祁连山区青海云杉林碳汇特征及调控因子
2024-04-29裴薇薇杨喆王云英王新杜岩功
裴薇薇 杨喆 王云英 王新 杜岩功


摘要:祁连山区是我国西部重要的生态安全屏障和固碳场所。为准确评估祁连山区青海云杉林生态系统生长季碳汇特征,利用涡度相关技术并结合增强回归树模型与结构方程模型,研究生长季其碳通量变化特征及其环境影响机制。结果表明,青海云杉林生长季净生态系统碳交换(net ecosystem carbon exchange, NEE)日变化呈“V”型,CO2通量变化范围在-0.71~0.08 mg CO2·m-2·s-1,季节尺度NEE变化范围在-20.93~11.75 g C·m-2,月均碳吸收量(188.27±17.85) g·m-2,生长季累积碳吸收941.34 g·m-2。增强回归树模型揭示植被指数对净生态系统碳交换量相对贡献率最高,为50.3%,其次是净辐射,为15.9%。结构方程模型表明,植被指数与相对湿度对净生态系统碳交换量的直接作用系数分别为0.61与-0.17。多元逐步回归模型表明植被指数与相对湿度对NEE具有显著影响(R2=0.74, P<0.01)。随着植被指数增加,祁连山森林生态系统碳汇功能显著增强。结果为准确评估祁连山区青海云杉林生态系统碳汇能力提供科学依据。
关键词:涡度相关;青海云杉林;碳通量;环境因子
doi:10.13304/j.nykjdb.2022.0513
中图分类号:S718.55 文献标志码:A 文章编号:10080864(2024)01022608
自然生态系统碳汇是我国实现碳增汇的重要方式[12]。作为地球碳库的重要组成部分,陆地生态系统对全球碳循环至关重要。森林是陆地最大碳汇,约占全球植被碳储量的77%[34]。近年来,森林生态系统源汇效应及主要环境调控成为研究热点[5]。1980—1990年间北半球森林是年固碳6亿t的重要碳库,北美森林生态系统年固碳量0.7亿t[6],因气候环境差异,不同地区森林生态系统碳汇能力存在强异质性,欧亚大陆森林碳汇高于北美森林生态系统,美国森林碳汇能力强于中国北方森林系统。过去50年,我国森林年固碳量4.34 亿t[7],中国森林生态系统亦具有较强碳汇潜力[8]。
森林碳汇驱动因素由于环境异质性存在差异[910],如受气候[11]、地形[12]、林龄[13]等影响。亚马逊雨林受厄尔尼诺极端干旱气候影响,树木死亡率增加,碳汇能力降低[14];东北地区落叶松碳汇主要受光合有效辐射影响[15];台湾地区亚热带阔叶林生物碳最大值出现在地形平坦区[16];林龄是中国寒温带地区白桦林碳含量的主导因素[17]。温度是影响祁连山区青海云杉林碳储量的最关键因子[18],亦有研究表明降水是影响青海云杉林碳储量的主要因素。目前,对祁连山区青海云杉林碳汇特征及其影响因素的研究主要通过生物量估算,缺少基于涡度相关技术的准确评估研究。祁连山是我国西部地区重要生态安全屏障,也是冰川与水源涵养国家重点生态功能区。本研究利用涡动相关技术监测,揭示生长季青海云杉林生态系统碳通量动态变化特征及其主要影响因素,为准确评估祁连山区青海云杉林生态系统碳汇能力提供科学依据。
1 材料与方法
1.1 研究区概况及试验材料
研究区位于青海省门源县珠固乡祁连山国家公园寺沟管护站 (37°08′N, 102°22′E),海拔2 523 m。该区属高原大陆性气候,太阳辐射强,日温差较大,冷季漫长,暖季短暂。土壤类型是森林灰褐土,有机碳含量65.19 g·kg-1[19]。研究区平均气温-2.3~2.2 ℃,平均降雨量277.4~504.5 mm。选择长势均匀的青海云杉林作为研究对象,样地面积100 m × 100 m,共有活体树木1 493株。研究区植被是以青海云杉林为代表的寒温性针叶林,也是祁连山区主要的乔木建群种,包括青海云杉(Picea crassifolia)、祁连圆柏(Sabina przewalskii)、山杨(Populus davidiana)、桦木(Betula spp)等,树龄60~70 a,为次生林。
1.2 研究方法
1.2.1 森林净生态系统交换观测试验系统 祁连山国家公园寺沟管护站通量塔位于自然保护区内地势平坦、地形开阔处。试验系统主要由开路式涡度相关通量监测系统和微气象观测系统2部分组成,用于监测森林净生态系统碳交换(netecosystem carbon exchange, NEE)和微气象因子。开路式涡度相关系统安装在通量塔35 m处,包括一体化开路式涡动协方差系统(IRGASON, CampbellScientific, USA),集成了开路式CO2/H2O分析仪和三维超声风速仪为一体,可同时测量CO2 浓度、水汽密度、空气温度、大气压力、饱和水汽压差、三维超声风速和超声温度;数据采集器(CR6-XT,Camp-bell Scientific, USA),采样频率为10 Hz,以TOB3 格式收集并存储30 min 通量数据平均值。微气象观测系统主要包括:净辐射传感器(CNR4, Kipp&Zonen, USA),测量净辐射,同时有4 个输出,分别是向上的短波、向下的短波以及向上的长波和向下的长波;二维超声风速传感器(WINDSONIC 1405-PK-038, Gill, United Kingdom);大气压传感器(CS106,Vaisala, Finland);空气温湿度传感器(HMP155, Vaisala, Finland),可同时测量空气温度和相对湿度;翻斗式雨量筒(TE525MM, Texas, USA),收集降雨量。涡度相关监测系统和微气象观测系统需保持同一采样频率,得到时间为30 min 的平均CO2 湍流通量(Fc)。按照微气象学符号,向下通量(即碳吸收)用负值表示。
1.2.2 植被指数 植被指数(normalized differencevegetation index, NDVI)是影响碳通量的主要因素,因此本研究引入植被指数来揭示碳通量的调控因素[20]。本文使用数据为MODIS 合成产品MOD13Q1,空间分辨率为250 m, 时间分辨率为16 d,时间跨度选取2015至2020年的5—9月。数据来源于AppEEARS(https://lpdaac.usgs.gov/tools/appeears),并将NDVI像元初始值转化到-1~1。
式中,NIR、RED分别代表近红外波段、红外波段的反射率。
1.2.3 数据处理 本研究选用数据合计153 d(2020年5—9月),每天48组数据,产生理论数据7 344条,实际记录数据7 341条,数据覆盖率99.9%,其中有效数据量6 871条,有效率为93.5%,同时收集环境因子数据。
观测仪器于野外工作,会受到降雨、降雪等天气的影响,观测到的通量存在数据缺失和异常。对于数据质量控制通过在线软件EddyPro进行平面拟合、坐标旋转和WPL密度校正等。对于空缺数据,若缺失时间小于2 h采用线性内插法,超过则用平均昼夜插补法(mean diurnal variation,MDV),使用邻近一段时间内相同时段的观测平均值代替缺失值[21],白天选14 d,夜晚选7 d的时间长度。对于异常值处理,采用阈值确定来剔除不合理数值,森林通量阈值在-2~2 mg·m-2·s-1 比较合理;本文摩擦风速阈值为0.2 m·s-1。
1.2.4 统计分析方法 利用Excel 2010对生长季每30 min的NEE数值进行处理,对比分析生长季期间各通量日变化、月变化动态;采用Origin 2019软件进行回归分析和作图。本研究借助R4.0.5,利用结构方程模型(piecewiseSEM)和增强回归树模型(Dismo),选用气温、降水、净辐射、相对湿度、归一化植被指数作为自变量评价相对重要性。增强回归树模型设置学习速率为0.001,树的复杂度为5,每次抽取90% 的数据,并进行10次交叉验证,结果以百分数来表示单个预测因子的相对重要性。结构方程模型可对 Fishers C 值进行卡方检验并通过AIC(akaike information criterion)值判定模型整体拟合度。Fishers C 进行卡方检验时要求P>0.05,且AIC 越小,说明模型结构越合理。
基于2020年生长季时间序列,以环境因子为自变量,NEE为因变量构建多元线性回归模型。计算各项标准化回归系数。
Y=a+X1 P+X2 Ta+X3 RH+X4 Rn+X5 NDVI (2)
式中,a为残差,表示通量受环境因素影响所产生的部分微小波动;P、Ta、RH、Rn与NDVI分别为2020年降雨量、气温、相对湿度、净辐射与归一化植被指数。
2 结果与分析
2.1 青海云杉林生长季NEE 日变化特征
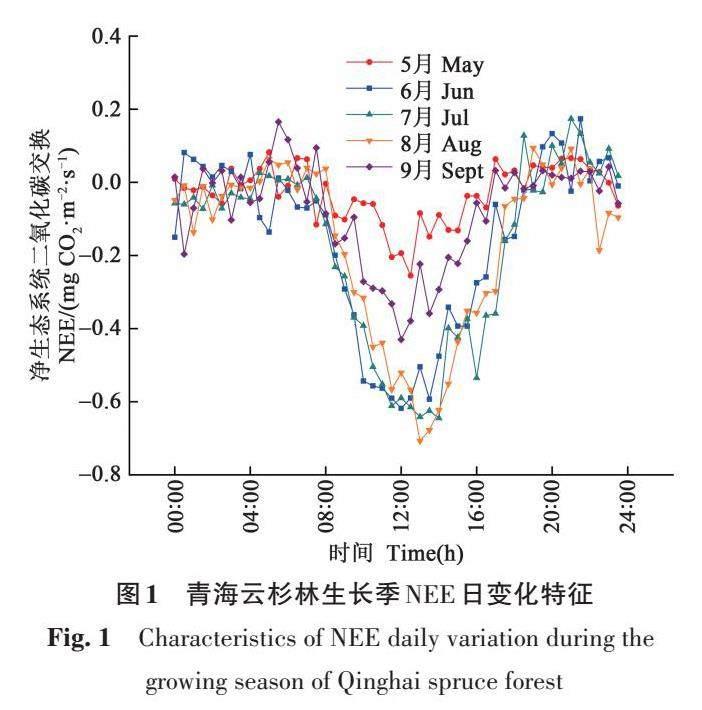
生长季青海云杉林NEE具有明显的脉冲式变化规律,总体呈“V”型曲线,白天和夜间分别为碳汇和碳源(图1)。从8∶00时开始,青海云杉林由释放CO2 转变为吸收CO2,12∶00时生态系统CO2 碳汇最高,约-0.71 mg CO2·m-2·s-1,17∶00时生态系统由吸收CO2 转化为释放CO2。夜间CO2 排放量与日间吸收量相比,明显较小且波动范围较窄。
在生长季5—9 月,青海云杉林NEE 每小时波动范围分别在-0.25~0.08、-0.62~0.17、-0.64~0.17、-0.71~0.09、-0.29~0.17 mg CO2·m-2·s-1,其中,7 和8 月变化幅度较高,而5、6 和9 月相对较弱。
祁连山区青海云杉林生态系统NEE 季节变化呈先增长后降低趋势。从5月19日开始碳汇能力增强,6月14日出现碳汇峰值-20.93 g C·m-2·d-1;从8月3日开始,碳汇开始降低且逐渐转为碳源,9月14日出现碳源峰值,达到11.75 g C·m-2·d-1。
生长季7 月5 日、8 月3 日和9 月5 日分别出现3 次强碳汇现象,其值分别为-20.63、-17.62 和-17.73 g C·m-2·d-1(图2)。5—9月碳通量平均值分别为(-1.50±0.08)(-8.15±0.14)(-8.92±0.12)(-8.16±0.12)(-4.02±0.15)g C·m-2·d-1。
2.2 青海云杉林生长季月尺度NEE 变化特征
生长季青海云杉林月均NEE 为(-188.27±17.85) g C·m-2,生长季NEE月累积量表现为7月(-276.67 g C·m-2·mon-1)>8月(-252.91 g C·m-2·mon-1)>6 月( -244.63 g C·m-2·mon-1)>9 月( -120.61g C·m-2·mon-1)>5月(-46.52 g C·m-2·mon-1),碳汇能力呈现先增加后降低趋势,7月碳汇能力最高,5月最低,最大值与最小值相差230.15 g C·m-2·mon-1(图3),6—8月碳吸收量显著高于5和9月(P<0.05),生长季NEE累积吸收总量为941.34 g C·m-2。
2.3 环境因素对青海云杉林生态系统NEE 的影响分析
增强回归树模型表明植被指数对NEE 相对贡献率最高,可以解释NEE 总变异的50.3%(图4),其次是净辐射,可以解释15.9%方差变异,二者累积贡献率达到66.2%,然后依次是气温(13.1%)>相对湿度(12.9%)>降雨量(7.7%)。结构方程模型结果表明,青海云杉林生长季NEE主要受植被指数和相对湿度调控(图5),直接影响系数分别为0.61(P<0.001)和-0.17(P<0.01)。气温对植被指数的直接作用系数为-0.57,达到极显著水平,对NEE间接作用系数为-0.35。2种方法均表明植被指数是影响青海云杉林生长季NEE的最主要正向调控因子。
2.4 基于多元线性回归模型的2015—2019 年生长季NEE 月变化特征
根据增强回归树模型与结构方程模型筛选出2个重要预测变量,即归一化植被指数与相对湿度,考虑引入并简化多元线性回归模型(式1),该模型R2 =0.75,矫正后R2 =0.74(P<0.01)。
Y=10.14-6.75NDVI-0.29RH (1)
由图6 可知,生长季月累积碳均值表现为2019 年(-413.56 g C·m-2·mon-1)>2018 年(-391.60g C·m-2·mon-1)>2015年(-333.53 g C·m-2·mon-1)>2016 年(-330.91 g C·m-2·mon-1)>2017 年(-323.40g C·m-2·mon-1)。其中,2018年变幅最大,为-318.59g C·m-2·mon-1,2019年最小,为-119.95 g C·m-2·mon-1。2015 年最大固碳量出现在5 月,2019 年出现在8月,其值分别为-390.38和-487.40 g C·m-2·mon-1,其余年份最大值皆出现在6月,与2020年最大固碳量出现在7月不同,但总体变化趋势基本一致,9月降至最低。
3 讨论
本研究结果表明,祁连山青海云杉林CO2通量日变化呈“V”型,固碳能力在12∶00左右达到最低,与其他森林生长季呈“U”型不同[2223],NEE变化趋势急促,是明显单谷峰型。究其原因,较温带森林所处位置,青海云杉林为高寒地带,昼夜温差较其他地区更大,白天光合作用强烈,对CO2吸收明显,夜间光合作用随气温下降迅速减弱,造成NEE波动范围更为剧烈。整个生长季NEE日变化特征仅有1个CO2吸收高峰,午间前后出现“午休”现象,可能是温度升高导致植物发生蒸腾作用而散失大量水分,气孔暂时关闭,导致植物在这段时间内CO2供应不足。与2020年相比,过去5年NEE的日变化较小,可能与祁连山区近10年气温与降水平均值略低有关。本研究结果也表明,气温与降水通过间接作用影响植被指数与相对湿度,从而达到影响NEE的效果。
青海云杉林生长季日变化范围在-20.93~11.75 g C·m-2·d-1,月累积平均值为-188.27 g C·m-2·mon-1;相同森林类型下,缙云山针阔混交林日变化范围在-26.91~15.74 g C·m-2·d-1,平均月累积为-100.64 g C·m-2·mon-1[24];凤阳山针阔混交林平均月累积为-50.2 g C·m-2·mon-1[25],相比之下青海云杉林固碳能力较强,可能是因为该地区固碳潜力相对较高[26],林龄[27]、蓄积量[28]、自然气候条件[11]等因素都可能影响针叶林生态系统固碳能力。综合各月碳通量总和与各月平均日变化可看出,碳通量总和月变化趋势与日变化趋势一致。
植被指数可以反映植被的生长状况,而植被生长状况是森林生态系统固碳能力的重要指标,青海云杉林生长季NEE 主要受植被指数调控,二者呈极显著正相关,说明植被生长状况越好,NEE就越高,这与张静茹等[29]的研究结果一致,说明植被自身生长状况是影响区域生态系统固碳能力的最主要因素。林冠是森林碳库的重要结构层次[30],植被指数最初也是从冠层结构来提取生长季植被动态变化,其中叶面积指数是量化森林生态系统冠层结构特性最常用的参数之一[31]。植被指数与叶面积指数之间具有线性关系,有研究通过监测林冠层的叶面积指数发现叶面积指数越大,植物固碳能力越强[32],也从侧面反映植被指数对森林NEE的正向影响。
净辐射的相对贡献较植被指数次之,但与NEE却无显著相关,这与纪小芳等[25]对凤阳山以及徐勇峰等[33]对湿地杨树林的研究结果不太一致。可能是由于祁连山是我国辐射资源最丰富的地区之一,长期处于强辐射环境下的森林植被经过长期适应具有较高的光合潜力。此外,在晴天的天气条件下,强烈的直接辐射也可能会使得植被光合作用出现光饱和现象,从而抑制光合效率[34]。也可能与下垫面性质有关,祁连山青海云杉林郁闭度较高,叶面积指数大,相较于其他森林对净辐射的吸收较低,因此作用不明显。气温也对NEE无显著影响,这与陈月明[35]对北美洲阔叶林与针叶林以及赵仲辉[36]对于亚热带杉木林生态系统的研究不同。温度升高会促进植物进入生理活动旺盛期,而青海云杉林处于大陆性气候,生长季均温仅12 ℃,并不足以对NEE产生强烈影响。但气温通过植被指数间接影响NEE并呈显著正相关,原因是生长季增温提高光合作用效率和水分利用率[37],植被生长加快使得植被指数增加间接增大NEE。
青海云杉林生长季NEE还受相对湿度调控,与NEE 呈显著负相关,这与Lu 等[38]研究结果不同。相对湿度不仅能够反映气温、降水等气候要素的综合影响,还可通过影响水汽凝结而改变降水发生概率。降水与气温是影响相对湿度的最主要因素。本研究中,降水对相对湿度的作用强于气温, 降水越大,空气水分含量越多,植物的光合作用随叶片含水量逐渐增强,导致NEE转换为负值,表现为碳汇,但青海云杉林作为干旱地区耐寒造林树种,受水分胁迫较小。
参 考 文 献
[1] 杨长进, 田永, 许鲜. 实现碳达峰、碳中和的价税机制进路[J].价格理论与实践, 2021(1):20-26, 65.
YANG C J, TIAN Y, XU X. The path of price and taxmechanism to achieve carbon peak and carbon neutrality [J].Price Theory Pract., 2021(1):20-26, 65.
[2] 王国胜, 孙涛, 昝国盛, 等. 陆地生态系统碳汇在实现“双碳”目标中的作用和建议[J]. 中国地质调查, 2021, 8(4):13-19.
WANG G S, SUN T, ZAN G S, et al .. Roles and suggestions ofterrestrial ecosystem carbon sink in achieving carbon emissionpeak and carbon neutrality in China [J]. Geol. Surv. Chin.,2021, 8(4):13-19.
[3] Food and Agriculture Organization of the United Nations(FAO). Global forest resources assessment(2020) [EB/OL]. [2022-05-22 ] . http://www.fao.org/.
[4] 尹晶萍, 张煜星, 付尧, 等. 中国碳排放与森林植被碳吸收潜力研究[J]. 林业资源管理, 2021(3):53-61.
YIN J P, ZHANG Y X, FU Y, et al .. The research of carbonemission and carbon sequestration potential of forest vegetationin China [J]. For. Resour. Manage., 2021(3):53-61.
[5] 谭丽萍, 刘苏峡, 莫兴国, 等. 华北人工林水热碳通量环境影响因子分析[J]. 植物生态学报, 2015, 39(8):773-784.
TAN L P, LIU S X, MO X G, et al .. Environmental controlsover energy, water and carbon fluxes in a plantation inNorthern China [J]. Chin. J. Plant Ecol., 2015, 39(8):773-784.
[6] AHLSTR?M A, RAUPACH M R, SCHURGERS G, et al .. Thedominant role of semi-arid ecosystems in the trend andvariability of the land CO2 sink [J]. Science, 2015, 348(6237):895-899.
[7] 国家林业和草原局. 中国森林资源报告(2014—2018) [M].北京:中国林业出版社, 2019.
[8] TANG X L, ZHAO X, BAI Y F, et al .. Carbon pools in Chinasterrestrial ecosystems:new estimates based on an intensive fieldsurvey [J]. Proc. Natl. Acad. Sci. USA, 2018, 115(16): 4021-4026.
[9] NAVE L E, SWANSTON C W, MISHRA U, et al ..Afforestation effects on soil carbon storage in the UnitedStates: a synthesis [J]. Soil Sci. Soc. Am. J., 2013, 77(3):1035-1047.
[10] MAKKONEN M, HUTTUNEN S, PRIMMER E, et al .. Policycoherence in climate change mitigation: an ecosystem serviceapproach to forests as carbon sinks and bioenergy sources [J].For. Policy Econ., 2015, 50:153-162.
[11] 周晓宇, 张称意, 郭广芬. 气候变化对森林土壤有机碳贮藏影响的研究进展[J]. 应用生态学报, 2010,21(7):1867-1874.
ZHOU X Y, ZHANG C Y, GUO G F. Effects of climate changeon forest soil organic carbon storage: a review [J]. Chin. J.Appl. Ecol., 2010, 21(7):1867-1874.
[12] 范叶青, 周国模, 施拥军, 等. 地形条件对毛竹林分结构和植被碳储量的影响[J]. 林业科学, 2013, 49(11):177-182.
FAN Y Q, ZHOU G M, SHI Y J, et al .. Effects of terrain onstand structure and vegetation carbon storage of Phyllostachysedulis forest [J]. Sci. Silvae Sin., 2013, 49(11):177-182.
[13] PUGH T A M, LINDESKOG M, SMITH B, et al .. Role of forestregrowth in global carbon sink dynamics [J]. Proc. Natl. Acad.Sci. USA, 2019, 116(10):4382-4387.
[14] BERENGUER E, LENNOX G D, FERREIRA J, et al ..Tracking the impacts of El Ni?o drought and fire in humanmodifiedAmazonian forests [J/OL]. Proc. Natl. Acad. Sci.USA, 2021, 118(30): 2019377118 [2022-04-22]. https://doi.org/10.1073/PNAS.2019377118.
[15] 李小梅, 张秋良. 兴安落叶松林生长季碳通量特征及其影响因素[J]. 西北农林科技大学学报(自然科学版), 2015,43(6):121-128.
LI X M, ZHANG Q L. Carbon flux and its impact factors oflarix gmelinii forest ecosystem during growing season [J]. J.Northwest. A&F Univ. (Nat. Sci.), 2015, 43(6):121-128.
[16] MCEWAN R W, LIN Y C, SUN I F, et al .. Topographic andbiotic regulation of aboveground carbon storage in subtropicalbroad-leaved forests of Taiwan [J]. For. Ecol. Manag., 2011,262(9):1817-1825.
[17] 魏红, 满秀玲. 中国寒温带不同林龄白桦林碳储量及分配特征[J]. 植物生态学报, 2019, 43(10):843-852.
WEI H, MAN X L. Carbon storage and its allocation in Betulaplatyphylla forests of different ages in cold temperate zone ofChina [J]. Chin. J. Plant Ecol., 43(10):843-852.
[18] 刘建泉, 李进军, 郝虎, 等. 祁连山青海云杉林生物量与碳储量及其影响因素分析[J]. 现代农业科技, 2017(12):140-143,146.
LIU J Q, LI J J, HAO H, et al .. Analysis on biomass andcarbon storage of Picea crassifolia forest in Qilian mountainsand its influence factors [J]. Mod. Agric. Sci. Technol.,2017(12):140-143,146.
[19] 马剑, 金铭, 敬文茂, 等. 祁连山中段典型植被土壤有机碳密度研究[J]. 中南林业科技大学学报, 2020, 40(8):99-105.
MA J, JIN M, JING W M, et al .. Study on soil organic carbondensity of typical vegetation in the middle Qilian mountains [J].J. Cent. South Univ. For. Technol., 2020, 40(8):99-105.
[20] WANG H, LI X, XIAO J, et al .. Carbon fluxes across alpine, oasis,and desert ecosystems in northwestern China:the importance ofwater availability [J/OL]. Sci. Total Environ., 2019, 697:133978[2022-04-22]. https://doi.org/10.1016/j.scitotenv.2019.133978.
[21] 周宇, 黄辉, 张劲松, 等. 森林生态系统涡度相关法碳通量长时间连续性缺失数据插补方法的比较[J]. 中国农业气象,2021, 42(4):330-343.
ZHOU Y, HUANG H, ZHANG J S, et al .. Comparison of gapfillingmethods for long-term continuous missing data in carbonflux observation by eddy covariance method of forest ecosystem [J].Chin. J. Agro. Meteorol., 2021, 42(4):330-343.
[22] 牛晓栋, 江洪, 张金梦, 等. 浙江天目山老龄森林生态系统CO2通量特征[J]. 应用生态学报, 2016, 27(1):1-8.
NIU X D, JIANG H, ZHANG J M, et al .. Characteristics of CO2flux in an old growth mixed forest in Tianmu mountain,Zhejiang, China [J]. Chin. J. Appl. Ecol., 2016, 27(1):1-8.
[23] 李润东, 范雅倩, 冯沛, 等. 北京松山天然落叶阔叶林生态系统净碳交换特征及其影响因子[J]. 应用生态学报, 2020,31(11):3621-3630.
LI R D, FAN Y Q, FENG P, et al .. Net ecosystem carbonexchange and its affecting factors in a deciduous broad-leavedforest in Songshan, Beijing, China [J]. Chin. J. Appl. Ecol.,2020,31(11):3621-3630.
[24] 王倩, 王云琦, 马超, 等. 缙云山针阔混交林碳通量变化特征及影响因子研究 [J]. 长江流域资源与环境, 2019, 28(3):565-576.
WANG Q, WANG Y Q, MA C, et al .. The characteristics andinfluencing factors of carbon fluxes in coniferous and broadleavedmixed forests in Jinyun mountain [J]. Resour. Environ.Yangtze Basin, 2019, 28(3):565-576.
[25] 纪小芳, 鲁建兵, 杨军, 等. 凤阳山针阔混交林碳通量变化特征及其影响因子[J]. 东北林业大学学报, 2019, 47(3):49-55.
JI X F, LU J B, YANG J, et al .. Carbon flux variationcharacteristics and its influencing factors in conifero us andbroad-leaved mixed forest in Fengyang mountain [J]. J. Northeast For. Univ. , 2019, 47(3):49-55.
[26] 吴振云, 李进军, 李娜, 等. 祁连山青海云杉林生长状况及固碳潜力空间分异研究[J]. 兰州大学学报(自然科学版),2020, 56(6):749-754.
WU Z Y, LI J J, LI N, et al .. Growth status of Qinghai spruceforest and its potential distribution of biomass C stock in Qilianmountains, Northwestern China [J]. J. Lanzhou Univ. (Nat.Sci.), 2020, 56(6):749-754.
[27] 冯源, 朱建华, 肖文发, 等. 干扰及林龄影响下迪庆州云杉老龄林生态系统碳储量动态[J]. 生态环境学报, 2017, 26(9):1465-1472.
FENG Y, ZHU J H, XIAO W F, et al .. Disturbances and ageingaffected carbon dynamics in old-growth spruce forest in Diqingprefecture [J]. Ecol. Environ. Sci., 2017, 26(9):1465-1472.
[28] 张颖, 李晓格, 温亚利. 碳达峰碳中和背景下中国森林碳汇潜力分析研究[J]. 北京林业大学学报, 2022, 44(1):38-47.
ZHANG Y, LI X G, WEN Y L. Forest carbon sequestrationpotential in China under the background of carbon emissionpeak and carbon neutralization [J]. J. Beijing For. Univ., 2022,44(1):38-47.
[29] 张静茹, 同小娟, 孟平, 等. 基于植被指数、叶绿素荧光和碳通量的华北山地人工林物候对比研究[J]. 北京林业大学学报, 2020, 42(11):17-26.
ZHANG J R, TONG X J, MENG P, et al .. Comparative studyon phenology in a mountainous plantation in northern Chinabased on vegetation index, chlorophyll fluorescence andcarbon flux [J]. J. Beijing For. Univ., 2020, 42(11):17-26.
[30] 田风霞, 赵传燕, 冯兆东, 等. 祁连山青海云杉林冠生态水文效应及其影响因素[J]. 生态学报, 2012, 32(4):62-72.
TIAN F X, ZHAO C Y, FENG Z D, et al .. Eco-hydrologicaleffects of Qinghai spruce (Picea crassifolia) canopy and itsinfluence factors in thw Qilian mountains [J]. Acta. Ecol. Sin.,2012, 32(4):62-72.
[31] 张艳丽, 费世民, 李智勇, 等. 成都市沙河主要绿化树种固碳释氧和降温增湿效益[J]. 生态学报, 2013, 33(12):3878-3887.
ZHANG Y L, FEI S M, LI Z Y, et al .. Carbon sequestration andoxygen release as well as cooling and humidification efficiencyof the main greening tree species of Sha River, Chengdu [J].Acta. Ecol. Sin., 2013,33(12):3878-3887.
[32] 刘志理. 东北典型森林叶面积指数的时空动态[D]. 哈尔滨:东北林业大学, 2015.
LIU Z L. Spatial and temporal dynamics of forests in northeasternChina [D]. Harbin: Northeast Forestry University, 2015.
[33] 徐勇峰, 季淮, 韩建刚, 等. 洪泽湖湿地杨树林生长季碳通量变化特征及其影响因子[J]. 生态学杂志, 2018, 37(2):322-331.
XU Y F, JI H, HAN J G, et al .. Variation of net ecosystemcarbon flux in growing season and its driving factors in a poplarplantation from Hung-tse Lake wetland [J]. Chin. J. Ecol.,2018, 37(2):322-331.
[34] 刘佳. 太阳辐射对黄河小浪底人工混交林生态系统碳、水交换的影响[D]. 北京:北京林业大学, 2014.
LIU J. Impacts of solar radiation on net ecosystem carbon andwater exchange in a mixed plantation in the Xiaolangdi area [D].Beijing: Beijing Forestry University, 2014.
[35] 陈月明. 北美洲阔叶林与针叶林生态系统碳水通量驱动因素分析[D]. 广州:华南农业大学, 2018.
CHEN Y M. Main drivers of carbon and water flux attemperature deciduous forests and evergreen needleleaf forestsin North American [D]. Guangzhou:South China AgriculturalUniversity, 2018.
[36] 赵仲辉. 亚热带杉木林生态系统与大气间的碳通量研究[D]. 长沙:中南林业科技大学, 2011.
ZHAO Z H. A study on carbon flux between Chinese firplanations and atmosphere in subtropical belts [D]. Changsha:Central South University of Forestry and Technology, 2011.
[37] 栾金凯, 刘登峰, 刘慧, 等. 汉江流域上游植被指数变化的影响因素分析[J]. 华北水利水电大学学报(自然科学版),2019, 40(1):46-54.
LUAN J K, LIU D F, LIU H, et al .. Analysis of the effectingfactors of vegetation index change in the upper reach ofHanjiang River basin [J]. J. North China Univ. Water Resour.Electr. Power (Nat. Sci.), 2019, 40(1):46-54.
[38] LU E, TAKLE E S. Spatial variabilities and their relationshipsof the trends of temperature, water vapor, and precipitation inthe North American regional reanalysis [J/OL]. J. Geophys.Res-Atmos., 2010, 115, D06110 [2022-05-22]. https://doi.org/10.1029/2009JD013192.
(责任编辑:胡立霞)
