytoplasm and nucleus[5]. In the cytoplasm, succinylation is highly concentrated in mitochondria and may be involved in regulating the tricarboxylic acid cycle, amino acid metabolism, and fatty acid metabolism[6-9]. In the nucleus, lysine succinylation is present in more than one-third of nucleosomes, and the succinylation sites are mainly enriched in the gene promoter region, suggesting that succinylation may be involved in the transcriptional regulation of genes[6,8,9]. Succinylated lysine residues have greater structural changes and charge differences than other typical covalent lysine modification groups such as acetyl and dimethyl[1,5]. Therefore, the influence and mechanism of succinylation on the target proteins and its potential application for the treatment of metabolic diseases have received increasing research attention.
Mechanisms for succinylation:On one hand, succinylation could be processedvianon-enzymatic manners, which relies on succinyl-CoA or succinate from mitochondrial and peroxisome sources[4,5,10-14]. Succinylation would occur if provided with sufficient succinyl-CoA[10]. It has been established that mixing succinyl-CoA with albumin or isocitrate dehydrogenase increases succinylation and mitochondrial pH in a pH-dependent and dose-dependent manner[4,5]. Sreedharet al[11] showed that nicotinamide adenine dinucleotide phosphate-specific isocitrate dehydrogenase mutation results in a 280% increase in cellular succinyl-CoA levels and mitochondrial hyper-succinylation. Succinate dehydrogenase inactivation induces excessive succinylationviaincreasing the accumulation of succinyl-CoA[12]. Notably, tissues with high levels of succinyl-CoA also show a strong extent of succinyl modification, such as in the heart and liver[13]. Succinate entering the cells could be converted to succinyl-CoA to enhance lysine succinylation[1]. A study has shown that dietary succinate increases the succinylation of intestinal and hepatic proteins with a molecular weight of 25-35 kD in zebrafish[14].
On the other hand, the extent of succinylation could be positively regulated by several enzymes that play succinylwriter roles (Figure 1)[15-19], even though no specific succinyltransferases have been identified to date. For example, lysine acetyltransferase 2A (KAT2A) was found to be a succinyltransferase[15,16], which can reportedly upregulate H3K79 succinylation and β-catenin stabilization, thereby promoting glycolysis[20]. Zhouet al[21] confirmed that KAT2A promotes the succinylation of K46 and K280 of C-terminal binding protein 1 and mediates the transcription-suppressing activity. In addition, histone acetyltransferase 1 (HAT1) was identified as a succinyltransferase of both histone and nonhistone proteins[17,22]. HAT1 mediates the succinylation of histones, and quantitative proteomic analysis revealed five succinylation sites on 45 histones[17]. Research has shown that HAT1 is necessary for the regulation of epigenetic and gene expression by H3K122 succinylation[17].
Wanget al[22] and Yanget al[17] demonstrated that phosphoglycerate mutase 1 (PGAM1), a critical enzyme in glycolysis, is succinylated by HAT1 at K99. The later report also mentioned that aspirin downregulates HAT1 by targeting NF-kappaB to induce PGAM1 K99 desuccinylation, which suppresses the glycolytic process[22]. Furthermore, the α-ketoglutarate dehydrogenase complex (α-KGDHC) regulates succinylation either by regulating succinyl-CoA levels or by directly catalyzing succinylation[4,18]. Inhibition of α-KGDHC reduces succinylation levels of both cytoplasmic matrix and mitochondrial proteins[4]. The E2k subunit of α-KGDHC was demonstrated to be essential for its transsuccinylase activity. The absence of the E2k subunit reduces succinylation, while the presence of alpha-ketoglutaric acid increases succinylation[4].
Another lysine succinyltransferase in mammalian cells is carnitine palmitoyltransferase 1A (CPT1A)[19]. Kurmiet al[19] demonstrated that CPT1A can play the role of a succinyltransferase bothin vivoandin vitroto regulate substrate proteins and related metabolic processes. Wanget al[23] discovered that CPT1A-mediated succinylation of S100A10 (a protein that is overexpressed in gastric cancer) increases human gastric cancer invasion. Moreover, CPT1A promotes the succinylation of mitochondrial fission factor at K302 and enhances the development of ovarian cancer[24].
In addition, significant progress has been made in the exploration of desuccinylases that negatively regulate succinylation (Figure 1). CobB was the first desuccinylase discovered in prokaryotes with both deacetylation and desuccinylation activities[25]. A high-performance liquid chromatography assay showed that CobB could deacetylate and desuccinylate a histone H3K9 peptide with similar efficiency, whereas the desuccinylation activity of CobB might be induced when cells are treated with succinate[25].
Sirtuin 5 (SIRT5) and sirtuin 7 (SIRT7) are currently known as important desuccinylases in eukaryotes[26-32]. SIRT5 acts in all cell compartments. The activity of SIRT5 is dependent on NAD+, which is influenced by the availability of NAD+(substrate) and the amount of nicotinamide (product)[26]. In SIRT5 knockouts, more than 80% of proteins are succinylated in the tricarboxylic acid cycle to enhance cell respiration, and 60% of proteins in fatty acid metabolism are succinylated[27]. At least 2565 succinylation sites on 779 proteins in mammalian fibroblasts and liver tissues were found to be regulated by SIRT5[27]. Novel targets for SIRT5 in regulating the mitochondrial lysine succinylome such as uncoupling protein 1 in mouse brown adipose tissue were recently identified[28,29].
SIRT7 is a member of the sirtuin family proteins that are described as NAD (+)-dependent class III histone deacetylases[30,31]. Research indicated that SIRT7 catalyzed the desuccinylation of H3K122, which promoted chromatin condensation and DNA double-strand break repair[30]. Yuet al[31] showed that SIRT7 restricted chronic hepatitis B virus (HBV) transcription and replication through catalyzing desuccinylation of H3K122 that is associated with covalently closed circular (ccc) DNA minichromosome. SIRT7 mediates the desuccinylation of arginine methyltransferase 5 (PRMT5) K387, which is involved in lipid reprogramming, tumor growth, and metastasis[32].
Collectively, succinylation is the process of transferring negatively charged four-carbon succinyl groups to amines of lysine residues through enzymatic and non-enzymatic manners using succinyl-CoA as a direct substrate. The succinylation degree could be promoted by succinyltransferases, such as KAT2A, HAT1, α-KGDHC, and CPT1A. Meanwhile, desuccinylases, including CobB, SIRT5, and SIRT7 negatively regulate the extent of protein succinylation. To date, the characterization of succinyltransferases and desuccinylases, their target specificity, the function of succinylation, and their clinical application still need to be further investigated, given their significance for proteomic analysis.
Effects of succinylation on hepatic metabolism
The liver is a crucial metabolic organ through which major metabolic processes including glucose, amino acid, and lipid metabolisms occur[33]. The overall abundance of lysine succinylation in the liver is higher relative to other tissues, with proteins and enzymes in several metabolic pathways being succinylated[34].
Influence of protein succinylation on glucose and amino acid metabolism:Glucose homeostasis is largely regulated by hepatic glycogen synthesis, gluconeogenesis, and glycolysis[35,36]. Enhancement of glycolysis contributes to the growth of tumor cells. Yanget al[17] performed a Kyoto Encyclopedia of Genes and Genomes pathway enrichment analysis on HAT1-targeted non-histone proteins and found that HAT1 mediates the succinylation of glycolytic-related proteins, including seven key enzymes including GPI, TPI, GAPDH, PGK, PGAM, enolase, and PKM. The authors further demonstrated that the HAT1-induced K99 succinylation of PGAM1 increased its activity, which further promoted tumorigenesis[17]. Wanget al[22] showed that aspirin reduced HAT1 expression, which decreased the K99 succinylation level of PGAM1, thereby restricting PGAM1 activity and inhibiting glycolysis in liver cancer (Figure 2).

Figure 2 Effect of succinylation on hepatic metabolic pathways. The influence of succinylation on hepatic glucose metabolism occurs in the following ways: (1) Under the stimulation of succinyl-CoA. Histone acetyltransferase 1 causes the K99 site of phosphoglycerate mutase 1 to be succinylated and promotes its enzyme activity, thus promoting glycolysis; (2) the influence of succinylation on hepatic amino acid metabolism. Sirtuin 5 promotes urea production by regulating the desuccinylation of arginine succinate synthetase 1 and carbamoyl phosphate synthase 1; and (3) the influence of succinylation on hepatic lipid metabolism. Sirtuin 5 induces desuccinylation of mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthetase 2 and promotes ketone body formation. HAT1: Histone acetyltransferase 1; SIRT5: Sirtuin 5; TCA: Tricarboxylic acid; HMGCS2: 3-hydroxy-3-methylglutaryl-CoA synthetase 2; PGAM1: Phosphoglycerate mutase 1; ASS1: Arginine succinate synthetase 1.
The liver is also a major tissue for the conversion of ammonia[37], which is a toxic metabolite derived from amino acid metabolism under physiological conditions[38]. For the conversion of ammonia to non-toxic ureaviathe urea cycle, carbamoyl phosphate synthase 1 (CPS1) is the first enzyme that is highly abundant in mitochondria, and it is expressed mainly in hepatocytes[39]. Pollettaet al[40] demonstrated that mitochondrial SIRT5 not only promotes ammonia detoxification by catalyzing desuccinylation of CPS1, but it also regulates glutamine homeostasis and ammonia levels by inhibiting glutaminase activity to reduce ammonia release and the conversion of glutamine to glutamate (Figure 2). Additionally, Zhanget al[41] conducted stoichiometry of lysine succinylation in mouse liver and found several highly succinylated lysine sites in arginine succinate synthetase (ASS1-a key enzyme in the urea cycle), which were regulated by SIRT5. Metabolomic analysis confirmed that SIRT5 deficiency reduced liver urea cycle activity, and more importantly, SIRT5 deficiency affected ammonia tolerance.
Influence of protein succinylation on lipid metabolism:The liver serves as an important regulator of lipid homeostasis[42], which includes lipid uptake, lipogenesis, fatty acid oxidation, ketogenesis, and lipid secretion[43]. When lipid synthesis exceeds lipolysis or export, it causes the accumulation of lipids in hepatocytes, ultimately leading to hepatic steatosis[32,44]. PRMT5 is a type II arginine methyltransferase that affects a variety of metabolites including phospholipids, fatty acids, and steroid hormones. Yuanet al[32] demonstrated that SIRT7-mediated desuccinylation of PRMT5 at K387 increases its methyltransferase activity, thereby upregulating lipid metabolism-related factors, such as sterol-regulatory element binding protein 1a (SREBP1a), FASN, ACACA, PPARγ, SCD,etc.Moreover, SIRT5 is also involved in the regulation of fatty acid β-oxidation[45]. When SIRT5 is deficient, fatty acid β-oxidation is reduced, which leads to fat accumulation in the liver[13].
Ketone bodies, which are comprised of acetoacetic acid, β-hydroxybutyrate, and acetone[46], are produced by the liver through fatty acid catabolism during glucose deficiency[47,48]. Mitochondrial 3-hydroxy-3-methylglutaryl-CoA synthetase 2 (HMGCS2) is a key enzyme required for ketogenic biosynthesis, which is regulated by succinylation[49]. Early studies on ketogenic regulation have shown that the accumulation of succinyl-CoA is the main process leading to enzyme inactivation in the liver. It was reported that glucagon drastically reduced succinyl-CoA levels and HMGCS2 succinylation, which led to strong ketogenic activation[4]. SIRT5 induces desuccinylation of HMGCS2 and promotes ketone body formation (Figure 2). Among the 15 succinylated lysine residues identified on HMGCS2, several sites appear to be highly targeted by SIRT5 including K83, K310, K350, K354, and K358[50]. Studies have shown that lysine adjacent to the HMGCS2 substrate binding site was strongly succinylated, suggesting that succinyl-CoA may interact with lysine residues around the catalytic pocket, resulting in non-enzymatic modification of these lysines[51,52].
At present, the discovery that various enzymes involved in liver glucose, amino acid, and lipid metabolisms were regulated by succinylation is only the tip of the iceberg, and whether other enzymes in the liver are modulated by succinylation remains to be ascertained.
Influence of succinylation on hepatic glucose metabolism:Under the stimulation of succinyl-CoA, HAT1 causes the K99 site of PGAM1 to be succinylated and promotes its enzyme activity, thus promoting glycolysis. The influence of succinylation on hepatic amino acid metabolism is shown by SIRT5 promotion of urea production by regulating the desuccinylation of ASS1 and CPS1. The influence of succinylation on hepatic lipid metabolism is shown by SIRT5-induced desuccinylation of HMGCS2 that promotes ketone body formation.
Succinylation in the progression of liver diseases
Several studies have established that succinylation is strongly associated with the progression of liver diseases, primarily for fatty liver disease, hepatitis, and hepatocellular carcinoma (HCC). Succinylation not only regulates fat deposition and thus fatty liver degeneration[45,53], but it also promotes HBV transcription and replication[31]. In addition, succinylation stimulates immune escape and tumor growth in HCC[54]. Therefore, the specific roles of succinylation in liver diseases are discussed herein.
Succinylation is involved in fatty liver disease:Fatty liver, which is caused initially by excessive fat accumulation in the liver, is a common chronic disease with a high prevalence worldwide[55,56]. As one of the metabolism-related posttranslational modifications, the succinylation degree is enhanced in fatty liver samples[45,57]. Chenget al[57] conducted quantitative succinylated proteome analysis using the livers of nonalcoholic fatty liver disease (NAFLD) rat models and identified 178 differentially succinylated proteins, which were involved in various metabolic and cellular processes and could promote the progression of NAFLD to varying degrees. Another study[45] also indicated that overexpression of SIRT5 in the liver resulted in decreased succinylation, enhanced fatty acid oxidation, and attenuated fatty liver degeneration. SREBP1, one of the transcription factors regulating hepatocellular lipogenesis, induces the expression of several lipogenic genes[58]. Guoet al[53] found that histone deacetylase 1 stabilized by P50 maintains SREBP1c activity through desuccinylation and promotes hepatic steatosis (Figure 3A). Yuanet al[32] also verified that SIRT7-mediated desuccinylation of PRMT5 at K387 promoted fatty liver by inducing arginine methylation of SREBP1a (Figure 3A). In summary, proteins with varied levels of succinylation may be potential targets for the treatment of fatty liver.
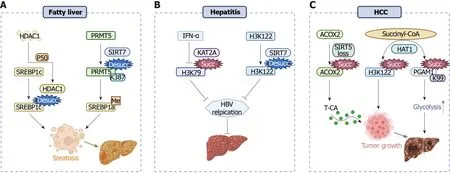
Figure 3 Succinylation affects the progression of fatty liver, hepatitis, and hepatocellular carcinoma. A: P50 stabilizes histone deacetylase 1 protein to keep desuccinylation of sterol-regulatory element binding protein 1c, thereby promoting fatty liver. Sirtuin 7 (SIRT7)-mediated desuccinylation of SIRT7 mediates the desuccinylation of arginine methyltransferase 5 at K387 promotes fatty liver by inducing arginine methylation of SREBP1a; B: IFN-α inhibits lysine acetyltransferase 2A-mediated succinylation of histone H3K79 and SIRT7 promotes desuccinylation of histone H3K122, which restrains viral replication and hepatitis; C: Sirtuin 5 deficiency activates acyl-CoA oxidase 2 succinylation, leading to elevated bile acid levels and promoting hepatocellular carcinoma (HCC). Histone acetyltransferase 1 not only promotes hepatocellular carcinogenesis by activating H3K122 succinylation but also promotes the glycolytic pathway by promoting succinylation of phosphoglycerate mutase 1 at K99, thereby promoting HCC. HBV: Hepatitis B virus; PRMT5: SIRT7 mediates the desuccinylation of arginine methyltransferase 5; SREBP: Sterol-regulatory element binding protein; ACOX2: Acyl-CoA oxidase 2; HAT1: Histone acetyltransferase 1; HCC: Hepatocellular carcinoma; PGAM1: Phosphoglycerate mutase 1.
Succinylation promotes hepatitis virus replication:Viral hepatitis is an infectious disease threatening human health, with a growing number of incidences in recent years[59]. HBV is a hepatotropic DNA virus that encodes multiple gene products for viral replication[60-62]. cccDNA plays an important role as a template for HBV transcription[63]. In the nucleus of HBV-infected cells, SIRT7 catalyzes the desuccinylation of cccDNA-bound histone H3K122, thereby limiting HBV transcription and replication[31]. KAT2A is identified as an important host factor for HBV replication[16]. Wanget al[15] confirmed that KAT2A is coupled to nuclear α-KGDHC, which acts as a histone H3 succinyltransferase. Later research[64] found that KAT2A can bind to cccDNA by interacting with the HBV core protein and catalyzing the succinylation of H3K79 on cccDNA (Figure 3B), thus promoting cccDNA transcription. Interestingly, Yuanet al[65] discovered that IFN-α restrains HBV cccDNA by downregulating KAT2A-mediated histone H3K79 succinylation. Collectively, targeting succinyl-modification enzymes and the succinylated proteins may provide new perspectives for the treatment of HBV.
Succinylation degree is associated with the progression of HCC:HCC is a common and highly lethal cancer, which ranks fourth in cancer incidence and second in cancer mortality[66,67]. In liver cancer patients, the expression of SIRT7 is significantly higher than that in normal liver tissues, and this initially increases at the first and middle stages of HCC but tends to decrease at the later stages[68]. Moreover, deficiency of SIRT5 promotes HCC and is associated with oxidative damage response[54]. Sunet al[54] showed that SIRT5 depletion led to increased lysine succinylation of acyl-CoA oxidase 2 (ACOX2) (Figure 3C), resulting in the synthesis of primary bile acids, which further promoted immune escape and tumor growth in HCC. In addition, Yanget al[17] confirmed that HAT1 promoted cell proliferation in HCC by catalyzing H3K122 succinylation (Figure 3C). Aspirin inhibits the succinylation level of PGAM1 at K99 by downregulating the expression of HAT1 and decreasing the level of glucose consumption and lactic acid production in liver cancer cells, thereby attenuating the glycolytic pathway in HCC[22,69]. In view of the complex roles of the succinylation signaling pathway in HCC, further studies are necessary to distinguish the pleiotropic effects of succinylation for its application in treating liver cancers.
CONCLUSlON
Through delineating the pleiotropic relationships between succinylation and hepatic metabolism, protein succinylation is involved in various physiological and pathological processes in the liver. Despite the significant progress in understanding this kind of post-translational modification, many issues remain unresolved, providing opportunities for future studies.
Succinylation is site-specific, and some proteins have several succinylation sites to make the substrate perform varied biological functions. Research has found that the 252 identified succinylated proteins have 1190 SuK sites and a total of 6579 lysines, with at least 18% of lysines on these proteins being modified by succinylation[49]. Whether these lysine succinylation sites overlap with known enzyme active sites may be an important sign to examine the function of succinylation regulation. Therefore, further research on the exact influences and mechanisms for succinylation on different proteins and/or different lysine sites of one target protein is of great importance.
Some specific succinylases regulate glycolysis and amino acid and lipid metabolisms by modifying the succinylation degree of critical enzymes. Are there other succinylases that are crucial for hepatic metabolism? Histone deacetylase 1 maintains SREBP1c activity through desuccinylation and promotes hepatic steatosis[53]. Similarly, some succinylationmodifying enzymes also exert other enzymatic activities. For instance, the demalonylation activity of SIRT5[27] and the acetylation activity of KAT2A are likely to contribute to regulating the biological processes of the liver. This suggests that some enzymes with other functions can also exert succinylation or desuccinylation activity, and some identified succinylases may act as other enzymes to participate in varied metabolic reactions.
In addition, succinylation-regulated metabolic processes could affect the progression of fatty liver, hepatitis, and HCC. In some cases, the effect of succinylation on disease development may not be common between histone and non-histone proteins. For instance, Yuanet al[32] verified that SIRT7-mediated desuccinylation of PRMT5 at K387 promoted fatty liver. Meanwhile, SIRT7 catalyzes the desuccinylation of cccDNA-bound histone H3K122, thereby limiting HBV transcription and replication[16]. This indicates that the roles of succinyltransferase/desuccinylase are not consistent with different metabolic environments or reactions. Therefore, we ask that the following scientific questions be resolved. What are the differences in succinylation levels and regulatory mechanisms during the occurrence and development of various metabolic diseases at different stages? How can we modulate more succinylation-related pathways in target tissues to improve human health?
In conclusion, the in-depth study of these issues would greatly enhance our understanding of protein succinylation, which further supports the theoretical basis for the treatment of metabolic diseases and the development of related drugs.
FOOTNOTES
Co-first authors:Shuang Liu and Rui Li.
Author contributions:Liu S and Li R wrote the original draft, created the figures, and revised the manuscript; Sun YW wrote the original draft and created the figures; Lin H supervised and verified the paper; Li HF supervised, conceived, verified, reviewed, and edited the manuscript; All authors were involved in the critical review of the results and have contributed to reading and approving the final manuscript. Liu S and Li R contributed equally to this work as co-first authors. The reasons for designating Liu S and Li R as co-first authors are twofold. First, the review was prepared as a collaborative effort with Liu S and Li R contributing equally to literature searching, draft writing, figure drawing, and manuscript revising. The designation of co-first authors authorship reflects the distribution of responsibilities and burdens associated with the time and effort required to complete the review and ensure effective communication and management of post-submission matters. Second, Liu S and Li R are skilled in different fields, which promotes the most comprehensive and in-depth discussion of the review topic, ultimately enriching reader understanding by offering various expert perspectives.
Conflict-of-interest statement:The authors declare that they have no conflicts of interest.
Open-Access:This article is an open-access article that was selected by an in-house editor and fully peer-reviewed by external reviewers. It is distributed in accordance with the Creative Commons Attribution NonCommercial (CC BY-NC 4.0) license, which permits others to distribute, remix, adapt, build upon this work non-commercially, and license their derivative works on different terms, provided the original work is properly cited and the use is non-commercial. See: https://creativecommons.org/Licenses/by-nc/4.0/
Country/Territory of origin:China
ORClD number:Shuang Liu 0000-0002-7654-9054; Rui Li 0009-0004-7385-8793; Ya-Wen Sun 0009-0009-5244-2322; Hai-Fang Li 0000-0002-2827-5925.
S-Editor:Qu XL
L-Editor:Filipodia
P-Editor:Zheng XM
杂志排行

World Journal of Hepatology的其它文章
- Update in lean metabolic dysfunction-associated steatotic liver disease
- Retrospective study of the incidence, risk factors, treatment outcomes of bacterial infections at uncommon sites in cirrhotic patients
- Palliative long-term abdominal drains vs large volume paracenteses for the management of refractory ascites in end-stage liver disease
- Comprehensive prognostic and immune analysis of sterol Oacyltransferase 1 in patients with hepatocellular carcinoma
- Prediction model for hepatitis B e antigen seroconversion in chronic hepatitis B with peginterferon-alfa treated based on a responseguided therapy strategy
- lnfluence of nonalcoholic fatty liver disease on response to antiviral treatment in patients with chronic hepatitis B: A meta-analysis