白菜种子cDNA 酵母文库的构建及BrTTG1 互作蛋白的筛选及分析
2024-03-12任延靖张鲁刚赵孟良李江邵登魁
任延靖 张鲁刚 赵孟良 李江 邵登魁
(1.青海大学农林科学院 青藏高原种质资源研究与利用实验室,西宁 810016;2.农业农村部青藏高原种质资源保护与遗传改良重点实验室,西宁 810016;3.西北农林科技大学园艺学院,杨凌 712100)
白菜(Brassica rapa.L.)是我国栽培面积最广,也是最常见的蔬菜作物之一,同时也是高海拔地区主要的油料作物,具有耐贫瘠、耐寒等优点。种皮颜色是十字花科芸薹属作物一个重要的园艺性状[1-2],根据种皮颜色的不同可以将芸薹属作物的种子分为黄色种子和棕色种子两大类,研究表明黄籽具有极耐贮存的特点[3],同时黄籽作为优质的榨油材料,具有含油量高、蛋白含量高等优点,在油料开发方面具有极大的潜力[4]。因此,对种皮颜色形成关键物质——原花青素的研究,对于白菜品质育种具有重要的意义。
在对种皮颜色的研究中发现,芸薹属作物种皮颜色的多样性,主要是由于原花青素含量的差异积累造成的,黄色种皮的出现是由于缺少原花青素的积累,使得种皮颜色呈现透明色,最终表现出种胚的颜色[5]。研究还发现,TTG1 的突变能够影响种皮色泽的形成[6]。在白菜中,Zhang 等[1]利用有光泽、黄籽的白菜和有刺毛、黑籽的白菜构建了一个DH 群体,用以研究白菜种皮颜色、刺毛这两个性状,结果显示TTG1 基因同时控制着种皮颜色和表皮毛性状,并且通过异源表达分析发现,TTG1 可以有效的恢复拟南芥种皮变黄的性状。
TRANSPARENT TESTA GLABRA 1(TTG1)是一种WDR(WD repeat)蛋白,研究表明WDR 蛋白家族具有广泛的生物化学和细胞生物学功能,如参与细胞分裂和胞质移动、细胞程序性死亡、光信号感受和传导、细胞运动、花发育、开花等过程[7]。TTG1 作为WDR 家族成员在大多数植物的系统发育过程中起着重要的作用[8-9],包括叶毛、茎、根的形成,类黄酮和花青素的生物合成,种皮和种皮色素的发育[6]等。在种皮颜色形成中,MYB 和bHLH 蛋白与TTG1 能够结合形成三元蛋白复合物,称为MBW复合物(MBW complex)。三元复合体调控植物类黄酮代谢相关结构基因的表达,从而催化花青素和原花青素的合成、修饰和转运,使其在特定组织部位积累,是植物种皮颜色形成的关键调控复合体[10-12]。
本研究前期采用正向遗传学的方法鉴定出TTG1为调控种皮颜色形成的候选基因,TTG1 的缺失突变导致了黄籽的产生[2-3],但是在白菜中关于种皮原色形成的三元复合体尚未见报道。基于此,本研究以前期获得的棕籽白菜自交系‘B147’为研究对象,构建白菜种子酵母双杂交文库并筛选TTG1 互作蛋白,以期为阐释白菜种皮颜色形成的调控网络奠定良好的基础,同时为其他作物种皮颜色形成机理的研究提供理论指导。
1 材料与方法
1.1 材料
选用棕籽白菜高代自交系‘B147’作为试验材料,于2017 年3 月定植于西北农林科技大学园艺学院试验基地,待抽薹开花后,选择花期进行自交授粉,获得不同发育时期的新鲜种子(花期自交授粉后10、14、18、22、26、30、34 d),选择原花青素开始积累后的不同发育时期的种子,包括14、18 和22 DAF 的种子,进行cDNA 文库的构建。
1.2 方法
1.2.1 cDNA 文库构建 参考TRIzol Reagent 试剂盒(Invirtrogen,USA)说明书分别进行总RNA 的提取,根据Oligotex mRNA Midi Kit 说明书(QIAGEN,德国)进行mRNA 分离纯化,并进行质量评估及纯度检测,用于后续的文库构建。
参考CloneMinerTMII cDNA Library Construction Kit(Invirtrogen,USA)试剂盒说明书合成双链cDNA 并与attB1 重组接头连接,将cDNA 分级分离并回收500-4 000 bp 的双链cDNA,采用BP 反应将双链cDNA 与pDONR222 载体连接并转入大肠杆菌(Escherichia coli)DH10B 感受态细胞中,获得初级文库菌液。取菌液10 μL 稀释100 倍后,取50 μL涂布于含有卡那霉素(Kan)的LB 平板进行库容量的鉴定。随机挑选24 个克隆进行菌落PCR,鉴定文库的重组率和插入片段长度。文库滴度(CFU/mL)=平板上的克隆数/平板上涂布菌液的体积(μL)×稀释倍数(100)× 103μL;文库总库容(CFU)=文库滴度(CFU/mL)×文库菌液总体积(mL)。
从上述鉴定合格的初级文库中抽提质粒并稀释至300 ng/μL,通过LR 重组反应与pGADT7-DEST进行连接,并转入大肠杆菌DH10B 感受态细胞中,获得次级文库菌液,并按照上述相同的方法进行库容量、重组率和插入片段长度的鉴定。
1.2.2 诱饵载体pGBKT7-TTG1 的构建与自激活性鉴定 根据TTG1 基因的序列[3]和pGBKT7 载体序列,设计TTG1 诱饵克隆的引物,引物上下游分别包含EcoR I 和BamH I 的酶切位点(上游引物为5'-CCGAATTCATGGACAACTCAGCTCCGGACTCC-3',下游引物为5'-ACGGATCCTCAAACTCTAAGGAGCTG CATTTT-3'),以棕籽白菜‘B147’花期自交授粉后第14 天的种子总RNA 反转录获得的cDNA 第一链为模板,扩增获得带有EcoR I 和BamH I 酶切位点的TTG1 基因序列。采用EcoR I 和BamH I 双酶切PCR 获得的TTG1 基因产物,获得含有酶切位点黏性末端的TTG1 基因序列并纯化,同时采用EcoR I和BamH I 双酶切pGBKT7 质粒并纯化,同样获得含有酶切位点黏性末端的线性化pGBKT7 质粒载体,最后采用DNA 聚合酶将TTG1 基因融合到线性化的pGBKT7 质粒载体上,获得含有TTG1 基因的重组pGBKT7-TTG1 质粒,参照Matchmaker Gold Yeast Two-Hybrid System(Clontech,USA)操作手册,将构建好的pGBKT7-TTG1 载体质粒以LiAc 法转化Y2H Gold 菌株,并将重组质粒整合到Y2H Gold 酵母菌株(Clontech,USA)基因组中,获得酵母双杂交TTG1-诱饵菌株。
以pGBKT7-Lam/pGADT7-T 为阴性对照、pGBKT7-53/pGADT7-T 为阳性对照、pGBKT7-TTG1/p-GADT7 为自激活检测组,分别共转至Y2H Gold 酵母菌株中,采用质量体积分数为0.9%的NaCl 溶液重新悬浮细胞并稀释后,分别取50 μL 涂布于SD/-Trp/-leu/X-α-gal(阴性对照、阳性对照和自激活检测组)、SD/-Trp 和SD/-Trp/X-α-gal/AbA(自激活检测组)培养基上,30℃倒置培养3-5 d 后,观察菌落生长情况。
1.2.3 酵母双杂交筛选TTG1 互作蛋白并验证 取5 μL pGBKT7-TTG1 质粒与10 μg 酵母文库质粒共转化后,涂布于SD/-Trp/-Leu 平板上用以计算转化效率,涂布于SD/-Trp/-Leu/X-α-Gal/AbA 进行初筛,将初筛平板上蓝色阳性克隆转移到二次筛选培养基SD/-Trp/-Leu/-Ade/-His/X-α-Gal/AbA 上进行 高严谨度筛选,30℃培养箱培养3-5 d,挑取平板上蓝色菌落,并提取酵母质粒,参考Matchmaker Gold Yeast Two-Hybrid System 操作手册,分别将Prey 质粒与Bait 质粒共转化Y2H Gold 菌株后涂布于SD/-Trp/-Leu/-His/-Ade/X-α-Gal/AbA 平板上进行验证,验证正确的互作蛋白基因送至上海生工生物科技有限公司进行测序,根据测序获得的编码候选互作蛋白的ORF 序列,参考NCBI 数据库预测各候选互作蛋白的功能。
1.2.4 互作蛋白基因的克隆及生物信息学分析 参考大白菜基因组数据库设计引物,上游序列为5'-AT GTCAGGTCCGTCTAGAAAGAACAT-3',下游引 物为5'-CTACTCCATCTTCCCAATACCGATT-3',以棕籽‘B147’的cDNA 为模板进行MYB73 基因序列的扩增,扩增反应体系如下:PrimeSTAR®Max DNA Polymerase 为10.0 μL,上、下游引物各0.5 μL,模板cDNA(100 ng/μL)为1.0 μL,用ddH2O 补充至20.0 μL。将反应产物使用Universal DNA Purification Kit(TaKaRa,大连,中国)进行纯化,将胶回收产物与pMD18-T 载体连接并将连接产物转入DH5α 感受态中,置于LB 平板上培养。挑单克隆进行菌液PCR 检测,阳性菌液送公司测序。
采用NCBI 数据库进行序列结构的预测分析,测序结果通过DNAMAN 软件进行分析整理,采用在线工具ExPaSy-Protparam 和ExPaSy-Translate[13]预测基因开放阅读框编码的蛋白质理化性质、ExPaSy-ProtScale[13]分析氨基酸的亲疏水性、SOPMA[14]软件分析蛋白质的二级结构,利用MEGA X 软件构建系统发育树。
1.2.5 互作蛋白基因的表达分析 选取棕籽白菜‘B147’和黄籽白菜‘B80’生殖生长过程中,共7 个时期的新鲜种子,采用TaKaRa MiniBEST Plant RNA Extraction Kit(TaKaRa,大连,中国)提取总RNA,并检测提取总RNA 的完整性、浓度及质量。通 过PrimeScriptTMRT reagent Kit with gDNA Eraser(TaKaRa,大连,中国)试剂盒去除基因组DNA 及合成cDNA。
根据获得的基因cDNA 序列,采用Primer Premier 6.0 软件设计实时荧光定量PCR 引物,上游序列为5'-GAAGAACCGACAACGCTATCA-3',下游引物 为5'-CCGTCAGTAACCGTATTCTCATT-3',分析MYB73 基因在棕籽材料‘B147’不同发育时期的种子中的mRNA 积累水平。每个样本3 次生物学重复,3次技术重复,采用2-ΔΔCT法计算基因的相对表达量。
2 结果
2.1 酵母文库的构建及鉴定
选用棕籽白菜高代自交系‘B147’花期自交授粉后14、18 和22 DAF 的种子提取总RNA 后,经过mRNA 分离、纯化、连接转化涂板后,得到cDNA 初级文库,平板计数显示菌群数为3.0×106CFU/mL,克隆总数约为1 500 个,计算库容量约为1.2×107CFU(图1-A),随机挑取24 个克隆进行菌落PCR 鉴定,凝胶电泳结果显示,24 个克隆均具有单一条带,片段平均长度>1 000 bp 且主要集中在1 000-2 000 bp(图1-B),重组率为100%,达到初级文库的要求。
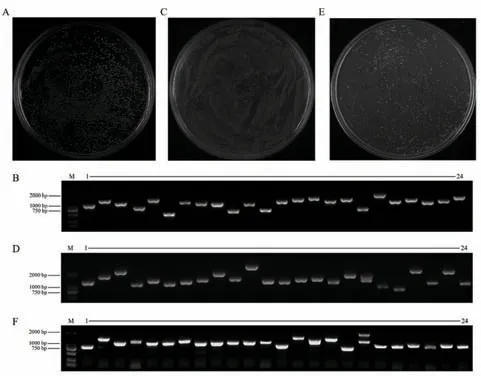
图1 文库的构建及鉴定Fig.1 Construction and identification of library
从上述鉴定合格的初级文库中抽提质粒,与pGADT7-DEST 进行连接转化并涂板,得到次级文库,菌群数为4.0×106CFU/mL,容量约为1.6×107CFU(图1-C),随机挑取24 个克隆进行菌落PCR 鉴定,凝胶电泳显示24 个克隆具有单一条带,片段平均长度>1 000 bp(图1-D),重组率为100%,表明其多态性好。
提取文库质粒转化至Y187 酵母菌株,细胞密度为5.0×107cells/mL(图1-E),随机挑取24 个克隆进行菌落PCR 电泳检测,结果显示插入片段均大于1 000 bp(图1-F)。以上结果显示该文库满足酵母cDNA 文库要求,具备筛库条件。
2.2 诱饵载体pGBKT7-TTG1的构建与自激活性鉴定
根据TTG1 基因全长,扩增获得目的片段1 014 bp,送公司测序后结果对比一致,使用同源重组方法获得含有TTG1 基因的重组pGBKT7-TTG1 质粒。提取重组质粒,分别将阳性对照和阴性对照共转至AH109 酵母感受态细胞中,转化后的酵母细胞涂布于SD/-Leu/-Trp/X-α-gal 固体培养基上倒置培养。将自激活检测组分别涂布于SD/-Leu/-Trp/X-α-gal、SD/-Leu/-Trp/-His/X-α-gal、SD/-Leu/-Trp/-His/-Ade/X-αgal/AbA(200 ng/mL)固体培养基上培养。结果(图2)显 示,pGBKT7-53 和pGADT7-T 于SD/-Leu/-Trp/Xα-gal 培养基中有克隆生长且颜色蓝色(图2-A),pGBKT7-Lam 和pGADT7-T 于SD/-Leu/-Trp/X-α-gal培养基中有克隆生长但不变蓝(图2-B),pGBKT7-TTG1+pGADT7 于SD/-Leu/-Trp/X-α-gal 培养基上生长且变蓝色(图2-C),在SD/-Leu/-Trp/-His/X-α-gal 培养基上生长(图2-D),说明pGBKT7-TTG1 质粒成功转入酵母菌株中,且在SD/-Leu/-Trp/-His/X-α-gal上存在自激活现象;在SD/-Leu/-Trp/-His/-Ade/X-αgal/AbA 培养基上不生长(图2-E),自激活活性受到抑制,说明SD/-Leu/-Trp/-His/-Ade/X-α-gal/AbA 筛选条件可以抑制pGBKT7-TTG1 的自激活,进行后续的筛库实验。
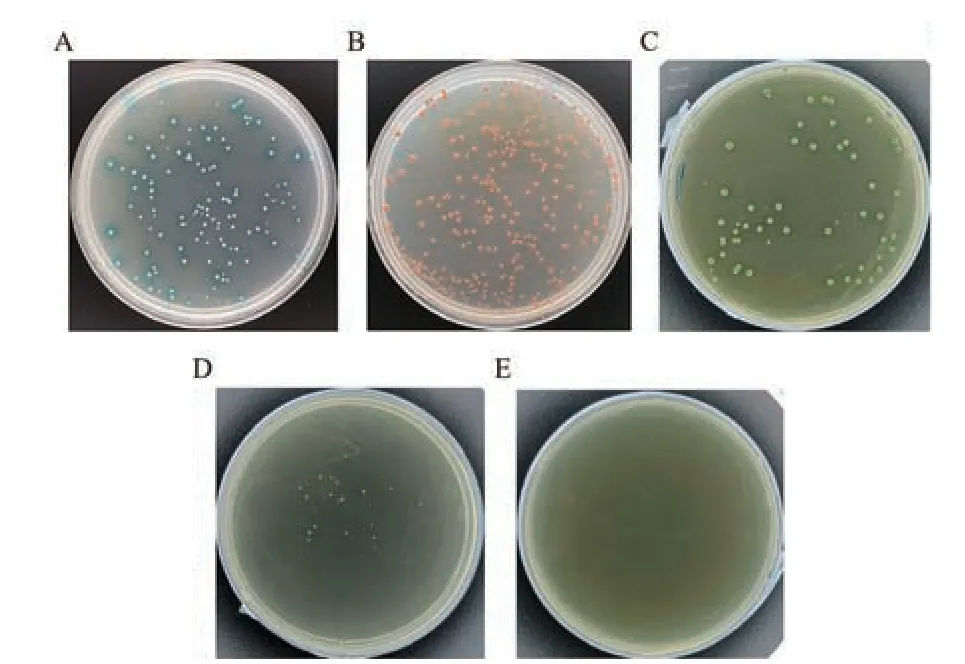
图2 pGBKT7-TTG1 诱饵载体的自激活鉴定Fig.2 Self-activation identification of pGBKT7-TTG1 bait vector
2.3 BrTTG1互作蛋白筛选
将pGBKT7-TTG1 质粒与文库质粒共转至AH109 酵母感受态细胞,于SD/-Leu/-Trp/-His/X-αgal(TDO/X)筛选平板上进行初次筛选,共有120个蓝色克隆。转化效率>2×105CFU/μg,总转化子数>2×106CFU,挑取TDO/X筛选平板上的蓝色克隆,转到SD/-Leu/-Trp/-His/-Ade/X-α-gal/AbA(QDO/X/A)筛选平板上进行二次筛选,共有82 个克隆生长蓝色(图3),随后挑取上述82 个蓝色克隆进行摇菌,提取酵母质粒,转化大肠杆菌,抽提AD 质粒后再与pGBKT7-TTG1 质粒进行一对一互作回转验证并测序,测序结果显示,蓝色克隆50、51、52 和53 的ORF 序列信息相同,预测为同一个基因,获得44个阳性互作基因。对44 个阳性克隆测序结果进一步分析,结果显示蓝色克隆11、18、21、32、58 以及72 的序列信息不是完整的开放阅读框,无法进行后续的试验,故排除这6 个阳性克隆,最终得到38 个测序正确阳性互作基因(图4,表1)。
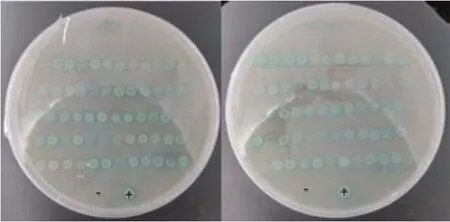
图3 BrTTG1 在SD/-Leu/-Trp/-His/-Ade/X-α-gal/AbA(QDO/X/A)平板筛选的蓝色克隆Fig.3 Blue colonies screened by BrTTG1 in SD/-Leu/-Trp/-His/-Ade/X-α-gal/AbA(QDO/X/A)medium

图4 BrTTG1 候选互作蛋白的一对一互验证Fig.4 One-to-one verification of BrTTG1 candidate interaction protein
综合分析这38 个基因,有一个阳性互作基因注释为MYB44 基因,根据其测序的结果及其在BRAD(http://brassicadb.org/brad/)数据库中比对结果联合分析,确定该候选基因为大白菜MYB73 基因,推测该基因可能为参与MBW 三元复合体的候选基因。
2.4 MYB73基因的克隆及生物信息学分析
测序结果(图5)显示,白菜MYB73 基因CDS全长为987 bp,编码328 个氨基酸的肽链,利用SMART 软件进行保守结构域预测及分析,结果(图6)显示,MYB73 含有2 个典型的SANT-MYB 结构域和MYB 型抑制子的保守基序C1-motif 和C2-motif,推测MYB73 属于R2R3-MYB 类型抑制子,理化性质分析显示,该蛋白相对分子质量35.36 kD,蛋白质等电点为6.27,不稳定指数为63.37,脂肪指数58.20,该蛋白富含Gly(13.7%)、Ser(10.7%)、Glu(8.8%)、Arg(7.0%),其中带负电荷的氨基酸残基(Asp,Glu)数为39 个,带正电荷的氨基酸残基(Arg,Lys)数为37 个。二级结构分析显示该蛋白中无规则卷曲占比最大,为62.5%;α-螺旋次之,占比26.83%,延伸链占比为7.01%,而β-转角所占比例最小为3.66%。蛋白质的碳端和氮端大多为α-螺旋和无规则卷曲结构,亲水性分析显示该蛋白为亲水性蛋白。
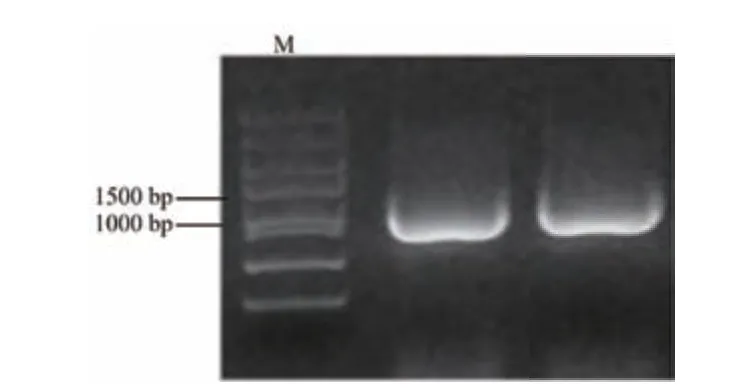
图5 MYB73 基因PCR 扩增电泳图Fig.5 Electrophoretic map of MYB73 gene PCR amplification

图6 MYB73 氨基酸序列与其他MYB 型抑制子序列比对图Fig.6 Sequences alignment of MYB73 amino acid with other MYB type suppressors
选取已知的参与MBW 三元复合体的MYB 转录因子构建系统发育树,包括R2R3-MYB 型激活子AtTT2(AED93980.1)、AtMYB5(NM_112200.3)、AaMYB3(MH_349476)、DkMYB2(AB_503699)、DkMYB4(AB_503701)、FaMYB1(AF_401220.1)、FaMYB9(OK_001453)、HtMYB2(MN887536.1)、MdMYB11(NM_001294029.1)、MdMYB12(HM122616)、MtMYB5(AES71860.2)、MtMYB14(JN157821)、TaMYB14(JN052773)和VvMYBPA1(NM_001281231),R2R3-MYB 型抑制 子AtMYB4(AB005889.1)、AtMYB6(NM_117014.3)、AtMYB7(NM_127224.6)和FaMYB1(AF_401220.1)。利用MEGA X 软件,采用最大似然法重复1 000 次进行MYB 转录因子的亲缘关系分析,结果(图7)显示,所有的MYB 转录因子被聚为5 类,其中所有的MYB 激活子被聚为一类,AtMYB6 和AtMYB7 被聚为一类,MYB73、FaMYB1 和HtMYB2 分别单独聚为一类。
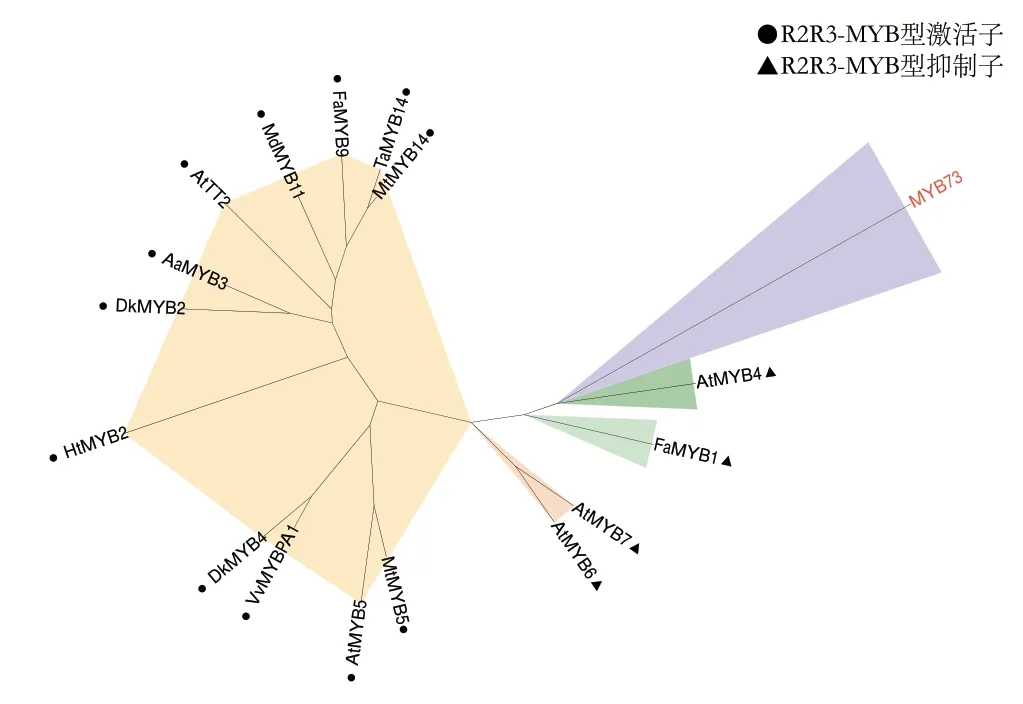
图7 MYB 转录因子的系统发育关系分析Fig.7 Phylogenetic relationships of MYB transcription factors
2.5 MYB73在棕籽和黄籽不同发育时期的表达特征分析
通过实时荧光定量PCR 分析MYB73 基因在棕籽白菜‘B147’和黄籽白菜‘B80’的7 个不同发育时期种子中的表达情况进行分析,结果(图8)显示,MYB73 基因在‘B147’的表达量始终高于在‘B80’中的表达量,且在14 d 时的表达量最高,该结果与TTG1 基因在‘B147’与‘B80’中的相对表达量趋势一致,暗示着MYB73 可能参与了白菜种皮原花青素形成,从而影响种皮的颜色。
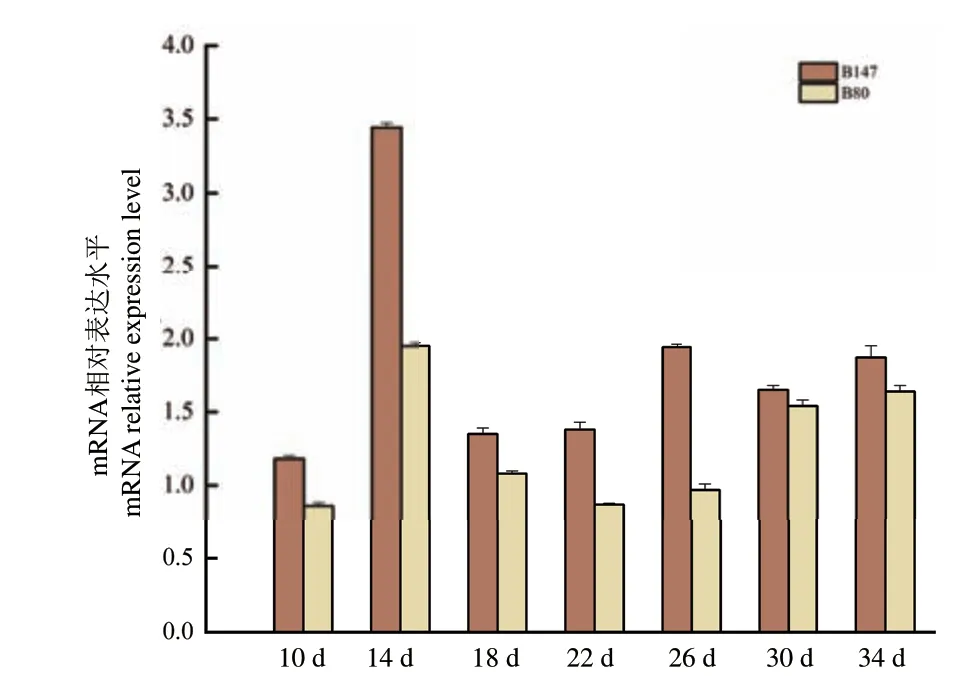
图8 MYB73 基因在‘B147’与‘B80’不同时期发育种子中的相对表达量Fig.8 Relative expression of MYB73 in seeds at different development stages in ‘B147’ and ‘B80’
3 讨论
随着功能基因组学的发展,利用酵母双杂交技术研究蛋白之间的相互作用已成为分子生物学领域的研究热点之一[15]。TRANSPARENT TESTA GLABRA 1(TTG1)作为WDR(WD repeat)蛋白,具有广泛的生物化学和细胞生物学功能,Imamura等[16]认为TTG1 是一种参与毛状体形成的WD40 蛋白,WD40 结构域的功能是能够提高与其他蛋白的相互作用,具有广泛的生物化学和细胞生物学功能,如参与细胞分裂和胞质移动、细胞程序性死亡、光信号感受和传导、细胞运动、花发育、开花等过程[7]。
本研究前期采用正向遗传学的方法,鉴定出TTG1 为调控种皮颜色形成的候选基因,TTG1 的缺失突变导致了黄籽的产生[2-3],但是在白菜中关于种皮原色形成的三元复合体尚未见报道,因此本研究首次开展了白菜种子组织酵母文库的构建,并筛选获得与TTG1 相互作用的一个MYB 抑制子MYB73,在其他作物种皮色素的研究中,MYB和bHLH 蛋白与TTG1 可以形成MBW 复合物,调控植物类黄酮生物合成相关结构基因的表达,从而催化花青素和原花青素的合成、修饰和转运,使其在特定组织部位积累,是植物种皮颜色形成的关键调控复合体[10-12]。在拟南芥(Arabidopsis thaliaba)中,目前已经报道了4 种MBW 复合物,包括TT2-TT8-TTG1、MYB5-TT8-TTG1、TT2-EGL3-TTG1 和TT2-GL3-TTG1,这些复合物在花青素积累中具有重要的调节作用[11],此外,Liu 等[17]在对蒺藜苜蓿(Medicago truncatula Gaertn)种皮颜色的研究中发现,MtMYB5 和MtMYB14 能够和bHLH、TTG1 形成单独的MYB5/MYB14-TT8-TTG1 三元复合体,两者之间还可以相互作用,最终形成MYB5-MYB14-TT8-TTG1 四元复合体调控蒺藜苜蓿种皮原花青素的积累,经鉴定MtMYB5 和MtMYB14 均为激活型的R2R3-MYB。Zhu 等[18]通过对彩叶草(Solenostemon scutellarioides)SsMYB3 在拟南芥中的异位表达及酵母双杂交试验发现,彩叶草SsMYB3 可以与AtTT8 与AtTTG1 相互作用,以改变其三元转录复合物。在转基因烟草(Nicotiana tabacum)中,还发 现SsMYB3 能够与烟草bHLH 蛋 白(NtAn1a 和NtJAF13-1)和一个WD40 蛋白NtAn11-1 相互作用增加浓缩单宁的积累。Zhao 等[19]发现烟草R2R3-MYB 因子NtMYB330 被表征为PA 特异性调节剂,其中PA 生物合成在NtMYB30 过表达系的花中被促进,而在NtMYB330 突变体的花中减少,NtMYB330可以与类黄酮相关bHLH 伴侣NtAn1b 和WDR 蛋白NtAn11-1 相互作用,并且需要NtMYB330-NtAn1b复合物来实现PA 相关结构基因NtDFR1、NtANS1、NtLAR1 和NtANR1 的强转录激活。Zhou 等[20]在桃子(Prunus persica)的研究中报道了桃子中的一个R2R3-MYB 抑制子PpMYB18,它可作为PA 积累的负调节剂;随后Rajput 等[21]在香蕉(Musa nana L.)的研究中发现了R2R3-MYB 激活子和R2R3-MYB 抑制子可以相互作用调节原花青素的生物合成。Qu等[22]在对油菜(B.napus)黄籽的研究中发现,编码R2R3-MYB 型转录因子的BnA09MYB47a 直接激活TT18 来促进原花青素的积累。本研究中以TTG1作为诱饵蛋白,筛选出了白菜中与TTG1 相互作用的一个MYB 抑制子,与已知的参与MBW 三元复合体的MYB 蛋白均不同,可能是尚未被报道的能够与TTG1 互作的R2R3-MYB 型抑制子,也是白菜种皮颜色中首次报道的R2R3-MYB 型抑制子。
综合分析目前对于原花青素的研究,已经报道了众多能够和bHLH、TTG1 相互作用促进或抑制原花青素合成的MYB 转录因子,且报道的基因各不相同,说明在不同作物中,原花青素的调控可能有不同的MYB 转录因子参与。本研究为种皮原花青素的形成提供了新的MYB 转录因子,有助于完善种皮原花青素调控网络的研究。
4 结论
本研究发现了一个能够与TTG1 相互作用的MYB 蛋白,注释为MYB73 基因,结合序列保守基序和系统发育树分析结果,说明MYB73 与已经报道的参与MBW 三元复合体的MYB 蛋白均不同,可能是尚未被报道的能够与TTG1 互作的R2R3-MYB型抑制子,这也是首次在白菜中发现调控原花青素形成的MYB 抑制子,暗示着白菜中可能存在不同MYB 转录因子参与的调控网络影响着原花青素的形成,为后期探究白菜种皮原花青素的调控网络奠定良好的基础。
