番茄HMA 基因家族的鉴定及SlHMA1 镉转运功能研究
2024-03-12赵曜文朗骆少丹栗子杏刘超超
赵曜 文朗 骆少丹 栗子杏 刘超超
(江苏科技大学生物技术学院,镇江 212100)
番茄(Solanum lycopersicum L.)是我国重要的设施蔬菜作物,但在设施生产过程中,人们大量施用含重金属的农药、化肥以及污水灌溉,使得设施土壤和蔬菜的镉等重金属污染问题十分突出[1-3]。因此,研究并鉴定番茄中重金属吸收和转运相关的关键基因对于利用现代分子育种技术培育低镉积累番茄品种具有重要的现实意义。
植物主要通过多种金属转运蛋白吸收并转运环境中的各种金属元素[4-6]。已经报道的与重金属运输有关的蛋白主要有ZIP 家族(Zn-regulated transporter and iron-regulated transporter-like)[7]、ABC 家族(ATP-binding cassette transporters)[8]、NRAMP家 族(natural resistance-associated macrophage protein)[9-10]、YSL 家族(yellow stripe-like transporter)[11]、MTP 家族(metal tolerance protein)[12]和P 型ATP 酶家族(P-type ATPases)等[13-14]。重金属ATP酶(heavy metal ATPase,HMAs)也被称为P1B型ATP酶,属于P 型ATP 酶家族中的一个亚类,通过水解ATP 提供能量进行金属离子的转运[15]。HMA 家族成员不仅吸收转运植物生长发育所必需的金属元素,如铜(Cu2+)、锌(Zn2+)和钴(Co2+)等,也吸收转运非必需的金属元素,如镉(Cd2+)和铅(Pb2+)等[16]。不同的HMAs 对其底物金属离子的转运具有选择性,基于此,HMAs 被分为2 个主要的亚类,即Zn/Co/Cd/Pb P1B-ATPase 亚类和Cu/Ag P1B-ATPase 亚类[17]。目前,拟南芥(Arabidopsis thaliana)[15]、水稻(Oryza sativa)[18]、苜 蓿(Medicago truncatula)[19]、大 麦(Hordeum vulgar)[20]、芥菜(Brassica juncea)[21]等多个物种的HMA 家族成员已完成了基因组水平的系统鉴定,并且,拟南芥和水稻中分别鉴定到8 个和9 个HMA 家族成员,其中,AtHMA1-AtHMA4 和OsHMA1-OsHMA3 属 于Zn/Co/Cd/Pb P1B-ATPase 亚类,而AtHMA5-AtHMA8 和OsHMA4-OsHMA9 属 于Cu/Ag P1B-ATPase 亚类。研究发现,水稻OsHMA3 定位于液泡膜,能将Cd 隔离到根细胞液泡中,从而限制Cd 从水稻根系向地上部的转运,降低Cd 在籽粒中的积累,同时也显著增强了水稻的Cd 抗性,表明HMA 家族基因在低镉积累作物品种选育方面具有重要的应用潜力[22]。
目前,关于番茄SlHMAs 成员的系统鉴定尚未见报道,SlHMAs 成员的金属转运活性,尤其是镉转运活性及其在镉胁迫响应过程中的功能也尚未得到有效鉴定。
本研究将从全基因组水平鉴定番茄SlHMAs 基因家族成员,并对其理化性质、染色体定位、基因结构、启动子顺式作用元件等特征进行分析。此外,研究其在镉胁迫下的基因表达响应,并结合酵母功能互补试验,研究SlHMA1 在番茄镉吸收过程中的重要功能,为解析番茄重金属转运机制及番茄低镉积累种质创新提供理论依据。
1 材料与方法
1.1 材料
以番茄栽培品种‘Ailsa Craig’作为试验材料。番茄种子依次经75%酒精消毒2 min、10%次氯酸钠消毒15 min 后,用灭菌水清洗4 遍,之后播种于1/2 MS 培养基中垂直放置(培养皿长×宽=15 cm×15 cm),待种子萌发7 d 后,选取长势一致、根长在8 cm 左右的幼苗,移栽于水培箱中,以白色泡沫板和水培棉固定,以1/2 霍格兰营养液进行培养,每5 d 换一次营养液。待番茄苗长至4 叶一心期,以50 μmol/L CdCl2进行处理,设置3 个生物重复,分别在处理0、1、3、5 和7 d 时取根系和叶片样品,液氮速冻,-80℃保存备用。
1.2 方法
1.2.1 SlHMAs 家族成员鉴定及蛋白理化性质分析 从拟南芥TAIR 数据库(www.arabidopsis.org/)和水稻数据库(http://rice.uga.edu/)分别下载拟南芥和水稻的全部HMA 成员的蛋白序列,从番茄数据库网站(http://solgenomics.net/)下载番茄的全基因组蛋白序列,利用TBtools 软件的BLAST 功能,在番茄全基因组蛋白序列中同源比对获得拟南芥和水稻全部HMA 成员的同源蛋白序列,进一步在HMMER网站(http://hmmer.org/)和NCBI Batch CD-Search 工具(https://www.ncbi.nlm.nih.gov/)对上一步得到的番茄HMA 候选蛋白序列进行保守结构域分析,以至少包含E1-E2 ATPase domain(PF00122)、hydrolase domain(PF00702)及HMA domain(PF00403)中2 个以上结构域的蛋白确定为HMA 家族基因。使用ExPASy(https://web.expasy.org/protparam/)在 线工具分析SlHMAs 蛋白的理化性质,包括氨基酸数目、分子量和等电点。在WoLF PSORT 网站(https://wolfpsort.hgc.jp/)对其亚细胞定位进行预测分析。
1.2.2 SlHMAs 家族基因染色体定位、系统进化树构建及共线性分析 使用TBtools 软件GFF3 gene position 工具绘制SlHMAs 家族基因在染色体上的位置;利用MEGA11 软件Align by Clustal W 工具进行蛋白多重序列比对,采用Neighbor-joining 法构建系统发育进化树,Bootstrap 值设定为1 000;使用TBtools 软件One step MCScanX 工具对SlHMAs 家族基因进行共线性分析。
1.2.3 SlHMAs 基因结构、蛋白保守基序及启动子顺式作用元件的分析 利用TBtools 软件,分别从番茄基因组序列文件和基因注释文件中筛选SlHMAs基因家族的DNA 和CDS 序列,利用Gene Structure Display Server(GSDS)(http://gsds.gao-lab.org/)对SlHMAs 家族基因进行基因结构分析。利用MEME Suite 5.5.4(https://meme-suite.org/meme/)对SlHMAs蛋白进行保守基序预测,并利用TBtools 软件对保守基序进行可视化分析。
利 用TBtools 软件提 取SlHMAs 基 因CDS 上游的2 000 bp 序列,通过PlantCare 数据库(http://bioinformatics.psb.ugent.be/webtools/plantcare/html/)预测顺式作用元件。
1.2.4 RT-qPCR 检测镉胁迫下SlHMAs 的基因表达模式 采用总RNA 提取试剂盒(南京诺唯赞,货号R401)提取总RNA,用反转录试剂盒(南京诺唯赞,货号R312)反转录获得总cDNA,使用染料法定量PCR 检测试剂盒(南京诺唯赞,货号Q441)以番茄SlActin(Solyc04g011500.3.1)作为内参基因进行实时荧光定量PCR(ABI Step One Plus,USA),利用2-ΔΔCt法计算基因的相对表达量。所用引物序列见表1。

表1 RT-qPCR 引物序列Table 1 Primers used for RT-qPCR
1.2.5 酵母异源表达及镉含量检测 以正向引物序 列SlHMA1-F:5'-cgacgatgacgataaggtacctATGtct-GAAGCTCTGCGTCTTTCAAC-3'(下划线 为Kpn I限制性酶切位点),和反向引物序列SlHMA1-R:5'-tgctggatatctgcagaattcTTACAAATGGGCTGCCTGAATGG-3'(下划线为EcoR I 限制性酶切位点),扩增SlHMA1 的CDS 目的片段,将其与酵母表达载体pYES2/CT(Invitrogen,USA)同源重组(南京诺唯赞,货号C112),获得重组质粒pYES2-SlHMA1,送上海生工测序验证。以测序正确的pYES2-SlHMA1质粒为模板,以引物SlHMA1-451D-F:5'-CATAGTATTGCATTTGctAAAACTGGTACATTGACA-3' 和SlHMA1-451D-R:5'-TGTCAATGTACCAGTTTTag-CAAATGCAATACTATG-3' 进 行PCR 扩 增,PCR 产物经Dpn I 酶消化去除模板质粒,纯化后进行同源重组反应,最后转化大肠杆菌,测序鉴定得到pYES2-SlHMA1D451A(D:Asp,天冬氨酸;A:Ala,丙氨酸)点突变质粒。
采 用PEG/LiAC 转化法 将pYES2、pYES2-Sl-HMA1 和pYES2-SlHMA1D451A分别转化至酵母突变菌 株Δycf1 中(MATa、ura3Δ0、leu2Δ0、his3Δ1、met15Δ0、YDR135c∷kanMX4 购买自南京瑞源生物技术有限公司),并涂布于SD/-Ura(Glu)固体培养基。经PCR 鉴定阳性克隆后,于SD/-Ura(Glu)液体培养基中振荡培养至OD600=1.0,离心收集细胞,用1×TE 梯度稀释成10-1、10-2、10-3和10-4等4 个不同浓度,分别取5 μL 菌液点样于SD/-Ura(Gal)和SD/-Ura(Gal)+20 μmol/L CdCl2固体培养基上,30℃黑暗培养48-72 h,观察并拍照菌落生长情况。另外,将上述3 组酵母菌加入40 mL SD/-Ura(Gal)+5 μmol/L CdCl2液体培养基中,调整初始OD600=0.05,置于30℃,200 r/min 条件下振荡培养,分别在0、12、24、36、48、60 和72 h 时测定菌液的OD600值,绘制其生长曲线。另设一组,同上述处理,在48 h 时,收集菌体。菌体经过4 次无菌去离子水洗后,在70℃烘干至恒重。然后,将干燥后的菌体称重,加入5 mL 的HNO3,再进行180℃的消解。最后,使用NexlON®5000 ICP-MS(PerkinElmer,USA)测定样品中的Cd 含量。
1.2.6 HMA1 在代表性物种中的进化分析 以SlHMA1 的氨基酸序列为种子序列,在Phytozome 网站(https://phytozome-next.jgi.doe.gov/blast-search)通 过BLAST 比对获得各代表性物种中的同源基因序列,蛋白多重序列比对及保守氨基酸序列分析同1.2.3。
2 结果
2.1 SlHMAs基因家族成员鉴定和理化性质分析
利用拟南芥和水稻HMA 蛋白序列进行BLAST和筛选,在番茄基因组中鉴定到8 个SlHMAs,这与拟南芥和水稻中的HMA 基因数量相近。为了确定其与拟南芥和水稻HMA 基因的进化关系,根据3 个物种HMAs 的蛋白序列全长,利用MEGA11 构建系统进化树。根据它们的同源关系,8 个SlHMAs 依次命名为SlHMA1-SlHMA8,其中,没有SlHMA2 的命名,而SlHMA5分为SlHMA5.1和SlHMA5.2两个同源基因。与前人报道的拟南芥和水稻HMA 基因一致,8 个SlHMAs 也被划分为2 个亚组,即分别为Zn/Cd/Co/Pb P1B-ATPase 亚组(SlHMA1 和SlHMA3)和Cu/Ag P1BATPase 亚组(SlHMA4-SlHMA8)(图1)。
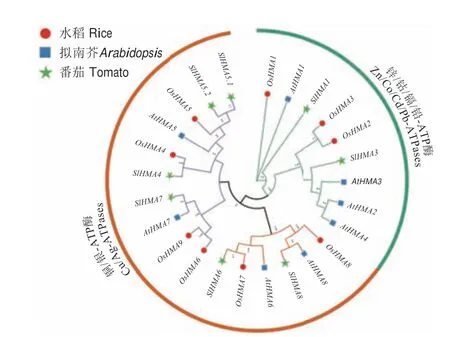
图1 番茄、拟南芥和水稻HMA 基因家族系统进化分析Fig.1 Phylogenetic analysis of HMAs in tomato(Solanum lycopersicum L.),Arabidopsis and rice(Oryza sativa)
蛋白理化性质分析表明,番茄HMA 基因的氨基酸数目介于698-1 302,对应的分子量介于76.04-140.44 kD,大多数SlHMAs 的等电点为5-6,为酸性蛋白,而SlHMA3 的等电点为6.94,SlHMA6和SlHMA1 则分别高达8.30 和8.44,为碱性蛋白。SlHMA3 的不稳定指数较高,达45.63,因此,其蛋白稳定性较差,而其他SlHMAs 的不稳定指数均低于40,为稳定蛋白。类似地,大部分SlHMAs 都为弱疏水性蛋白(0.092-0.324),仅有SlHMA3 表现为弱亲水性(-0.231)。蛋白亚细胞定位预测显示,SlHMA8、SlHMA6 和SlHMA1 定位于叶绿体,其他SlHMAs 含有6-10 个不等的跨膜结构域,均定位于细胞质膜上(表2)。
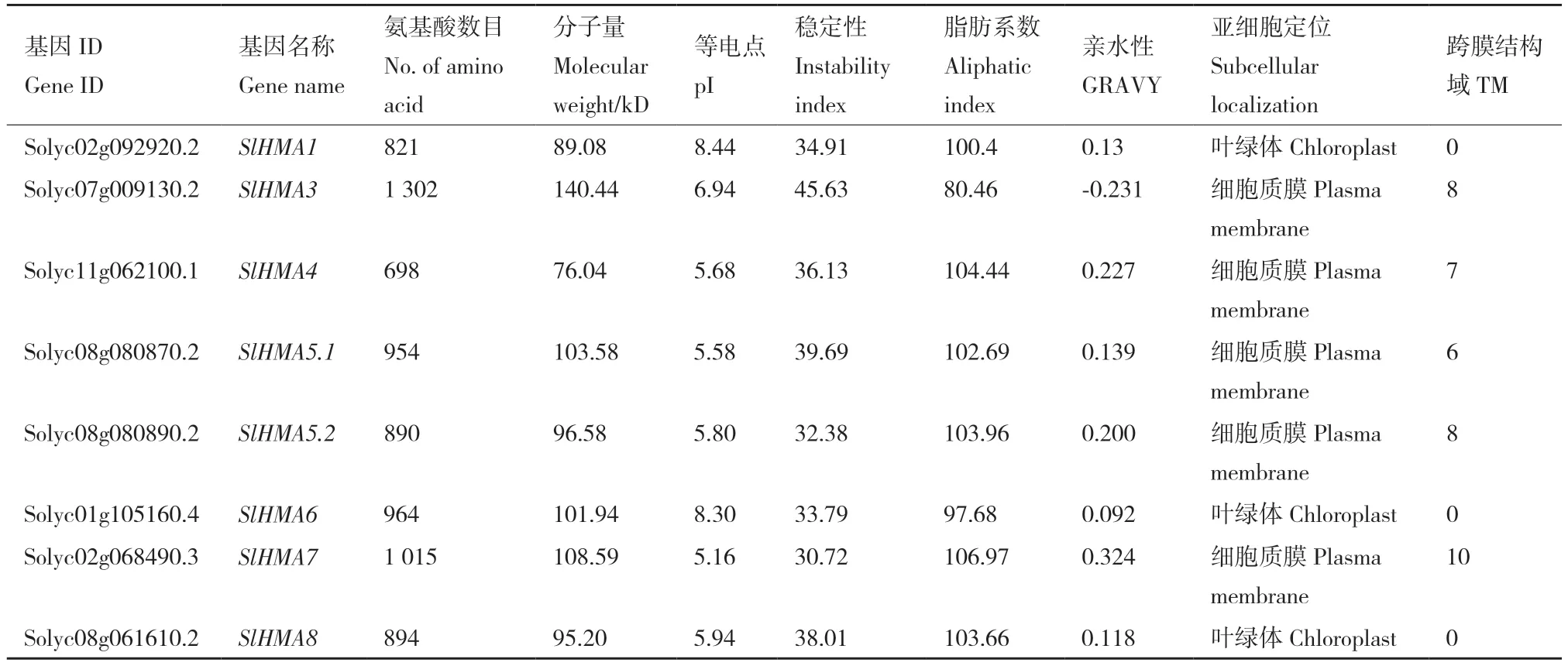
表2 SlHMAs 蛋白理化性质分析Table 2 Physicochemical properties of SlHMAs protein
2.2 SlHMAs染色体定位分析及物种间共线性分析
染色体定位分析发现,8 个SlHMAs 不均匀分布于5 条染色体,其中第8 染色体最多,有3 个HMA基因,分别为SlHMA8、SlHMA5.1 和SlHMA5.2,其次第2 染色体有2 个HMA 基因,分别为SlHMA7和SlHMA1,其余3 个基因分别位于第1 染色体(SlHMA6)、第7 染色体(SlHMA3)和第11 染色体(SlHMA4)。从染色体定位图也可看出,SlHMAs 大多位于染色体3'端,只有SlHMA3 位于5'端。另外,SlHMA5.1 和SlHMA5.2 以基因簇的形式存在于第8 染色体3'端,表明二者可能存在串联重复事件(图2)。
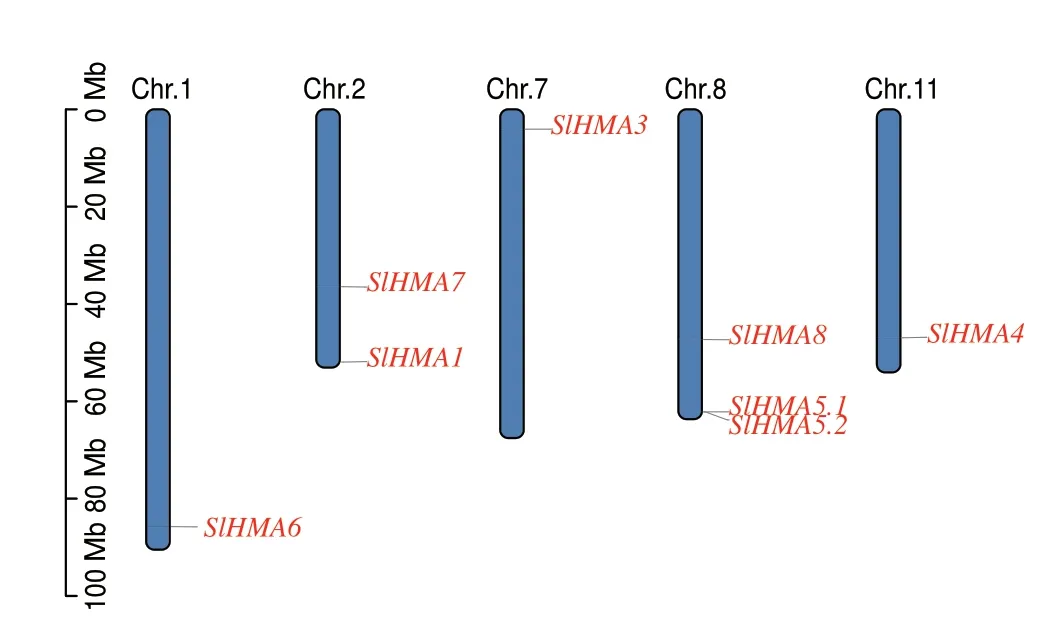
图2 SlHMAs 家族成员的染色体定位Fig.2 Chromosomal localization of SlHMAs family memebrs
番茄与拟南芥、水稻基因组共线性分析表明(图3),番茄各有3 个SlHMAs 与拟南芥和水稻存在共线性,其中,SlHMA5.1 和SlHMA7 在拟南芥和水稻中均存在直系同源基因,表明这两个基因在双子叶和单子叶植物进化过程中较为保守。此外,SlHMA1 在拟南芥中、SlHMA6 在水稻中也各存在一个直系同源基因。

图3 HMA 家族成员在番茄、拟南芥和水稻基因组中的共线性分析Fig.3 Collinearity analysis of HMAs among tomato,Arabidopsis,and rice
2.3 SlHMAs基因家族成员基因结构和序列保守性分析
为了解SlHMAs 家族成员的基因结构特征,对番茄、拟南芥和水稻HMA 基因的内含子和外显子进行了比较分析。结果表明,SlHMAs 序列长度为3 458-21 556 bp,其中最长的为SlHMA8,SlHMA6和SlHMA3 次之,最短的为SlHMA5.2。所有SlHMAs的CDS 均被数量不等的内含子分隔成多个外显子,外显子数量为5-17 个。进一步分析发现,除了SlHMA5.1、SlHMA5.2 和SlHMA4 的基因 长度与其拟南芥和水稻中的同源基因类似或更短外,其他SlHMAs 长度均比其各自同源的AtHMAs 和OsHMAs更长,尤其是SlHMA8 和SlHMA3,二者的基因长度甚至超过其同源基因的2 倍。总之,SlHMAs 在外显子长度和内含子数量等方面,与其同源的AtHMAs 和OsHMAs 无显著差异,而内含子长度则是造成物种间同源基因的长度产生显著差异的主要原因(图4-A)。

图4 番茄、拟南芥和水稻HMA 家族成员的基因结构和保守基序分析Fig.4 Gene structure and conserved motif analysis of HMAs in tomato,Arabidopsis,and rice
通过对蛋白保守基序的比较分析发现,所有的SlHMAs 中共存在5 个保守基序,其中,Cu/Ag P1B-ATPase 亚组的所有SlHMAs(除 了SlHMA4)和Zn/Cd/Co/Pb P1B-ATPase 亚组的SlHMA1/AtHMA1/OsHMA1 均含有全部5 个保守基序,而其中的SlHMA4 缺失基序4 和基序5;Zn/Cd/Co/Pb P1B-ATPase 亚组的其他HMA 都含有4 个保守基序(基序1-4),但其中SlHMA3 的同源蛋白AtHMA3 缺失基序4(图4-B-C)。SlHMA4 与其同源基因在保守基序上的差异表明其在功能进化上可能与其同源基因产生了分歧。
2.4 SlHMAs启动子区顺式作用元件分析
如图5 所示,番茄各个HMA 基因启动子均含有多个不同类型的顺式作用元件,其中光响应顺式作用元件数量最多,分布最为广泛,在所有SlHMAs 启动子区均有分布,表明SlHMAs 均受光信号调控。此外,逆境响应相关的顺式作用元件,如厌氧诱导元件、干旱诱导元件、脱落酸(ABA)响应元件、低温响应元件也有广泛分布,至少存在于5 个以上的SlHMAs 启动子中。而发育过程相关的元件,如生长素响应元件、赤霉素响应元件、分生组织表达元件及细胞周期调控元件等数量较少,仅存在于少数几个SlHMAs 启动子区。结果表明,SlHMAs 更多地参与抗逆反应过程。

图5 SlHMAs 家族成员的启动子顺式作用元件分析Fig.5 Analysis of cis-acting elements in the promoter of SlHMAs
2.5 镉处理下SlHMAs的表达量分析
为了探讨SlHMAs 成员对Cd 胁迫的响应特征,通过RT-qPCR 技术对其开展研究。结果(图6)发现,在番茄根系或叶片中,几乎所有SlHMAs 的表达水平都受镉胁迫调控。具体而言,镉胁迫下,SlHMA5.2和SlHMA7 在根系和叶片中均表达上调,而SlHMA5.1 在根系和叶片中表达均受到抑制。SlHMA1 和SlHMA8 在根和叶中的表达趋势则相反,亦即在根中上调表达而在叶中下调表达。SlHMA4 和SlHMA6 则分别仅在叶片或根系中上调表达,而SlHMA3 在镉胁迫初期上调表达,而在镉胁迫后期其表达受到显著的抑制。揭示SlHMAs 家族基因可能在番茄应对镉胁迫时发挥重要作用。
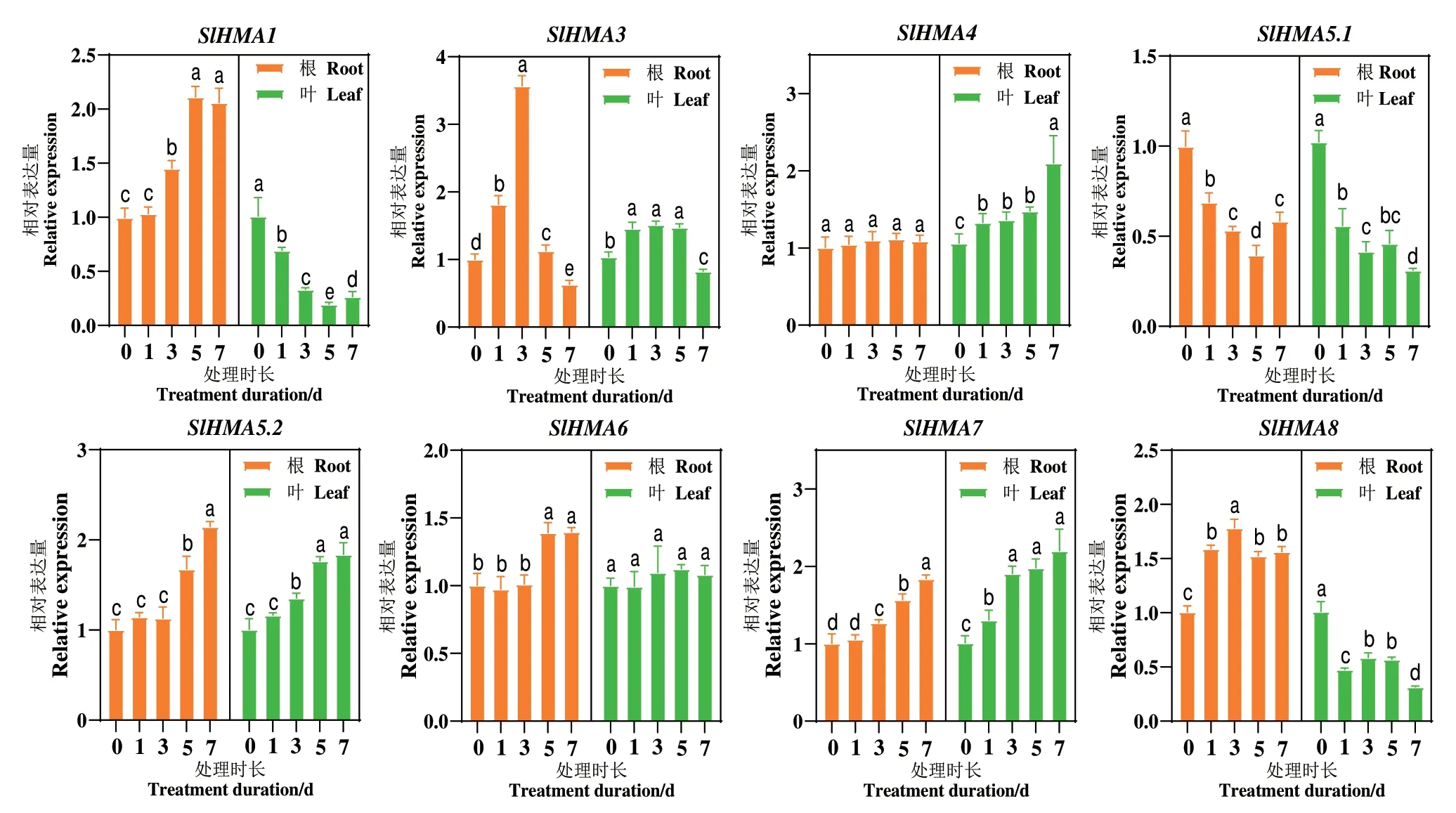
图6 镉胁迫下SlHMAs 家族成员在番茄根系和叶片中的表达模式Fig.6 Gene expression pattern of SlHMAs in the root and leaf of tomato under cadmium stress
2.6 SlHMA1镉转运功能验证
基于以上RT-qPCR 结果,SlHMA1 的表达显著受镉胁迫调控,且SlHMA1 属于Zn/Co/Cd/Pb P1B-ATPase 亚类成员,推测其可能具有镉转运活性,并在镉胁迫中发挥重要作用。因此,克隆了SlHMA1 全长CDS 序列,并转化至酵母镉敏感型突变体Δycf1 中。结果表明,在正常生长条件下,过表达SlHMA1 与转入空载体(EV)的酵母菌株生长状态良好,无显著差异。而在20 μmol/L CdCl2处理下,过表达SlHMA1 的酵母生长明显受到抑制,而转入空载体的酵母菌则尚能正常生长,表明SlHMA1 增强了酵母对镉的敏感性(图7-A)。在5 μmol/L CdCl2的液体培养基中,相比于转入空载体的酵母菌,过表达SlHMA1 的酵母生长也同样受到显著抑制(图7-B)。收集酵母菌体,检测镉含量发现,过表达SlHMA1 显著提高了酵母菌体内的镉含量,进一步表明SlHMA1 具有镉转运活性(图7-C)。
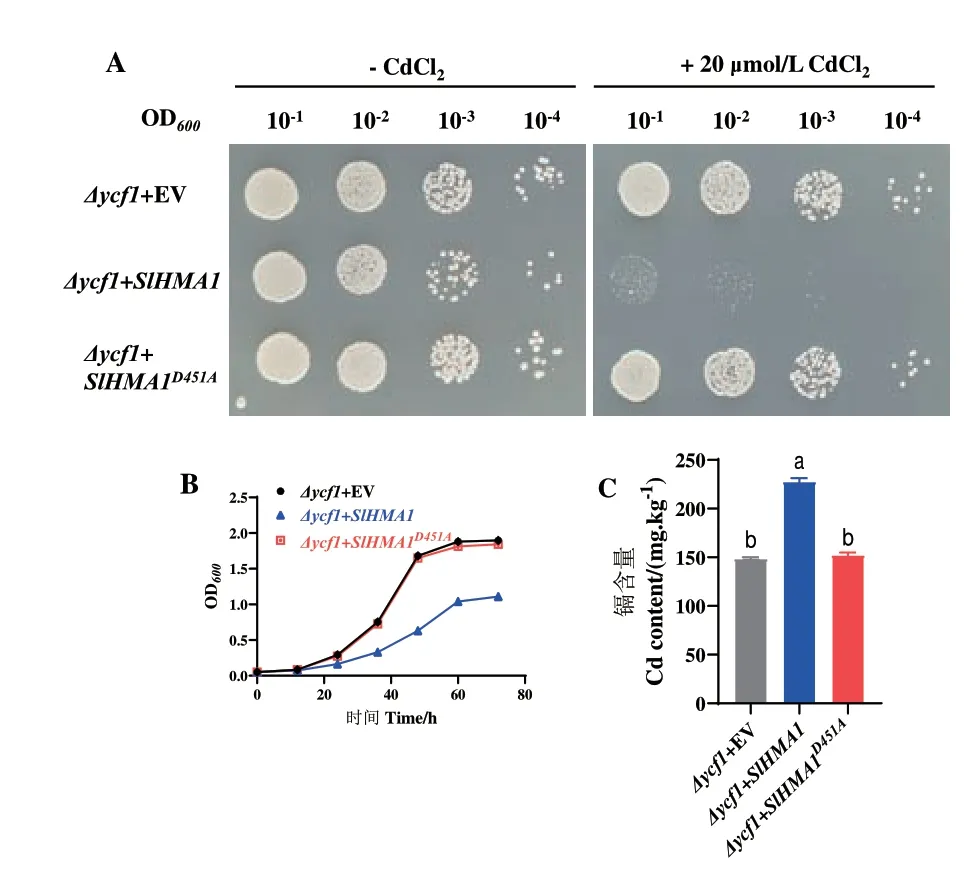
图7 SlHMA1 在酵母中的镉转运活性分析Fig.7 Cadmium-transporting activity of SlHMA1 in yeast
2.7 HMA1基因的进化分析
鉴于SlHMA1 在酵母镉转运和镉抗性中的重要功能,在Phytozome 数据库中,通过氨基酸序列同源比对鉴定了22 个代表性植物物种中的SlHMA1 同源基因。结果表明,从低等的苔藓植物(Marchantia polymorpha、Physcomitrella patens)、蕨类植 物(Selaginella moellendorffii)到高等的被子植物(如大豆、苜蓿等)中,均广泛存在SlHMA1 的同源基因,表明SlHMA1 在植物界中具有广泛的保守性(图8-A)。通过氨基酸序列比对发现,DKTGT 基序在所有代表性物种中高度保守(图8-B)。把SlHMA1 中DKTGT 基序的磷酸化位点D(Asp,天冬氨酸)突变为A(Ala,丙氨酸),并转化酵母后发现,不同于野生型的SlHMA1,过表达SlHMA1D451A的酵母与转化空载体的酵母相比,生长状况和体内镉含量均无显著差异,说明该位点对于SlHMA1 的镉转运活性也至关重要(图7)。同时,结果也表明,保守基序DKTGT 中的天冬氨酸位点可能对植物界中所有HMA1 蛋白的镉转运活性都具有广泛保守的功能。
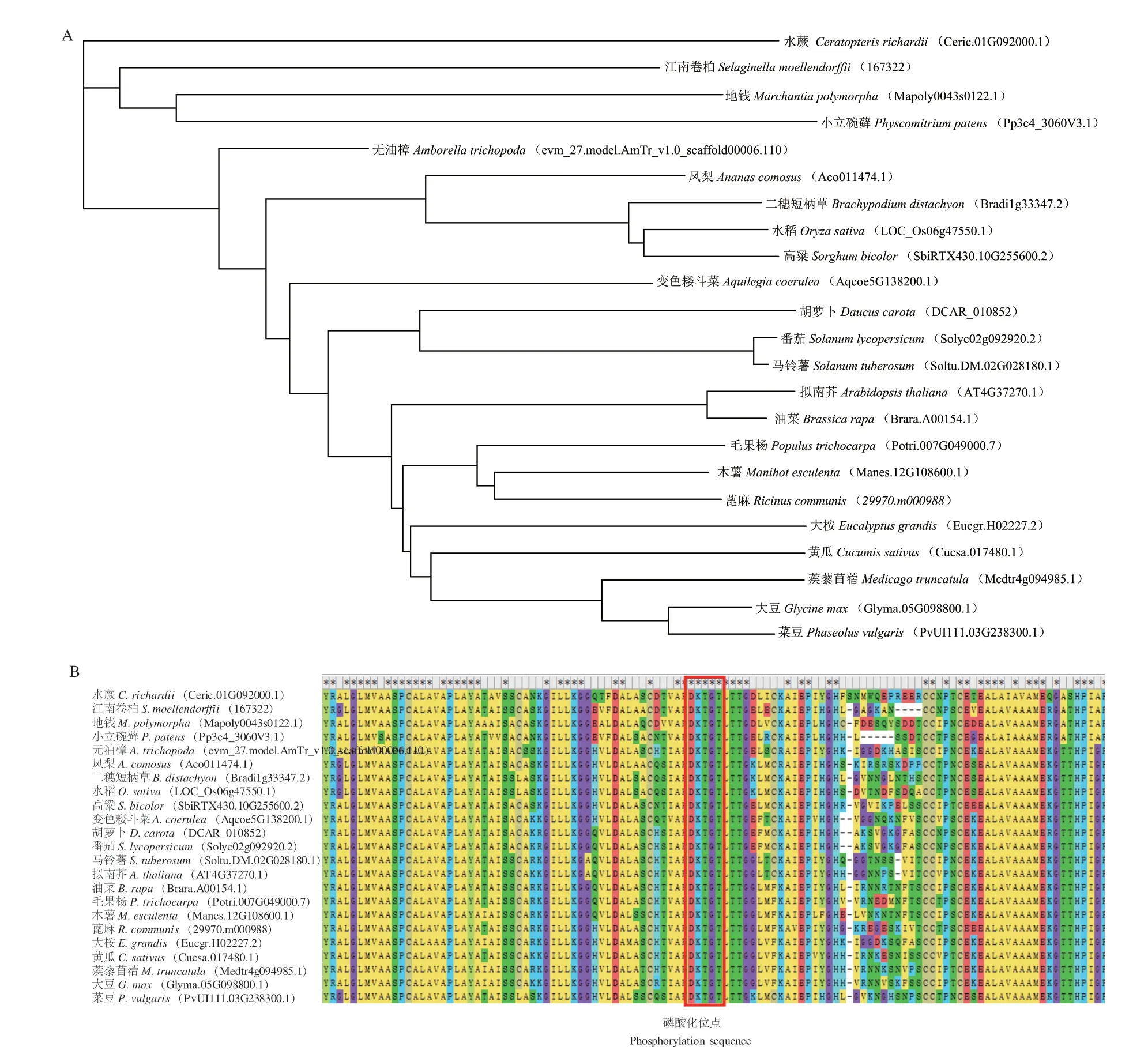
图8 HMA1 在23 个代表性植物物种中的进化分析Fig.8 Evolutionary analysis of HMA1 in 23 representative plant species
3 讨论
HMA 家族基因直接参与体内多种重金属离子的主动转运,是植物维持体内重金属离子平衡的一个重要转运蛋白家族。本研究以番茄这一果实模式植物为材料,从全基因组水平鉴定到8 个SlHMAs,这与前人在拟南芥(8 个)[15]、水稻(9 个)[18]、苜蓿(9个)[19]等二倍体植物中报道的数量相接近。有趣的是,它们基因组大小差异悬殊,分别为785 Mb[23]、135 Mb(www.arabidopsis.org/)、500 Mb(http://plants.ensembl.org/Oryza_sativa/Info/Index)和410 Mb(https://phytozome-next.jgi.doe.gov/info/Mtruncatula_Mt4_0v1),表明HMA 家族基因数量与基因组大小无显著正相关性。然而,在多倍体植物中,大豆(Glycine max,四倍体,20 个)[24]、油菜(Brassica napus,四倍体,31 个)[25]和芥菜(四倍体,27 个)[21]等的HMA 家族基因数量都远高于二倍体植物,表明染色体多倍化事件在HMA 家族基因数量扩张过程中发挥更加重要的作用。
已有大量研究表明,外显子-内含子的结构对于基因家族的进化及内部各成员之间的功能分化发挥重要的作用,而内含子数量及长度等对真核生物的基因表达也有至关重要的调控功能[26-28]。研究发现,在Cu/Ag P1B-ATPase 亚组内各成员的内含子数量为4-16 个不等,而内含子长度差异也较大,暗示该亚组内的成员之间基因功能差异比较大。此外,大部分SlHMAs 的外显子数量和长度都与其拟南芥和水稻中同源基因较为接近,然而,SlHMAs 的内含子长度却普遍长于其同源基因,表明在物种进化过程中,为适应其生长环境,番茄HMA 基因家族进化出了更加复杂的功能和调控机制。
HMA 家族基因是一类P1B型ATPase,在植物中通过水解ATP 提供能量,从而跨膜转运重金属离子[16]。进化分析表明,拟南芥有4 个HMA 基因(AtHMA1-AtHMA4)属于Zn/Co/Cd/Pb P1B-ATPase 亚类,具有潜在的镉转运活性,然而,番茄仅有2 个HMA 基因(SlHMA1 和SlHMA3)属于此亚组。已有大量研究表明,拟南芥、水稻、玉米和高粱等多个不同物种的HMA3 蛋白都定位于液泡膜,参与将根细胞中的Cd 区隔化到液泡中,从而缓解Cd 毒性,并限制Cd 从根系向地上部的转运,最终提高植物的镉耐受性[22,29-32]。基于此,推测SlHMA3 可能也发挥类似的功能,然而该推测需要今后开展更多的实验进行验证。镉胁迫下,SlHMA3 的基因表达在根系和叶片中都呈现先上升后下降的趋势,表明其可能存在复杂的反馈调控机制。相比而言,目前,关于HMA1 在镉胁迫响应过程中功能的研究则较为匮乏。酵母功能互补试验表明,SlHMA1 在酵母中过表达增强了酵母对镉的敏感性,同时酵母体内镉含量显著增加,表明SlHMA1 具有镉转运活性。拟南芥和伴矿景天(Sedum plumbizincicola)中的研究表明,HMA1 蛋白的保守基序DKTGT 中,磷酸化的天冬氨酸(Asp,D)位点对HMA1 的ATPase 酶催化活性和金属转运活性至关重要[33-34]。将保守基序DKTGT 中的氨基酸D 突变成A 之后,SlHMA1 则丧失了其镉转运活性。这与前人在伴矿景天中的研究结果一致[34]。本研究发现,HMA1 在从低等的苔藓植物到高等的被子植物中都广泛存在,说明其可能在早期的植物演化中就已经存在,并且通过繁殖和遗传机制得以保留,在帮助植物更好地适应环境并存活过程中发挥着重要功能。蛋白比对结果表明,SlHMA1 氨基酸序列中的保守基序DKTGT 在所有代表性物种中都高度保守,同时,番茄、拟南芥和水稻所有HMA 家族成员的保守基序motif 1 中也都存在DKTGT 基序。这一结果表明,ATP 水解酶活性相关的DKTGT 基序不仅对所有代表性物种中HMA1的镉转运活性至关重要,同时对HMA 家族其他成员的金属转运活性也至关重要。此外,镉胁迫诱导SlHMA1 在根系中表达,抑制其在叶片中表达,而根系则是植物接触并吸收镉的主要部位,表明在遭受镉胁迫时,SlHMA1 可能将更多的镉固存于根系,同时减少叶片对镉的吸收,从而减少镉对叶片的毒害作用,保护叶片的正常生理功能和植株的存活,未来利用SlHMA1 过表达株系或功能缺失突变体等遗传材料开展相关试验将为该推测提供更加明确的证据。总之,本研究将为探索HMA 基因家族在植物响应重金属胁迫过程中的分子机制提供重要的数据支撑,也将为利用基因编辑技术选育低镉积累作物品种提供重要的理论依据。
4 结论
番茄SlHMAs 具有HMA 家族基因的典型特征,同时也存在功能多样性。SlHMAs 及其氨基酸保守基序DKTGT 与金属离子转运及镉胁迫响应密切相关。
