HMGN3基因表达状态在骨肉瘤患者中的预后价值
2024-04-25蔡秀英曾慧意叶萍萍
蔡秀英,曾慧意,叶萍萍
(福建中医药大学附属泉州市正骨医院小儿骨科,福建 泉州 362000)
骨肉瘤(OS)又名成骨肉瘤,是儿童和青少年时期最常见的原发性骨恶性肿瘤之一,其发病率2~4人/(年·百万人),是一种致命的恶性肿瘤,5年生存率为50%~65%[1]。发生于青少年的OS,具有转移能力强、预后差的特点,是导致青少年死亡的第二大病因,也是青少年因病致残最常见的原因之一[2]。尽管20世纪70年代末随着新辅助化疗的应用使OS生存率得到大幅度提高,但仍有20%~30%患者需要接受截肢治疗,因此OS是一种预后较差的恶性肿瘤[3]。高迁移率族核小体结合蛋白 3(HMGN3)是高迁移率族蛋白N(HMGN)家族中的一员,研究表明HMGN可能密切参与恶性肿瘤的发生发展,并与恶性肿瘤的预后有关[4]。目前 HMGN3在肿瘤中的研究较少,一些研究表明HMGN3 mRNA在乳腺癌中的表达水平上调;而在子宫内膜癌中的表达水平被下调,说明HMGN3在不同肿瘤中可能具有不同的作用[5]。目前HMGN3在骨肉瘤中的表达状态及其预后价值尚未被报道,本研究拟从GEO数据库筛选OS表达芯片,并结合随访资料进行预后分析,以探讨HMGN3在骨肉瘤中的表达状态及预后。
1 资料与方法
1.1数据来源及筛选:以“Osteosarcoma”作为关键词在GEO公共数据库(https://www.ncbi.nlm.nih.gov/geo/)检索并获取相关基因表达谱数据。最终确定了GSE12865、GSE14359、GSE21257三个数据集作为数据分析来源。其中,GSE12865数据集是基于GPL6244平台获得的测序数据,包含了12例OS样本和2例正常人成骨细胞(HOB)样本的基因表达数据[6]。GSE14359数据集是基于GPL96平台获得的测序数据,包含10例原发骨肉瘤组织、8例骨肉瘤肺转移组织和2例HOB样本[7]。GSE21257数据集是GPL10295平台获得的测序数据,包含53例OS长期随访信息[8]。
1.2差异表达分析(DEA):利用R软件中的limma软件包,分别对GSE12865、GSE14359芯片的骨肉瘤样本和正常对照样本进行DEA,分别提取HMGN3基因的DEA结果,根据LogFC绝对值>1和校正P值(FDR)<0.05判断HMGN3基因表达状态。
1.3预后分析(PA):使用R软件中的“survival”和“survminer”两个包进行预后分析。
1.4统计学处理:采用R 4.2.0软件中的“limma”包进行差异基因表达分析,采用“survival”和“survminer”包等进行生存分析。三组计量资料采用单因素方差分析。Kaplan-Meier 法绘制生存曲线。COX比例风险回归模型用于评价HMGN3基因不同表达状态的预后价值,以风险比(HR)作为主要效应指标。
2 结果
2.1HMGN3基因差异表达分析结果:GSE12865、GSE14359两个数据集中,HMGN3基因均呈现高表达状态,差异具有统计学意义(P<0.05)。见表1。
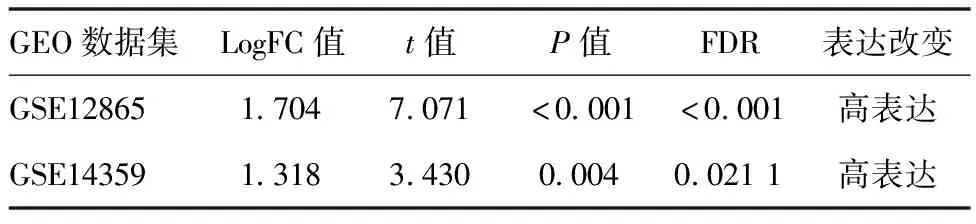
表1 HMGN3基因差异表达分析结果
2.2HMGN3基因的预后分析:HMGN3基因高表达组1年、3年、5年生存率分别为89.66%、50.98%、45.90%,而低表达组分别为95.83%、83.33%、74.56%,差异具有统计学意义(P<0.05)。Kaplan-Meier生存曲线见图1。HMGN3基因高表达患者死亡风险为HR=2.58,95%CI=(1.07,6.22),差异有统计学意义(P<0.05)。

图1 HMGN3基因在骨肉瘤中的生存曲线图
2.3不同转移状态,HMGN3基因的表达结果:GSE21257数据集中,根据是否发生远处转移及转移发生的时间,将53例样本分为无转移(19例)、初诊时转移(14例)以及后期转移(20例),无转移组HMGN3基因表达水平为14.73±3.17,初诊时转移组HMGN3基因表达水平为24.13±14.21,后期转移组HMGN3基因表达水平为19.03±9.25。三组患者HMGN3基因的表达水平比较差异有统计学意义(F=4.021,P<0.05),其中初诊时发生转移的骨肉瘤患者,其HMGN3基因的表达水平最高。
3 讨论
OS是青少年时期最常见的原发性骨恶性肿瘤,也是目前人类异质性最高、预后较差的实体肿瘤之一。OS在肿瘤确诊早期即有可能发生远处转移,因此,其病死率高且预后较差[2]。OS具有高度异质性,这种异质性不仅表现在病理学层面,而且还表现在基因组、转录和表观遗传水平上[9]。有研究表明,不同临床表型的OS,其基因表达谱存在显著差别[10]。OS可以发生于任何年龄,但以青少年人群发病率最高,其发生发展机制尚不明确[11]。转录组测序为探索OS肿瘤异质性以及肿瘤细胞与肿瘤微环境的相互作用提供了强有力的手段。近年来,GEO数据库的微阵列数据的共享,为揭示各类疾病发生发展的机制提供了便利[12]。
HMGN家族成员广泛存在于真核生物的细胞中,是含量最为丰富的非组蛋白家族,广泛参与细胞发育和细胞分化的调节[13]。研究表明,HMGN家族成员与肿瘤的发生发展密切相关[14]。如HMGN1参与小鼠肝癌的发生过程,并引起癌细胞表观遗传变化[15]。HMGN2 通过抑制连接组蛋白 H1与染色质调节区的结合,继而增强了STAT5对启动子的调控,最终促进乳腺癌的增殖和转移[16]。HMGN4也被证明参与了乳腺癌恶性行为的转化[17]。HMGN5通过FGF/FGFR信号通路促进直肠癌的增殖和侵袭[18]。HMGN3是HMGN家族的成员之一,主要功能是调节染色质的结构并通过转录因子调节转录[19]。HMGN3与多种癌症的发展有关,然而其潜在机制仍不清楚。Sorin等[20]发现,HMGN3的高表达与肝吸虫感染诱导的胆管癌(CCA)的转移特性相关。研究发现,胆管癌细胞中HMGN3的敲除显著降低了集落形成、迁移和侵袭的致癌能力。研究显示,HMGN3抑制CCA细胞中上皮调节基因(如CDH1/E-cadherin和TJAP1基因)的表达并阻断其胞内极性。染色质免疫沉淀测序分析显示,HMGN3通过与转录因子SNAI2结合并抑制上皮调节基因的表达,也可能通过抑制组蛋白脱乙酰酶(HDACs)抑制上皮调节因子重新激活,导致细胞迁移能力受损。因此,HMGN3过度表达通常以依赖于SNAI2基因和HDACs的方式抑制CCA细胞中上皮调节因子的转录并阻断其极性,从而提升了CCA细胞的致癌特性。目前HMGN3的生物学作用还有待继续探索。
本研究提示HMGN3基因参与了OS的疾病发生过程。Su等[21]研究发现,乳腺癌细胞中HMGN3 mRNA的表达水平比正常乳腺细胞上调,提示其可能与乳腺癌发生相关。李迎春[22]在肺癌中研究发现,与正常癌旁组织相比,肺癌组织HMGN3基因表达水平显著升高,并参与了肺癌的恶性行为,如促进迁移、侵袭及 EMT 过程。上述研究表明,HMGN3 是重要的致癌基因[23]。Zierau等[24]通过在小鼠子宫内膜癌细胞株研究发现,HMGN3在子宫内膜癌中表达下调,但这种低表达可能受miR-203表达调控,其具体机制还有待进一步研究。
目前关于HMGN3基因在恶性肿瘤中的预后价值尚未得到学者的报道,因此有必要开展研究探索HMGN3基因的预后价值。本研究提示,HMGN3基因是重要的癌基因,与其致癌特性相符,HMGN3基因参与了OS的发展过程。本研究为进一步开展免疫组化等检查等验证HMGN3基因在OS中的生物学作用提供了研究方向。
综上,HMGN3基因在OS中呈高表达状态,而且HMGN3基因高表达对OS患者的预后具有显著影响,但其具体机制,还需要进一步探索。
