荒漠草原植物在氮磷添加下叶功能性状特征及其可塑性
2024-03-22鲍平安邱开阳黄业芸王思瑶崔璐瑶骆欣怡杨云涛谢应忠
鲍平安,邱开阳*,黄业芸,王思瑶,崔璐瑶,骆欣怡,杨云涛,谢应忠
(1. 宁夏大学林业与草业学院,宁夏 银川 750021;2. 宁夏大学宁夏草牧业工程技术研究中心,宁夏 银川 750021;3. 宁夏大学农业农村部饲草高效生产模式创新重点实验室,宁夏 银川 750021;4. 宁夏大学西北土地退化与生态恢复国家重点实验室培育基地,宁夏 银川 750021)
植物功能性状是植物体生命周期中获取、利用和保留资源等一系列活动的关键属性,它可以通过单独或者协同作用使植物生长策略对环境变化做出响应,还可以反映生态系统功能和过程的变化[1]。可塑性是植物为适应环境变化在形态和生理上做出改变的具体体现,可视为不同性状的协调响应,功能性状可塑性在植物对全球变化的响应中起决定性作用[2]。植物能够改变自身功能性状及时调整适应策略以抵抗环境的干扰,如气候、地形、土壤等环境因子的变化,因此,植物功能性状具有很强的可塑性,能够起到良好的生态指示作用[3-4]。
叶片是植物地上部分获取资源的主要器官,在陆地生态系统中起着至关重要的作用[5],不仅能产生植物生长发育所需的营养物质,还是植物和外界环境资源交换的基础[6],其形态性状和组织化学性状是决定植物生理和生物地球化学循环的重要因素[7]。其中,比叶面积与植物光合作用的净同化速率密切相关,比叶面积大的植物生长速率相对较快[5];叶片干物质含量是对植物叶组织密度的估计,干物质含量高的植物通常具有较高的资源储存能力[8];植物氮含量能够很好地反映土壤的营养状况[9];植物的碳氮比和碳磷比均能反映植物的生长状况,与植物生长速率呈负相关关系[10]。
氮(N)和磷(P)是影响植物地上地下生理生态过程的重要营养元素[11],也是植物根系从土壤中吸收的大量矿质元素,而且在生态系统中,N 和P 也是限制植物生长的重要因子。外源N、P 输入可以通过改变陆地生态系统的养分供应状况影响植物的形态、生理和化学性状[12]。研究表明,外源N 输入会使植物比叶面积和干物质含量发生显著的变化,随着N 添加量的增加,比叶面积随之增加,干物质含量下降[13],植物叶片N 含量增加[14-15],且N 添加也影响植物对P 的吸收[16]。因此N、P 的输入可能会通过影响植物功能性状进而对生态系统功能产生较大的影响。通过了解不同N、P 添加条件下植物群落叶片功能性状的可塑性响应,对预测植物和生态系统对全球变化因素的反应,认识异质生境下植物的生存适应策略至关重要。
荒漠草原是我国北方面积较大的一类草原,其干旱少雨、植被稀疏、水资源极度短缺,极易受到外界环境变化的影响[17]。目前,针对荒漠草原不同养分添加对植物功能性状的影响报道较少,N、P 添加对植物功能性状的作用机制尚不明确。因此,在大气N、P 沉降逐年加剧的背景下研究荒漠草原植物功能性状对N、P 添加的响应机制是有必要的,这对揭示荒漠草原植物对全球变化特别是氮沉降的反馈策略和适应机制具有重大生态学意义。为此,本研究以宁夏盐池荒漠草原为对象,通过分析不同N、P 添加处理下各个植物群落功能性状的变化情况,有助于从功能性状的角度理解变化环境下植物的响应、适应策略。
1 材料与方法
1.1 研究区概况
研究区位于宁夏回族自治区盐池县四墩子行政村荒漠草原(37°47′ N, 107°25′ E)。属于典型温带大陆性气候,年均气温7.7 ℃,≥0 ℃积温为3430.3 ℃。年平均降水量在250~350 mm,年蒸发量约2136 mm,年均无霜期162 d。土壤类型以灰钙土为主。主要植物有短花针茅(Stipa breviflora)、蒙古冰草(Agropyron mongolicum)、赖草(Leymus secalinus)、牛枝子(Lespedeza potaninii)、猪毛蒿(Artemisia scoparia)等。
1.2 样地设置
本研究采用随机区组设计,设置了4个N、P 养分添加处理,分别为对照(CK)、N 添加处理、P 添加处理和N+P 添加处理,每个处理3个重复,共计12个试验小区,每个小区面积为3 m×4 m,相邻小区间隔1 m。其中,N、P添加浓度水平参考中国科学院内蒙古草原生态系统定位研究站自2000 年设置的长期N、P 添加试验平台[18]的试验处理设置的。N 添加选用尿素(CH4N2O),P 添加选用过磷酸钙[Ca(H2PO4)2H2O],添加量分别为10 g·m-2·a-1和8 g·m-2·a-1。施用时将每个试验小区年施用量平分成2 等份,于6、7 月的月初选择雨天并于下雨前将尿素溶于10 L 的水中,用喷雾器在试验小区中来回均匀喷洒,对照组样地喷施同体积的水,防止溶液添加导致的水分效应;施用过磷酸钙时,称好重量后直接均匀撒在试验小区中。
1.3 试验方法与指标测定
1.3.1 植被群落调查 2022 年9 月,在每个小区中随机设置1个1 m×1 m 的样方,进行群落特征调查。对样方内出现的物种随机选取4~6 株,测量其高度;植被盖度采用针刺法,统计样方内每个物种的数量;频度采用样圆法[19]测定;地上生物量测定时分种齐地刈割,并将剪下的植物装入纸袋,65 ℃ 烘至恒重获得地上生物量。
1.3.2 植株采样 根据植被群落调查结果,选取各个样方内重要值大于0.1 的物种作为优势物种。随机选取3~5 株成熟、发育良好、株丛大小相似的植株,装入密封袋后立即置于有冰袋的样品箱(<5 ℃)带回室内。
1.3.3 植物功能性状测定 从采集到的各植株上随机选取5 片健康叶片,首先去除叶柄用精度为0.01 mm 的电子游标卡尺(美耐特,MNT-150T,德国)测定叶片厚度(leaf thickness, LT),测定时避开主脉位置,分别测定叶片上、中、下位置并取其平均值;然后将叶片放置在叶面积扫描仪(AM350,英国)上测定叶面积(leaf area, LA);将扫描后的叶片放入装有水的密封袋中,在5 ℃冰箱中静置12 h 后取出,用滤纸迅速吸干叶片表面水分并称量得到叶片饱和鲜重(leaf saturated fresh weight, LSFW);再将称重完的叶片置于65 ℃烘箱中烘48 h 至恒重,称得其干重(leaf dry weight, LDW);最后将植株剩余的叶片与测定完的叶片混合在105 ℃烘箱中杀青30 min,然后置于65 ℃烘箱中烘至恒重,用球磨仪(格瑞德曼,GT600,德国)磨细过0.15 mm 筛,干燥保存,进行叶片碳(leaf carbon, LC)、氮(leaf nitrogen, LN)、磷(leaf phosphorus, LP)含量的测定。全碳使用总有机碳分析仪(VARIO TOC,德国)测定;叶片氮和磷测定时,首先对样品进行H2SO4-H2O2消煮,采用全自动凯氏定氮仪(K-360,BUCHI Labortechnik AG, 瑞士)法测定叶片N 含量,全P 采用钼锑抗比色法测定[17]。
1.4 数据分析
根据植被群落调查及功能性状测定数据,计算以下指标:
1)物种重要值[20]
2)叶片功能性状计算[21]:
3)群落功能性状加权平均值[22]
群落加权平均值(community weighted mean, CWM)代表植物群落中某项功能性状的平均值。本研究中群落功能性状特征值采用群落加权平均值进行分析,计算公式为:
式中:Pi为群落内物种i的相对多度,traiti为物种i的性状值。
4)可塑性指数计算[23]
采用表型可塑性指数(phenotypic plasticity index,PI)来表达各功能性状的可塑性对N、P 添加的响应程度,计算公式如下:
式中:MAX为试验样地群落叶片某一性状的最大值,MIN为群落叶片某一性状的最小值。
5)功能性状变异系数计算[23]
采用变异系数(coefficient of variation,CV)表示植物群落叶片各功能性状在不同N、P 处理下的变异程度,计算公式如下:
式中:SD为植物群落叶片各性状的标准偏差,Naverage为植物群落性状平均值。
使用Excel 2017 对数据进行录入和整理,使用SPSS 26.0 进行数据分析,数据均采用平均值±标准误表示。采用单因素方差分析(One-way ANOVA)研究不同处理水平间的差异;对植物群落叶片性状进行Spearman 相关性分析,该情况下不区分处理;绘图在GraphPad Prism 9.2.0 中完成。
2 结果与分析
2.1 植物群落叶片功能性状对N、P 添加的响应
2.1.1 形态性状对N、P 添加的响应 N、P 添加对植物群落叶片形态性状均有显著影响(图1)。N 单独添加提升了植物群落叶片形态性状,其中,显著增加了植物群落叶面积、比叶面积和叶组织密度(P<0.05),较对照组分别提升了247.33%、106.43%和141.77%;植物群落叶片形态性状在P 单独添加下,与对照组相比呈降低趋势,而在N+P 共同添加下有上升趋势,但均不显著(P>0.05)。

图1 N、P 添加对植物群落叶片形态性状的影响Fig.1 Effects of N and P addition on leaf morphological traits of plant community
各个处理对植物群落叶片形态性状的影响具有显著差异。N 单独添加对植物群落叶面积的影响显著高于P单独添加(P<0.01)和N+P 共同添加(P<0.05);植物群落叶片比叶面积对N 单独添加的响应极显著大于P 单独添加(P<0.01),N+P 共同添加对比叶面积的影响也大于P 单独添加(P<0.05);N 单独添加对干物质含量和叶组织密度的作用显著强于P 单独添加(P<0.05)。
2.1.2 化学性状对N、P 添加的响应 对于植物群落叶片化学性状(图2),N、P 添加对植物群落叶片磷含量、碳氮比和氮磷比无显著影响(P>0.05);N 添加提升了叶片碳含量,P 添加降低了叶片碳含量,但差异均不显著;N单独添加显著提升了植物群落叶片氮含量(P<0.05),且影响程度显著强于N+P 共同添加处理;N+P 共同添加显著提升了植物群落叶片碳磷比(P<0.05),作用强度显著高于P 单独添加处理(P<0.05)。

图2 N、P 添加对植物群落化学性状的影响Fig.2 Effects of N and P addition on chemical traits of plant community
2.2 N、P 添加下植物群落功能性状可塑性特征及变异性
通过对N、P 添加下10个叶片性状进行分析和排序(图3),结果显示,不同叶片性状指标可塑性指数大小排序为:叶面积>比叶面积>叶组织密度>叶片碳含量>叶干物质含量>叶片氮含量>叶片磷含量>叶碳磷比>叶氮磷比>叶碳氮比。叶片形态性状和化学性状的可塑性指数分别为0.55~0.77 和0.14~0.58,其中可塑性较大的指标是叶面积、比叶面积、叶组织密度、叶片碳含量、叶干物质含量和叶片氮含量,可 塑 性 指 数 分 别 为0.77、0.76、0.61、0.58、0.55 和0.51,为敏感性状;叶片磷含量、叶碳磷比、叶氮磷比和叶碳氮比可塑性指数较小,分别为0.47、0.36、0.26和0.14,为惰性性状。
由图3 可知,10 种叶片性状的变异系数具有差异,且对变异系数大小排序后结果与可塑性指数排序一致。叶片形态性状变异系数为0.39~0.64,其中变异性最大的为叶面积,其次是比叶面积、叶组织密度和叶干物质含量;叶片化学性状变异系数为0.08~0.41,其中叶碳氮比变异性最小,相对稳定,叶片碳含量变异性最大。总的来说,可塑性指数较大的性状一般也具有较大的变异性,即叶片敏感性状其变异性较大。
2.3 N、P 添加下植物群落叶片功能性状间的相关性
由表1 可知,植物群落叶面积与比叶面积、叶组织密度、叶干物质含量、叶片碳和磷含量均呈显著或极显著正相关关系(P<0.05;P<0.01),植物群落比叶面积与叶干物质含量、叶片碳和磷含量呈显著正相关关系(P<0.05);植物群落叶组织密度、叶干物质含量和叶片碳含量均与叶片氮、磷含量呈显著或极显著正相关关系(P<0.05;P<0.01);植物群落叶碳磷比与氮磷比之间存在极显著正相关关系(P<0.01)。
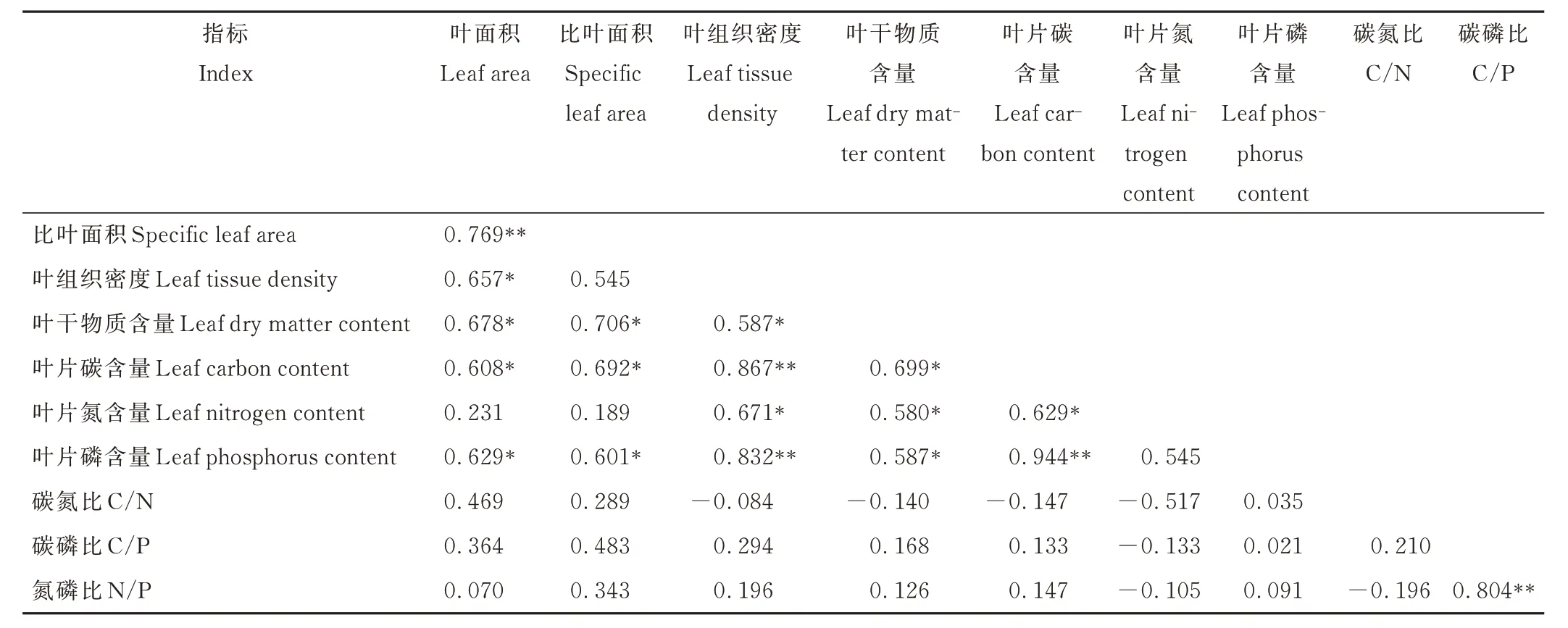
表1 N、P 添加下叶片功能性状间的相关性Table 1 Correlation among leaf functional traits under N and P addition
3 讨论
3.1 N、P 添加对植物群落叶片功能性状的影响
植物通过调节自身形态、化学性状来应对环境变化对其产生的影响。功能性状的群落加权平均值在一定程度上反映了群落内物种的“平均”行为(central behavior),也被认为是获得群落水平性状潜在的重要途径,有研究表明,植物群落加权平均功能性状能够对环境变化做出响应,并影响生态系统过程[24]。本试验中,N、P 添加对植物群落叶片形态性状与化学性状产生了显著的影响。
植物功能性状中,叶面积、比叶面积、叶干物质含量和叶组织密度可以反映植物获取资源的能力[25]。有研究发现,N 添加显著增加了叶片的总面积,显著提高了植物叶片光合能力、气孔导度和蒸腾速率[26]。本研究中,植物群落叶面积与比叶面积在N 单独添加下均得到显著提高,可能随着土壤养分有效性的提高,植物可获取的养分也随之增多,植物比叶面积得以增大增加了其光合能力,采取较快的养分循环策略适应环境[27],这与李颖等[28]的研究结果相似。在本研究中,P 单独添加与N+P 共同添加对植物群落叶面积与比叶面积无显著影响,原因可能是试验样地所处环境受到了N 限制。
干物质含量是叶片的关键性状,反映着植物抵御外界胁迫及资源利用的能力[29]。叶片干物质含量越大,植物叶片组织密度越大,其抵抗力和耐受力越强。本研究中,除了N 单独添加对植物群落叶组织密度有显著影响,其他处理对干物质含量与叶组织密度均无显著影响,与张宇平[30]的研究结果相似。但有研究表明,在施N 处理下,植株高度和比叶面积显著增大,而叶干物质含量显著降低,这反映了植物为提高对资源的利用能力而采取的生存策略[31]。本研究结果与其产生差异的原因可能是养分添加量的不同以及添加方式的差异,还有可能由于研究区环境气候的差异,致使植物对养分添加的响应不同。
叶片的养分浓度反映植物对土壤环境养分吸收的能力。研究表明,外源N 的输入提高了土壤N 矿化速率和N 的有效性[32],从而促进了植物对N 的吸收,使叶片的氮含量增加;而P 的输入也会导致植物叶片中磷含量的增加[33]。本研究结果显示,N 单独添加下,植物群落叶片氮含量显著提升;P 单独添加下叶片氮含量上升,但不显著,N+P 共同添加下叶片氮含量无显著变化。植物群落叶片氮含量对单独P 或N+P 共同添加不敏感的原因可能是植物在养分吸收过程中还受到水分等其他因素影响,养分吸收利用能力降低。Neff 等[34]研究发现,N 添加会增加植物叶片碳含量,本研究结果与之相似,植物群落叶片碳含量在N 单独添加下呈上升趋势,原因可能是N 添加加快了土壤轻组分有机碳的分解速率[35]。植物群落磷含量在N、P 添加下相对稳定,无显著变化。但有研究表明,植物叶片磷含量的变化可能比氮含量的变化更好地反映环境中P 的供应[36],并且单独P 或N+P 共同添加显著提升了植物叶片磷含量[30]。导致结果相反的原因可能是本研究中P 添加的水平较低,为8 g·m-2·a-1,可能更高浓度的P 输入会对植物叶片磷含量的影响更显著。
植物的碳氮比与碳磷比均能反映植物在生长过程中对碳、氮的同化和固定能力[37],其大小与植物的营养利用效率密切相关,当碳氮比较小时,植物分解速率较快,需吸收较大量的N 和P 元素[38]。本研究结果显示,植物群落叶片碳氮比在P 单独添加时略低于CK,而单独N 和N+P 共同添加时,略高于CK,说明N 的输入,提高了植物群落里受N 限制物种的N 吸收率[39]。植物群落叶片碳磷比在N 单独添加下略微高于CK,在N+P 共同添加下显著提升,说明N 添加提高了植物对P 的利用率,导致碳磷比上升。植物叶片氮磷比可以判断其生长的养分受限情况。研究表明,当植物叶片的氮磷比<14 时,受到N 限制,氮磷比>16 时,则受到P 限制[39]。本研究结果显示,各处理下植物群落叶片氮磷比都低于14,表明研究区所处地域植物生长受到N 限制,N、P 添加下叶片氮磷比有上升趋势,但不显著。
3.2 N、P 添加下植物群落叶片性状可塑性特征及变异程度
叶片作为植物主要的营养器官,其功能性状具有较强的可塑性与变异性。可塑性指数与变异系数高的性状特征对环境有着较强的潜在适应能力。表型可塑性是植物用于优化资源获取的重要机制[40],表型可塑性越高,越有利于植物抵御环境变化,可塑性指数能够直观地表达植物形态、生理生态性状对资源变异的响应程度。一般来说,敏感性状受环境变量的影响更大,可塑性倾向于在短时间内使植物获取资源最大化[41]。变异系数可用于表示植物性状的变异程度,通常可塑性较大的功能性状指标其变异性也较大。本研究通过对植物群落叶片可塑性指数与变异系数进行分析排序,发现植物群落叶片功能性状可塑性指数大小排序结果与变异系数相同;叶片形态性状与叶片碳含量的可塑性和变异性较大,说明植物群落在应对由N、P 添加导致的环境变化过程中,形态性状起到主导作用。通常敏感性状受环境变量的影响更大,可塑性倾向于在短期内最大化资源的获取和增长速度[42]。植物群落叶片碳氮比、碳磷比和氮磷比可塑性指数与变异系数较低,其原因可能是在荒漠草原中植物群落绝对优势种能够使植物体内的化学计量比保持一定程度的稳定[42],并且这些惰性性状对充分发挥其在干旱环境的生产功能具有重要作用。研究表明,羊草(Leymus chinensis)在长期和短期N 添加下,都是随着N 添加量的增加而由N限制逐渐转为P 限制[43]。本研究试验样地已经进行了3 年的养分添加处理,但N、P 添加量较小,且较为单一,并未缓解植物生长受到的N 限制,导致植物群落叶片化学计量比在N、P 添加下无显著变化。
3.3 植物群落叶片性状间的相关性
植物叶功能性状之间的关系能够表示植物对其生境资源的不同利用方式[44],比叶面积和叶干物质含量是能够综合反映植物生境适应性的关键性状[45]。大量研究表明比叶面积与叶干物质含量之间呈负相关关系[46-47],资源丰富的环境中植物比叶面积更大,其对资源的利用、光合和生长能力更强,而在贫瘠的环境中比叶面积小的植物适应能力更强[48-49]。Rose 等[50]认为具有低比叶面积和高叶干物质含量的植物,其资源获取能力以及水分的利用效率往往更高, 更容易适应恶劣的环境条件。本研究结果显示植物群落叶片比叶面积与叶干物质含量呈显著正相关,同样Qi 等[51]研究发现,在海拔梯度上辽东烁(Quercus wutaishansea)叶片比叶面积和叶干物质含量有着密切的正相关关系,都间接证明了叶片比叶面积和叶干物质含量在叶片中有相同的促进植物吸收养分资源的功能。然而,大多数研究表明比叶面积与叶干物质含量呈负相关关系[52],本研究结果与其不一致,可能是受植物生长地域及各个物种自身的生理特征和生长策略的影响,使植物对N、P 添加的响应不同。本研究中,植物群落叶干物质含量与叶组织密度呈显著正相关关系,且二者均与叶片碳、氮、磷含量呈显著正相关关系,表明植物叶片在N、P 添加下叶组织密度的增加导致单位面积干物质含量的增加,这与许多研究结果相似[53],并且说明叶片碳、氮、磷含量是影响叶组织密度和叶干物质含量的关键因子。植物群落叶片比叶面积与叶片碳含量和叶片磷含量呈显著正相关关系,说明比叶面积增大,植物生长加快,光合能力加强,对养分的需求增加。植物群落叶片氮含量与叶片磷含量之间呈正相关关系,可能由于氮磷比是判断植物生长是否受到限制的指标,一个稳定的群落,其限制植物生长的养分元素也是稳定的,因此叶片氮、磷含量呈正相关关系。
4 结论
综上所述,荒漠草原植物群落叶片形态性状与化学性状的变异响应了N、P 添加。结果表明,研究区植物生长受到N 限制, N 添加显著提升了植物群落叶面积、比叶面积、叶组织密度与叶片氮含量,N+P 共同添加下植物群落叶氮磷比有所增加,但不显著,预测更高浓度的N、P 添加会缓解这种限制;群落叶片形态性状为响应N、P 添加的敏感性状,表明植物群落主要通过改变自身形态来应对环境的变化;植物群落叶片性状之间存在显著的相关性,并且表现出应对养分环境变化的协同作用。
因此,大气N、P 沉降加剧可能会导致植物群落性状的变异从而对荒漠草原生态系统功能产生影响,该影响还有待研究。此外,后续研究需要考虑环境因素对植物功能性状的影响,例如降水、光照等,还要考虑更多维度功能性状对不同养分添加的响应,从而更加全面地了解荒漠草原植物群落对养分添加的适应策略。
