酶制剂与吡啶甲酸铬替代豆油对鲤鱼生长性能和饲料利用的影响
2024-04-02王子蔚王晓亮付大波张成杰吴金平吴建军
■ 王子蔚 王晓亮 付大波 张成杰 吴金平 刘 伟 吴建军*
(1.武汉新华扬生物股份有限公司,湖北 武汉 430206;2.农业农村部淡水生物多样性保护重点实验室,中国水产科学研究院长江水产研究所,湖北 武汉 430223)
鲤鱼是我国淡水养殖的主要经济鱼类,无论是养殖面积还是养殖产量在淡水养殖业中都占有较大的比重[1]。近年来,由于鱼粉、豆粕等优质蛋白原料和各种油脂等能量原料价格的不断上涨,鲤鱼的饲料成本和养殖成本也不断提高,另一方面,受整个经济大环境的影响,鲤鱼的销售价格呈现持续下滑的态势,导致这两年来,河南、辽宁、山东、河北等鲤鱼养殖区域不少养殖户纷纷转养或转行,鲤鱼养殖面积和养殖产量严重缩减。因此,降低鲤鱼饲料和养殖成本,提升养殖效益,已成为广大养殖户、饲料行业从业人员以及科研人员共同关注的问题。
在此背景下,植物性原料逐渐替代鱼粉、肉粉等动物性原料[2-6],成为鲤鱼饲料中的主要原料。然而植物性原料通常含有大量的木聚糖、纤维素、果胶、葡聚糖、甘露聚糖等非淀粉多糖类抗营养因子[7],这些抗营养因子是植物细胞壁的主要构成成分,它们将大量的营养物质包裹在细胞内,阻碍消化酶对营养物质的消化吸收,从而降低饲料的利用率[8]。随着生物工程和发酵工程的发展,使大规模工厂化生产用于降解各种非淀粉多糖的酶制剂成为可能[9]。研究表明,通过添加外源性非淀粉多糖酶,可以消除饲料中非淀粉多糖的抗营养作用,使饲料中更多营养物质得到消化吸收,从而提高饲料的利用率[10],有节约饲料中能量源原料添加的可能;且吡啶甲酸铬在促进鱼体的糖酵解供能上具有正向作用[11]。通过上述研究推知:酶制剂与吡啶甲酸铬均存在从能量节约角度替代鲤鱼饲料中能量源原料的可能,而有关酶制剂和吡啶甲酸铬对水产饲料中油脂类原料替代以及替代后对养殖鱼类生长性能和饲料利用的研究尚不多见。因此,本试验旨在研究通过在饲料中添加酶制剂和吡啶甲酸铬来提高鲤鱼对饲料中蛋白质、碳水化合物的利用率,从而减少饲料中油脂类原料的添加量,节约饲料成本和养殖成本,提高鲤鱼养殖的经济效益。
1 材料与方法
1.1 试验鱼及饲养管理
试验用鲤鱼为兴国红鲤(Cyprinus carpio var.singuonensis),购于武汉市正好养殖场,养殖试验在中国水产科学院长江水产研究所室内试验养殖车间进行。试验前先将试验鱼用高锰酸钾浸泡消毒后,置暂养池暂养2 周,投喂商品饲料以适应养殖环境。待试验鱼驯养稳定后,挑选大小一致、鳞片完整的平均体重为(7.53±0.02) g的健康鱼,随机分成7组,每组设3个重复,每个重复30 尾鱼,于直径30 cm、盛水100 L 的循环系统水缸中饲养。试验期为8 周,采用循环水控温养鱼,24 h 充气增氧,水温控制在(25±2) ℃,pH 控制在7.5~8.5,氨氮浓度控制在0.3 mg/L 以下,亚硝酸盐浓度低于0.1 mg/L。每天投喂2次,时间分别为09:00和16:00,投喂率为鱼体重的4%~5%。
1.2 试验饲料
以鱼粉、豆粕、菜粕、棉粕、小麦和米糠等为原料配制7 组饲料。HL 组为基础对照组,ML、LL 组为负对照组,分别减少0.5%、1.0%豆油的添加,MLE、LLE组为加酶组,MLEC、LLEC 组为加酶加铬组。试验酶制剂由武汉新华扬生物股份有限公司提供,酶谱组成为:木聚糖酶600 U/g,β-葡聚糖酶5 600 U/g,β-甘露聚糖酶400 U/g,纤维素酶1 600 U/g,中性蛋白酶1 000 U/g。吡啶甲酸铬(铬含量0.1%)由湖北神舟化工科技有限公司提供。试验饲料组成及营养水平见表1。原料粉碎后过60 目筛,按配方准确称量各种饲料原料,微量组分采用逐级扩大法混合,混合均匀后用小型环模颗粒机压制成粒径为2.0 mm的颗粒饲料,风干后用封口袋包装置于-20 ℃保存备用。
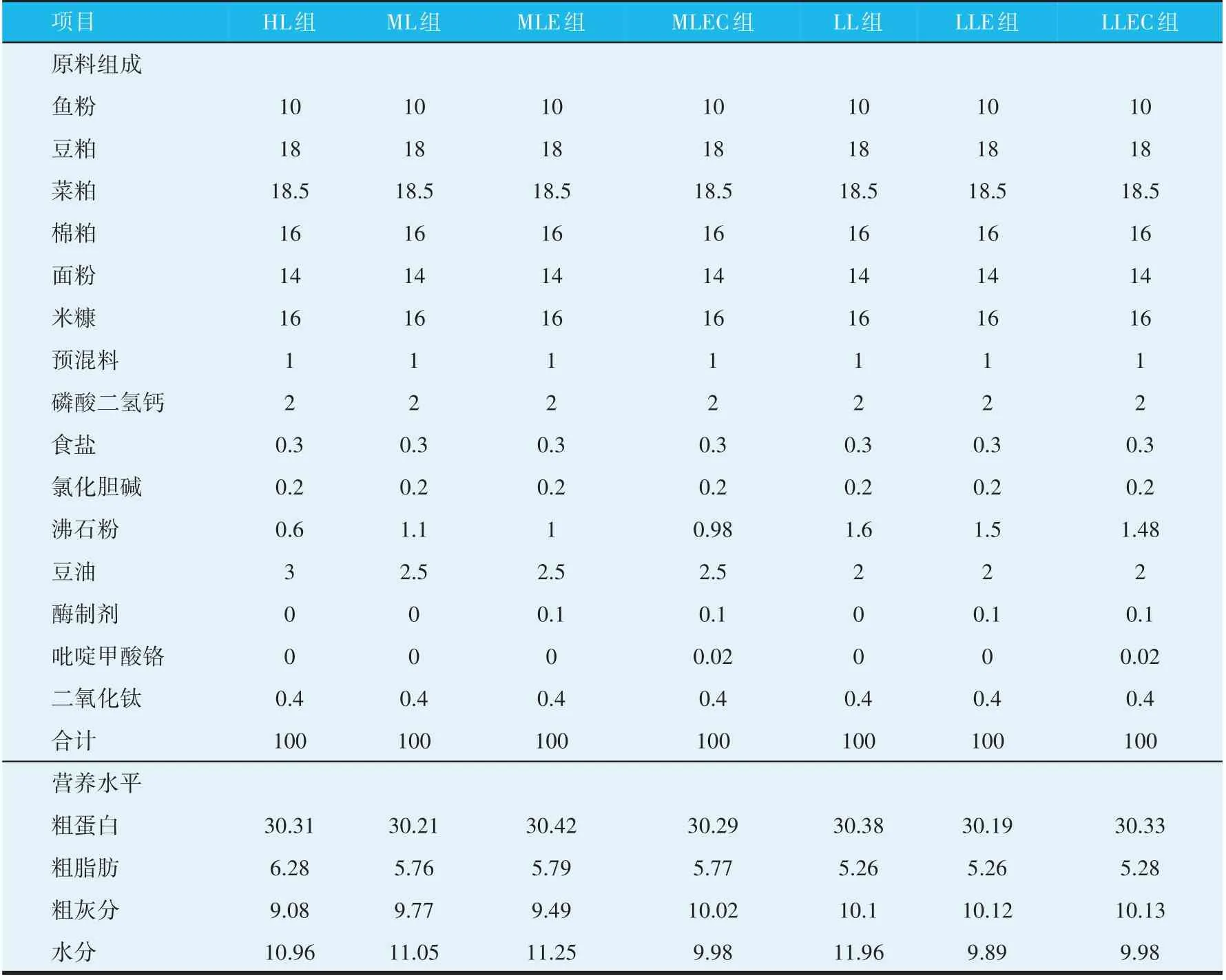
表1 试验饲料的原料组成及营养水平(干物质基础,%)
1.3 样品采集与指标测定
1.3.1 生长性能测定
试验开始时,准确称量并记录每个养殖桶中所放养鱼的尾数和重量,作为初始尾数和初始重量。试验过程中如出现死鱼情况,必须及时捞出死鱼并记录死鱼尾数和重量。试验结束后,停食24 h,以MS222 麻醉后,准确称量并记录每个养殖桶中所放养鱼的尾数和重量,作为终末尾数和终末重量(如出现过死鱼,需加上死鱼数量和死鱼重量),以进行生长指标计算。另在每个养殖桶取3 尾鱼测定体长、体重、肝重和内脏重,以计算鱼的肝体比、脏体比和肥满度。
增重率(WGR,%)=100×(终末均重-初始均重)/初始均重
特定生长率(SGR,%/d)=100×(ln 终末均重-ln 初始均重)/养殖天数
肥满度(CF,g/cm3)=100×平均体重/平均体长3
肝体比(HIS,%)=100×肝重/体重
脏体比(VSI,%)=100×内脏重/体重
成活率(SR,%)=100×初始尾数/终末尾数
1.3.2 饲料利用指标的测定
统计养殖期间饲料的投喂量、试验鱼的增重量和养殖天数,计算摄食量、饲料效率等指标。
饲料系数(FCR)=饲料投喂总量/(试验末鱼均重-试验初鱼均重)
摄食率(FI,%)=100×饲料投喂总量/[0.5×(初始尾数+终末尾数)×天数]
蛋白质效率比(PER,%)=100×(终末均重-初始均重)/(饲料投喂总量×饲料蛋白质含量)
1.3.3 表观消化率的测定
养殖试验开始后2 周,投喂2 h 后,等试验鱼排粪便时,用捞网收集水中的粪便,将粪便冷冻后置于真空干燥器中干燥,收集集中,并于-20 ℃冷藏备用。
饲料和粪便中的营养成分按《Official Methods of Analysis.17th ed., Association of Official Analytical Chemists, Washington, DC, USA》进行,即采用直接干燥法测定水分含量,凯氏定氮法测定粗蛋白含量,索氏抽提法测定粗脂肪含量,灼烧称重法测定灰分含量,二苯碳酰二肼丙酮法测定吡啶甲酸铬含量。最终计算获得饲料干物质、粗蛋白和粗脂肪的表观消化率。
干物质表观消化率(ADCDM,%)=100×(1-饲料吡啶甲酸铬含量/粪便吡啶甲酸铬含量)
粗蛋白表观消化率(ADCPro,%)=100×[1-(饲料中吡啶甲酸铬含量/粪便中吡啶甲酸铬含量)×(粪便蛋白质含量/饲料蛋白质含量)]
粗脂肪表观消化率(ADCFat,%)=100×[1-(饲料中吡啶甲酸铬含量/粪便中吡啶甲酸铬含量)×(粪便蛋白质含量/饲料蛋白质含量)]
1.3.4 血清生理生化指标分析试验结束时,经MS222麻醉后,每个养殖桶取3尾鱼,从尾部静脉采血,4 ℃冰箱静置2 h,以3 000 r/min的转速离心10 min,取上层血清用Sysmex 全自动生化分析仪(型号Chemix-800)测定血糖浓度、极低密度脂蛋白含量、高密度脂蛋白含量、三酰甘油浓度、胆固醇浓度、白蛋白浓度。肝糖原、肌糖原含量采用南京建成生物工程研究所有限公司试剂盒进行测定。
1.3.5 肝脏组织切片
养殖试验结束后,从每个养殖桶中取3 尾鱼,解剖后,取同一部位的肝脏,放入4%多聚甲醛溶液中固定24 h 后,用石蜡包埋,连续切片,H.E.染色,光镜下逐片观察及照片,运用IPP 6.0(Media Cybernetics Inc., Bethesda, MD, USA)图像分析软件对HE 染色结果进行分析、处理。
1.4 数据分析
试验结果用“平均值±标准差(mean±SD)”表示。数据采用SPSS 22.0 软件进行one-way ANOVA 分析,并Duncan’s 法作多重比较,以P<0.05 作为差异显著性判断标准。
2 结果与分析
2.1 不同脂肪水平饲料中添加酶制剂与吡啶甲酸铬对鲤鱼生长性能与形态学指标的影响
由表2 可知,在不添加酶制剂和吡啶甲酸铬时,随着饲料脂肪水平的降低,ML 组鲤鱼的增重率和特定生长率分别较HL 组显著降低了8.34%和5.99%(P<0.05),LL 组鲤鱼的增重率和特定生长率分别较HL 组显著降低了19.46%和13.77%(P<0.05)。在ML组中添加酶制剂后的MLE 组试验鱼的增重率和特定生长率与HL组无显著差异(P>0.05),但分别较ML组显著提高8.20%和5.73%(P<0.05);在LL 组中添加酶制剂后的LLE 组试验鱼的增重率和特定生长率较LL分别显著提高13.73%和9.03%(P<0.05),而较HL 组分别显著降低8.40%和5.99%(P<0.05)。在ML 组中同时添加酶制剂和吡啶甲酸铬后的MLEC 组试验鱼的增重率和特定生长率较HL 组分别显著提高8.59%和5.39%(P<0.05),较ML 组分别显著提高18.47%和12.10%(P<0.05);在LL 组中同时添加酶制剂和吡啶甲酸铬后的LLEC组试验鱼的增重率和特定生长率较HL 组分别降低5.90%和4.38%(P>0.05),较LL 组分别显著提高16.84%和11.11%(P<0.05)。LL 组和LLE组试验鱼的脏体比低于其他各组,而LLEC 组的脏体比则高于其他各组,HL 组、ML 组、MLE 组和MLEC 组各组之间的脏体比均无显著差异(P>0.05)。试验各组在肥满度、肝体比和成活率方面均无显著差异(P>0.05)。

表2 饲用酶制剂与吡啶甲酸铬对鲤鱼生长与形态的影响
2.2 不同脂肪水平饲料中添加酶制剂与吡啶甲酸铬对鲤鱼饲料利用的影响
由表3 可知,饲料中脂肪水平分别降低0.5%和1.0%时,ML 组和LL 组鲤鱼饲料的饲料系数较HL 组分别升高14.53%和13.37%(P<0.05),而在饲料中添加酶制剂后,MLE 组的饲料系数较ML 组降低了8.63%,LLE 组的饲料系数较LL 组降低了2.05%(P>0.05)。同时在饲料中添加酶制剂和吡啶甲酸铬后,MLEC 组的饲料系数较HL 组和ML 组分别降低了8.72%和20.30%(P<0.05),LLEC 组的饲料系数较LL组降低了8.72%(P>0.05)。ML 组和LL 组的蛋白质效率比较HL 组分别降低了12.50%和11.98%(P<0.05),而MLE 组和LLE 组的蛋白质效率比较ML 组和LL 组分别提高8.93%和2.96%(P>0.05),MLEC 组和LLEC组的蛋白质效率比较ML 组和LL 组分别提高25.00%(P<0.05)和10.65%(P>0.05),MLEC 组的蛋白质效率比最高。MLE 和MLEC 组的干物质、粗蛋白及粗脂肪的表观消化率均显著高于ML 组(P<0.05),LLE 组和LLEC 组的粗脂肪的表观消化率显著高于LL 组(P<0.05)。
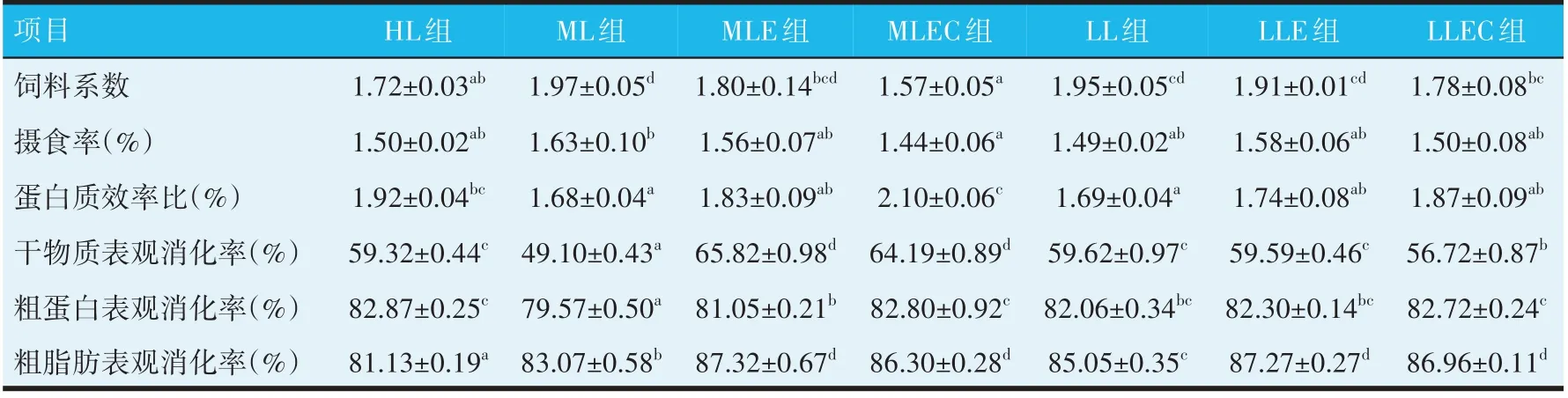
表3 饲用酶制剂与吡啶甲酸铬对鲤鱼饲料利用的影响
2.3 不同脂肪水平饲料中添加酶制剂与吡啶甲酸铬对鲤鱼饲料成本的影响
由表4可知,在不添加酶制剂和吡啶甲酸铬(ML、LL 组),饲料的脂肪水平分别降低0.5%和1.0%时,每千克鱼的饲料成本分别增加1.02 元和0.86 元,而投喂1 吨饲料的产鱼量分别减少71.38 kg 和66.42 kg。在ML组饲料中添加酶制剂或同时添加酶制剂和吡啶甲酸铬,每千克鱼的饲料成本较ML 组分别降低0.70 元和1.73元,较HL组增加0.32元和降低0.71元;投喂1吨饲料的产鱼量较ML 组增加47.87 kg 和127.67 kg,较HL组分别减少23.51 kg和增加56.29 kg;在LL组饲料中添加酶制剂或同时添加酶制剂和吡啶甲酸铬,每千克鱼的饲料成本较LL组分别降低0.15元和0.72元,较HL组分别降低0.71元和0.14元,投喂1吨饲料的产鱼量较LL组增加10.27 kg和51.15 kg,但较HL组分别减少56.15 kg和15.27 kg。
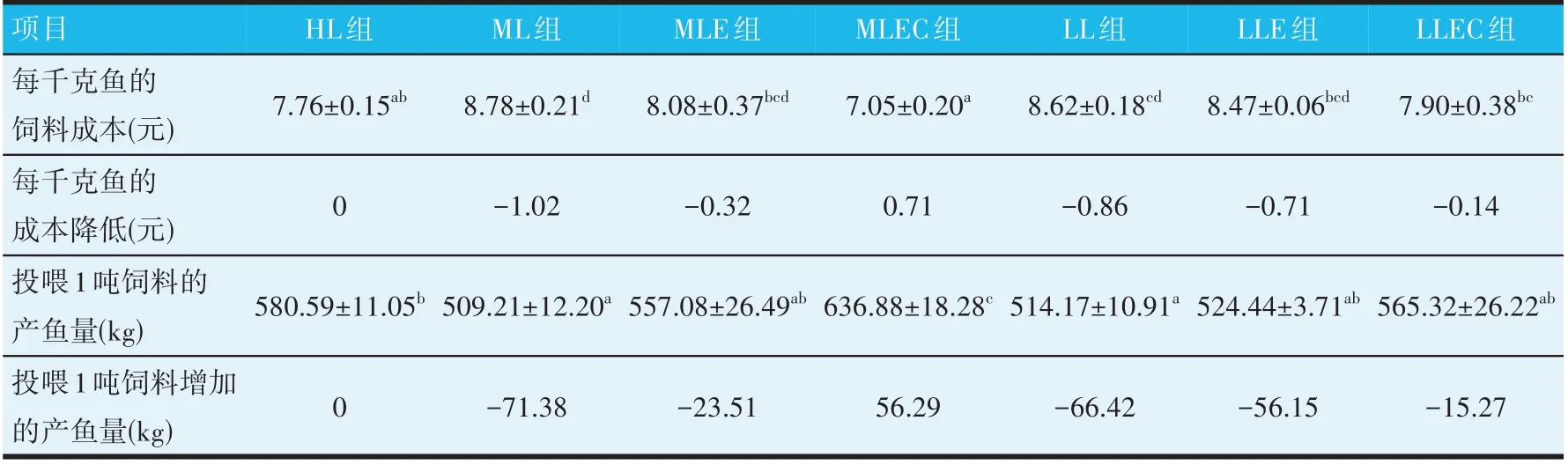
表4 饲用酶制剂与吡啶甲酸铬对鲤鱼饲料成本及产鱼量的影响
2.4 不同脂肪水平饲料中添加酶制剂与吡啶甲酸铬对鲤鱼血液生化指标的影响
由表5 可知,与HL 组相比,随着饲料脂肪水平的降低,ML 组和LL 组鱼肌糖原和肝糖原含量显著降低(P<0.05),而在ML 和LL 饲料中添加酶制剂后,MLE和LLE 组肌糖原含量较ML 组和LL 组分别提高43.30%(P<0.05)和12.50%(P>0.05),肝糖原含量较ML 组和LL 组显著提高12.17%和58.87%(P<0.05),同时添加酶制剂和吡啶甲酸铬后,MLEC 和LLEC 组肌糖原含量较ML 组和LL 组显著提高117.53%和54.76%(P<0.05),肝糖原含量较ML 组和LL 组分别提高31.80%和62.54%(P<0.05)。在ML 和LL 饲料中添加酶制剂后,MLE和LLE组极低密度脂蛋白的含量分别较ML 组和LL 组升高,而同时添加酶制剂和吡啶甲酸铬后,MLEC 和LLEC 组极低密度脂蛋白的含量又较MLE 和LLE 组显著降低(P<0.05)。各组试验鱼血糖浓度、高密度脂蛋白含量、三酰甘油浓度、胆固醇浓度及白蛋白浓度等均无显著差异(P>0.05)。
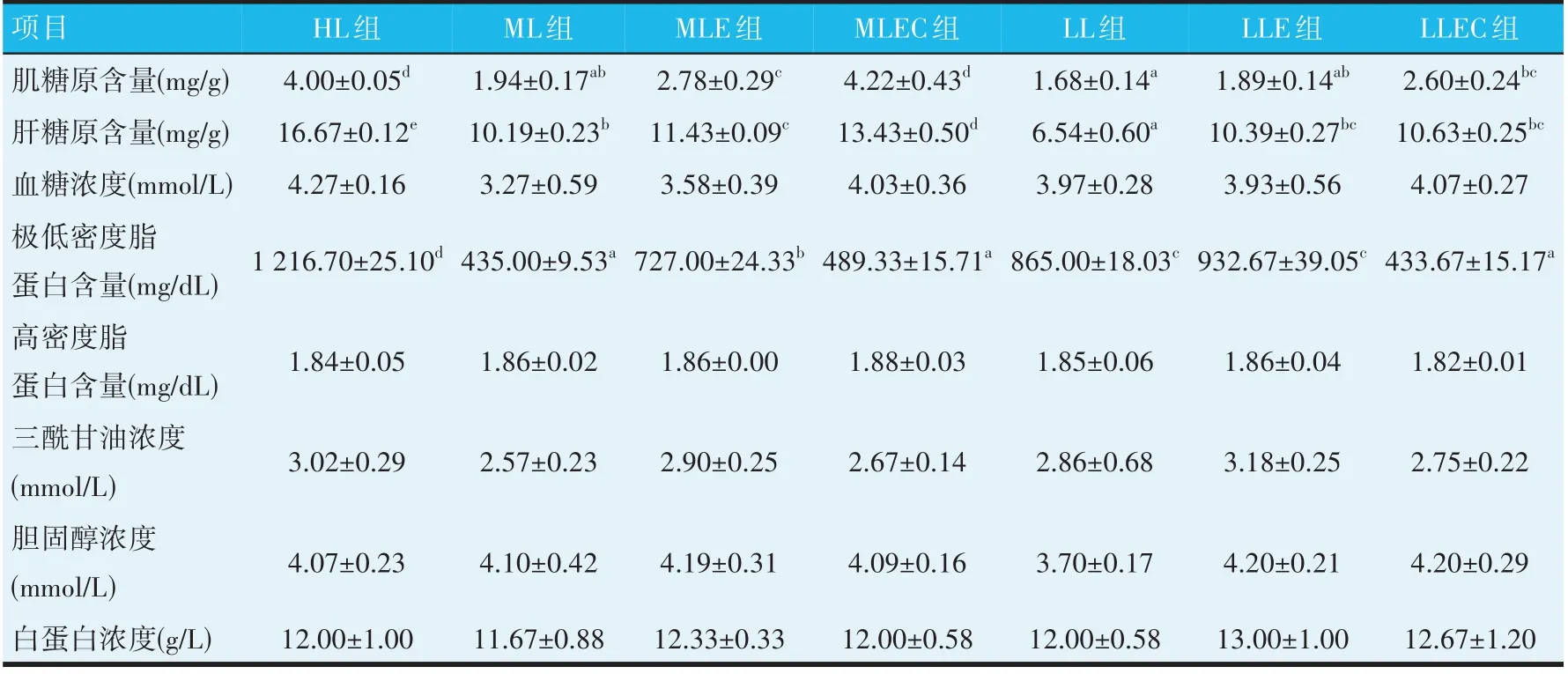
表5 饲用酶制剂与吡啶甲酸铬对鲤鱼血清生理生化指标的影响
2.5 不同脂肪水平饲料中添加酶制剂与吡啶甲酸铬对鲤鱼肝脏健康的影响
从图1 中可以看出,HL 组的试验鱼肝细胞核偏移,存在空泡变性的现象,逐步降低饲料脂肪含量后,ML 组和LL 组的核偏移和肝细胞空泡变性的情况得到明显改善,在饲料中添加酶制剂后,MLE 组和LLE组肝细胞空泡变性程度得到较大程度的改善,添加酶制剂和吡啶甲酸铬后,MLEC 组和LLEC 组肝细胞均排列整齐,大小均一,细胞膜分界明显,无核偏移和空泡变性现象出现,肝脏细胞健康。
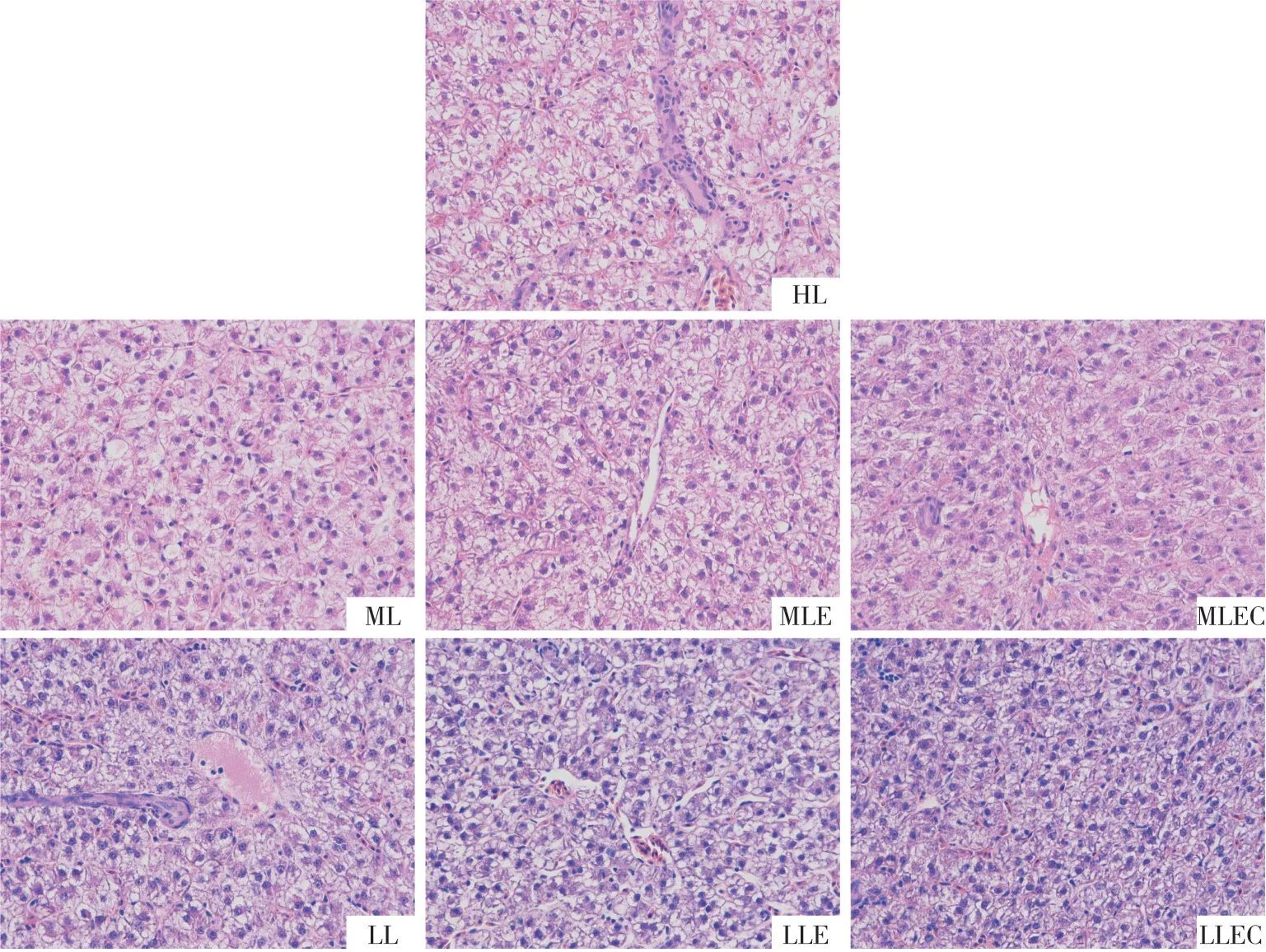
图1 饲用酶制剂与吡啶甲酸铬对鲤鱼肝脏健康的影响
3 讨论
3.1 酶制剂与吡啶甲酸铬替代豆油对鲤鱼的生长性能的影响
研究表明,在鲤鱼成鱼饲料中粗蛋白为30%左右时,推荐的饲料粗脂肪含量应为5%左右[12],本试验的饲料配方设计及营养水平符合该要求。当鲤鱼饲料中粗脂肪含量不足时,会出现增重率和特定生长率显著下降的情况[13]。文献指出,豆油中的粗脂肪含量为99%[14]。由此推测,作为水产饲料中的主要脂肪来源之一的豆油[15],其含量的减少将极大程度上对鲤鱼生长性能造成负面影响。
在本试验用饲料中,较基础对照组HL组而言,负对照组ML组和LL组分别减少0.5%和1.0%的豆油添加量,ML 组和LL 组的粗脂肪水平分别对应降低0.52%和1.02%。在本试验结果里,较HL 组而言,ML组和LL 组试验鱼的增重率分别下降8.34% 和19.46%,特定生长率分别下降5.99%和13.77%,证实了上述推测。
本试验中,在ML 组和LL 组的基础上仅添加酶制剂形成的MLE 组和LLE 组饲料对应饲喂的试验鱼的增重率的数值,虽然仍比基础对照组HL组低,但较其相应的负对照组分别提高了8.20%、13.73%,特定生长率较其相应的负对照组分别提高了5.73%、9.03%。这部分试验结果展现的比较趋势,与在黄羽肉鸡[16]和在鲫鱼[17]方面的研究结果一致。这意味着在饲料中添加酶制剂,可能在一定程度上弥补由饲料中低豆油添加量导致饲料粗脂肪水平降低对鲤鱼生长性能造成的负面影响。
本研究中,在ML 组和LL 组的基础上同时添加酶制剂和吡啶甲酸铬形成的MLEC 组和LLEC 组饲料对应饲喂的试验鱼,与其对应的负对照组、仅添加酶制剂组比较增重率和特定生长率的结果,在数值上均增加。据此推测,酶制剂和吡啶甲酸铬联合应用在鲤鱼饲料中替代豆油对鲤鱼生长性能的提高具有一定的协同作用。最近20 年的国内外多项研究表明,在饲料中添加酶制剂和吡啶甲酸铬能够有效促进营养物质的消化吸收和沉积,从而改善养殖动物的生长性能[18-23],佐证了我们的推测。
更有趣的是,MLEC 组的试验鱼的增重率和特定生长率甚至高于基础对照组HL 组,这说明酶制剂和吡啶甲酸铬联用能在不影响养殖鱼类生长性能的情况下,节约饲料中0.5%的脂肪水平。
3.2 酶制剂与吡啶甲酸铬替代豆油对鲤鱼的饲料利用程度及饲料成本的影响
在常规情况下,任何饲料都不会被完全消化,在动物饲料中添加外源酶制剂,一方面可以弥补动物内源消化酶分泌的不足,另一方面可以帮助降解动物自身不能消化利用的一些非淀粉多糖,从而提高饲料中营养物质的消化率及代谢能值[24]。而吡啶甲酸铬可以通过葡萄糖耐受因子协助或增强胰岛素在体内的功能作用而影响蛋白质、脂肪、糖等多种营养物质的代谢,尤其是在糖的吸收利用方面具有正向作用[25],因此可以推测,将二者联合应用,可能提高营养物质的吸收和利用,从而提升鲤鱼的饲料利用程度,降低养殖鲤鱼所需的饲料成本。
本研究中,与中脂饲料ML 组相比,MLEC 组在中脂饲料中添加酶制剂和吡啶甲酸铬后,其蛋白质效率比、粗蛋白表观消化率和粗脂肪表观消化率显著提高,饲料系数显著降低;与在中脂饲料中仅添加酶制剂的MLE 组相比,MLEC 组蛋白质效率比和粗蛋白表观消化率也有了显著提高,这说明酶制剂与吡啶甲酸铬联用替代豆油的确达到了协同增效的作用,提升了鲤鱼的饲料利用程度。
在水产养殖过程中,每千克鱼的饲料成本和每吨饲料的产鱼量是决定养殖效益高低的两个重要因素,也是养殖从业人员最为关注的两个指标。本研究中,在中脂饲料ML 组基础上同时添加酶制剂和吡啶甲酸铬的MLEC 组,每千克鱼的饲料成本较高脂饲料HL 组降低0.71 元,而每吨饲料的产鱼量较高脂饲料HL 组增加56.29 kg。由此可知,酶制剂与吡啶甲酸铬联用替代豆油确实能够提升鲤鱼的饲喂每吨饲料的产鱼量,降低了养殖鲤鱼所需的饲料成本。
综上,将酶制剂与吡啶甲酸铬联用替代豆油,能够提升鲤鱼的饲料利用程度,降低养殖鲤鱼所需的饲料成本,证实了上述推测。
3.3 酶制剂与吡啶甲酸铬替代豆油对鲤鱼血清生理生化指标的影响
血清生理生化指标能反映鱼类的生理代谢状态,与营养状况密切相关,生化指标的变化可以阐明营养素在机体代谢变化的作用机制[26]。糖原是葡萄糖在动物体内的一种储存形式,它们在维持血糖稳定、体内能量循环及物质代谢等方面具有重要的作用[27-28]。在本试验中,与中脂饲料ML 组和低脂饲料LL 组相比,同时添加酶制剂和吡啶甲酸铬的MLEC 组和LLEC 组,均显著提高了肌糖原和肝糖原的含量,这说明酶制剂和吡啶甲酸铬的使用,可能促进了饲料中碳水化合物的消化、吸收和沉积,从而提高了营养物质的利用效率。造成这种效果可能是饲料中添加了吡啶甲酸铬后,铬作为葡萄糖耐量因子的组成成分,通过增强胰岛素的作用而增强了肝糖原与肌糖原的合成。有趣的是,Brooks 等[29]有关鸡的研究的结果,与我们的推论相似。
极低密度脂蛋白是内源性三酰甘油体内运输的重要载体[30],即三酰甘油是极低密度脂蛋白的重要组成成分,而葡萄糖可提供生成三酰甘油所需要的甘油[31]。在本试验中,与中脂饲料ML 组和低脂饲料LL组相比,使用添加了酶制剂的MLE组和LLE组饲料饲喂鲤鱼后的血清中血糖浓度、极低密度脂蛋白浓度均提升,变化趋势一致。由此可以推测,添加酶制剂替代豆油能通过调节血清中血糖浓度方式调节极低密度脂蛋白浓度,提升鱼体内源性三酰甘油运输效率,从而提高了营养物质的利用效率。而令人意外的是,与中脂饲料ML 组和低脂饲料LL 组相比,使用添加了酶制剂的MLE 组和LLE 组饲料饲喂鲤鱼后的血清中三酰甘油浓度、极低密度脂蛋白浓度变化趋势也一致,结合血糖浓度同样的变化趋势,推测这可能是由于饲料中酶制剂的添加促进鱼体血糖生成过多,进一步转化成三酰甘油导致的[32]。
脂蛋白酯酶会从极低密度脂蛋白中夺取三酰甘油,用来储存或制造能量[33]。与中脂饲料加酶组(MLE 组)和低脂饲料加酶组(LLE 组)相比,使用同时添加了酶制剂和吡啶甲酸铬的MLEC 组和LLEC组饲料饲喂鲤鱼后的血清中三酰甘油浓度和极低密度脂蛋白浓度均下降,变化趋势一致。结合血清中血糖浓度、肝糖原含量、肌糖原含量升高的结果,表明酶制剂与吡啶甲酸铬联用可能通过增强胰岛素的作用进一步利用由酶制剂产生的高血糖影响,促进外源性糖转化为内源性糖原沉积,降低鱼体内源性糖酵解供能压力;推测还可能降低葡萄糖通过磷酸戊糖途径转化生成三酰甘油的效率,从而降低鱼体由于过高的脂肪含量导致的脂肪性肝损伤等疾病的可能[34]。
3.4 酶制剂与吡啶甲酸铬替代豆油对鲤鱼肝细胞组织形态的影响
肝胰脏是鱼类重要的代谢器官,在集约化养殖过程中,饲料中的脂肪水平过高常会引起养殖鱼类脂肪肝病的发生[35]。在本试验中,投喂高脂饲料HL 组的鲤鱼肝细胞核偏移,存在空泡变性现象,表明投喂高脂饲料可能会对鲤鱼的肝脏健康造成负面影响。Ye等[36]有关高脂饲料投喂草鱼的研究的结论,与本推测一致。
酶制剂的添加可能有利于改善肝脏的健康状况[37]。本试验中,与高脂饲料HL 组相比,将高脂饲料HL 组的脂肪水平分别降低0.5%和1.0%后形成的中脂饲料ML 组与低脂饲料LL 组分别饲喂鲤鱼,试验鱼的肝脏空泡面积明显减少。在中脂饲料ML组和低脂饲料LL 组中分别添加了酶制剂后,由对应的MLE 组饲料与LLE 组饲料饲喂的试验鱼的肝细胞中空泡化面积进一步显著降低,细胞质面积明显增加,说明酶制剂的添加确实有利于改善肝细胞组织形态,证实了上述推论。
本研究里,用在低脂饲料LL 组中同时添加了酶制剂和吡啶甲酸铬形成的LLEC 组饲料饲喂鲤鱼,肝细胞中空泡化面积较HL 组、LL 组和LLE 组都有降低,细胞质面积都有明显增加,此结果表明酶制剂与吡啶甲酸铬替代豆油降低鲤鱼饲料1%粗脂肪水平可能有利于改善肝细胞组织形态,且酶制剂与吡啶甲酸铬联用替代豆油对改善肝细胞组织形态可能具有协同作用。
本试验中,将中脂饲料ML 组添加酶制剂和吡啶甲酸铬形成的MLEC 组饲料饲喂鲤鱼,肝细胞中空泡化面积较HL组、ML组有降低,细胞质面积都有增加,此结果表明酶制剂与吡啶甲酸铬替代豆油降低鲤鱼饲料0.5%粗脂肪水平可能有利于改善肝细胞组织形态。但与由MLE 组饲料饲喂的鲤鱼相比,MLEC 组鲤鱼的肝细胞中空泡化面积升高,细胞质面积减少。由此推测,在鲤鱼饲料中由于低豆油添加量降低0.5%的粗脂肪水平的情况下,加入等同于降低部分豆油质量分数4%的吡啶甲酸铬与质量分数20%的酶制剂联用,对肝细胞组织形态的影响可能具有拮抗作用,加入的等同于降低部分豆油质量分数4%的吡啶甲酸铬相对于降低鲤鱼饲料0.5%的粗脂肪水平来讲可能过量。这可能是由于过量的吡啶甲酸铬对细胞DNA 的负面作用导致的[38]。
4 结论
酶制剂和吡啶甲酸铬联合应用在鲤鱼饲料中替代豆油,是具有一定的协同作用的,且能节约饲料中0.5%的脂肪水平。酶制剂和吡啶甲酸铬联合应用在鲤鱼饲料中替代豆油,没有负面影响甚至提高鲤鱼生长性能,提升鲤鱼的饲料利用程度,降低养殖鲤鱼所需的饲料成本,改善鲤鱼肝细胞组织形态。
