IL-10通过上调socs3抑制衣原体感染细胞炎症因子表达
2024-04-09罗玲艳杜昆长江大学附属第一医院输血科荆州434000
罗玲艳 杜昆 (长江大学附属第一医院输血科,荆州 434000)
衣原体是一种全球普遍流行的性传播疾病的病原体,常常感染生殖道、肛门直肠和口咽部的黏膜上皮细胞。女性生殖道感染衣原体后,易引起输卵管性不孕及宫颈炎等并发症[1-3]。有研究表明,衣原体感染后能诱导机体产生大量炎症因子,如IL-6、IL-8、IL-12及TNF-α等,过度产生的炎症因子会对机体组织造成损伤[4-5]。因此,有效抑制炎症因子的产生可降低对机体组织的损伤。IL-10具有很强的免疫抑制能力,能防止过度炎症对机体组织造成的损伤[6]。IL-10与其受体结合后,通过激活JAK/STAT信号通路发挥作用。STAT3能诱导细胞因子信号转导抑制因子3(suppressors of cytokine signaling 3,SOCS3)蛋白产生,而SOCS3具有很强的抑制炎症因子产生的能力。因此,推测IL-10可能通过诱导SOCS3蛋白抑制衣原体感染细胞炎症因子的产生。本研究旨在探讨IL-10抑制衣原体感染细胞炎症因子的产生是否与SOCS3蛋白有关及其可能的机制,为衣原体感染防治工作提供实验依据。
1 材料与方法
1.1 材料 沙眼衣原体L2血清型和HeLa细胞均由中南大学余平教授惠赠;DMEM培养基购自Invitrogen;RT-PCR试剂盒购自碧云天生物技术有限公司;ELISA试剂盒购自武汉博士德生物公司;人IL-10购自RD公司;所有抗体及socs3siRNA均购自Santa Cruz公司;Stattic购自Merck公司。
1.2 方法
1.2.1 细胞培养与沙眼衣原体增殖 将HeLa细胞解冻后接种到50 cm2的细胞培养瓶中,DMEM培养基作为细胞培养液(含10%小牛血清)。细胞置于37 ℃、含5%CO2培养箱中培养,待细胞长成单层后用胰蛋白酶消化细胞并传代培养。将沙眼衣原体L2悬液加入长有细胞的培养瓶中,2 h后弃上清,加入含有2 mg/L放线菌酮的细胞培养液继续培养,感染48 h并收集细胞,反复冻融3次,3 000 r/min,10 min离心,吸取上清并于-80 ℃保存备用。
1.2.2 RT-PCR 提取培养细胞总RNA,然后按照试剂盒说明书进行逆转录。PCR扩增基因,socs3基因正向引物5'-ATGGTCACCCACAGCAAGTT-3',反向引物5'-CTTAAAGCGGGGCATCGTACTG-3';β-actin基因正向引物5'-AGGCTGTGCTGTCCCTCT-3',反向引物5'-TCCGGTGAGGAGGATGCG-3'。扩增条件:95 ℃预变性15 min;94 ℃变性15 s,55 ℃退火10 s,40个循环;最后72 ℃延伸30 s。
1.2.3 ELISA检测细胞因子 按试剂盒说明书进行。加100 μl标准品和已用样品稀释液稀释的待测样本到反应孔中,37 ℃反应90 min;吸弃液体,每孔加入100 μl相应抗体工作液,37 ℃反应60 min;洗涤3次,每孔加入100 μl ABC工作液,37 ℃反应30 min;洗涤5次,每孔加入90 μl TMB显色液,37 ℃避光反应15 min;每孔加入100 μl终止液,在450 nm处测定OD值。
1.2.4 Western blot 实验 提取细胞总蛋白,取80 μg蛋白加入2×SDS缓冲液中,100 ℃沸水中加热5 min,冷却后上样电泳;4 ℃、80 V转膜 2 h,5%脱脂奶粉封闭1 h;加入一抗4 ℃孵育过夜,TBST洗膜3次;加入HRP 标记二抗,孵育1 h,洗膜3次。最后用ECL发光试剂盒检测蛋白条带。
1.3 统计学分析 采用SPSS26.0软件统计分析数据,计量资料用±s表示,采用t检验,P<0.05表示差异有统计学意义。
2 结果
2.1 IL-10上调沙眼衣原体感染细胞socs3基因表达 检测外源性IL-10对STAT3的活化情况,结果如图1A所示,沙眼衣原体对STAT3活化不明显,但IL-10(10 ng/ml)能使磷酸化的STAT3蛋白增多。进一步研究在IL-10作用下socs3基因表达情况,结果如图1B所示,衣原体感染和未感染细胞,在IL-10(10 ng/ml)的作用下,socs3mRNA表达明显增强;但在抑制剂Stattic作用下,socs3mRNA表达明显降低。加抑制剂组和未加抑制组比较,差异有统计学意义(P<0.05)。
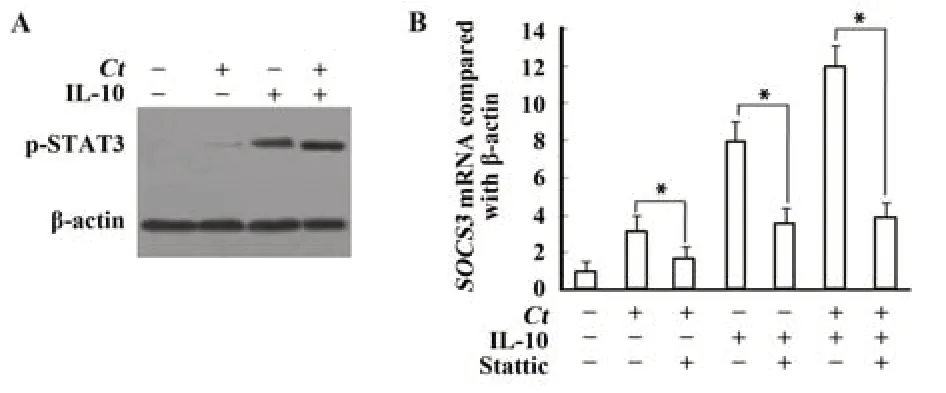
图1 沙眼衣原体通过活化STAT3蛋白上调socs3基因表达Fig.1 Chlamydia trachomatis up-regulates socs3 expression by activating STAT3 protein
2.2 沙眼衣原体感染诱导IL-6、IL-12表达 用不同滴度的衣原体感染细胞,在感染后不同时间点检测细胞培养上清中IL-6和IL-12产生情况。结果如图2A、B所示,沙眼衣原体能诱导感染细胞产生IL-6和IL-12,且随着感染时间的延长,产生的量越多;同时在感染相同的时间点,感染滴度越大,产生的量也越多。感染后12 h,不同滴度的衣原体诱导的IL-6和IL-12比较差异均有统计学意义(P<0.05);相同滴度的衣原体在感染不同时间诱导IL-6和IL-12比较差异也均有统计学意义(P<0.05)。

图2 沙眼衣原体感染诱导IL-6和IL-12表达Fig.2 Chlamydia trachomatis induces IL-6 and IL-12 expression in infected cells
2.3 IL-10通过诱导socs3抑制沙眼衣原体感染细胞IL-6、IL-12表达 用不同浓度的IL-10作用于衣原体感染细胞后检测IL-6和IL-12表达情况,结果如图3A所示,IL-10能明显抑制IL-6和IL-12产生,差异具有统计学意义(P<0.05)。由于IL-10能诱导socs3表达,而SOCS3蛋白具有抑制细胞因子产生的能力。采用RNA干扰抑制socs3表达,结果如图3B所示,抑制socs3表达后,IL-10对IL-6和IL-12的抑制作用明显减弱,差异具有统计学意义(P<0.05)。

图3 IL-10通过诱导socs3抑制沙眼衣原体感染细胞IL-6和IL-12表达Fig.3 IL-10 inhibits expressions of IL-6 and IL-12 in Chlamydia infected cells by inducing socs3
2.4 SOCS3蛋白抑制p38和ERK活化 利用RNA干扰抑制socs3基因转录,然后检测p38和ERK1/2活化情况。结果如图4所示,沙眼衣原体感染能活化细胞p38和ERK1/2信号通路,而IL-10(10 ng/ml)能抑制该信号通路;但当socs3基因转录被抑制时,IL-10抑制感染细胞p38和ERK1/2信号通路的作用明显减弱。表明IL-10能通过SOCS3蛋白抑制p38和ERK1/2通路活化。

图4 SOCS3蛋白抑制p38和ERK活化Fig.4 SOCS3 protein inhibits p38 and ERK activation
3 讨论
病原微生物入侵机体后,机体免疫系统会通过多种不同方式清除入侵的病原体。其中,炎症因子的产生是清除入侵病原体的重要因素之一[7]。然而,炎症因子产生过多,导致机体炎症过强,虽然有利于病原体的清除,但同时也会对机体组织造成不可逆性的损害[8-9]。衣原体具有二相性生活周期,其整个生活周期都必须依赖宿主细胞。女性生殖道感染沙眼衣原体后,会发生盆腔炎、宫颈炎等并发症,严重时甚至会发生输卵管性不孕[10-11]。因此,有效抑制衣原体感染后机体过强的炎症反应,能减少并发症的发生。
IL-10能抑制衣原体感染细胞炎症因子的产生,然而对其机制的报道较少[12-13]。IL-10与其受体结合后,主要通过JAK/STAT信号通路产生作用。为研究IL-10抑制衣原体感染细胞炎症因子产生的机制,课题组首先检测了IL-10对STAT的活化情况。结果表明,沙眼衣原体对STAT3活化并不明显,但IL-10能使磷酸化的STAT3蛋白增多,证实IL-10能有效激活STAT3蛋白。STAT3蛋白活化后进入细胞核内并调节许多基因的表达,其中socs基因是其重要的调节基因之一。为证实IL-10活化STAT3后是否调节socs3基因表达,本研究利用Stattic抑制STAT3活性,检测在STAT3活性被抑制后,socs3基因表达是否发生变化。结果显示,未加抑制剂时,在IL-10的作用下,衣原体感染和未感染细胞socs3基因表达明显上调;但当加入抑制剂时,感染和未感染细胞socs3基因表达明显降低;表明IL-10能通过STAT3蛋白诱导socs3基因表达。沙眼衣原体感染后能诱导感染细胞产生炎症因子,课题组也进一步研究了本实验中炎症因子产生情况,用不同滴度的衣原体感染细胞,在感染后不同时间点检测细胞培养上清IL-6和IL-12产生情况。结果显示沙眼衣原体能诱导感染细胞产生IL-6和IL-12,且呈时间和剂量依赖性,与以往的报道相同[5,14]。由于IL-10有很强的抑制炎症因子产生的能力,课题组继续研究了IL-10是否能够抑制衣原体感染细胞IL-6和IL-12的产生。课题组用3个不同浓度的IL-10作用于沙眼衣原体感染细胞,结果发现不同浓度的IL-10均能抑制IL-6和IL-12的产生,且随着IL-10浓度的增加,抑制作用越明显。课题组前期的研究显示IL-10能诱导socs3基因表达,而SOCS3蛋白具有抑制细胞因子产生的能力[15]。为证实IL-10抑制IL-6和IL-12产生的能力是否与SOCS3蛋白有关,本研究通过RNA干扰抑制socs3基因表达,然后检测socs3基因表达被抑制后,IL-10是否还能抑制IL-6和IL-12的产生。结果显示,socs3基因表达被抑制后,IL-10的抑制作用明显减弱。以上结果表明,IL-10有可能通过诱导SOCS3蛋白表达从而抑制衣原体感染细胞IL-6和IL-12产生。衣原体感染细胞后主要通过激活不同信号通路而诱导炎症因子的产生,如p38、ERK1/2等信号通路[16-17]。为研究IL-10是否可能通过诱导SOCS3蛋白影响细胞信号通路,从而影响炎症因子的表达。本研究通过RNA干扰抑制SOCS3蛋白表达,然后检测p38和ERK1/2活化情况。结果显示衣原体感染后,宿主细胞p38和ERK1/2信号通路被活化;当感染细胞预先加入IL-10时,该信号通路明显被抑制;但当socs3基因表达被抑制时,IL-10抑制感染细胞p38和ERK1/2信号通路的作用明显减弱。表明IL-10可能通过SOCS3蛋白抑制p38和ERK1/2通路活性。
总之,IL-10能活化STAT3蛋白并诱导SOCS3蛋白表达,进一步抑制p38和ERK1/2通路活化,从而抑制炎症因子的产生。这可能是IL-10抑制沙眼衣原体感染细胞炎症因子产生的机制之一。
