CRISPR/Cas9 介导的adeG 基因敲除大肠杆菌细菌模型的建立
2024-03-12朱恬仪孔桂美焦红梅郭停停乌日汗刘翠翠高成凤李国才
朱恬仪 孔桂美 焦红梅 郭停停 乌日汗 刘翠翠 高成凤 李国才
(1.扬州大学医学院,扬州 225100;2.江苏省人兽共患病学重点实验室,扬州 225100)
鲍曼不动杆菌(Acinetobacter baumannii,AB)是临床常见的非发酵革兰阴性杆菌[1],也是引起医院内感染的重要条件致病菌之一。其具有较强的感染力及耐药性[2],常导致严重肺炎、血流感染及尿路感染[3]。2017 年,世界卫生组织(WHO)将耐碳青霉烯类鲍曼不动杆菌添加到优先研究目录中,以开发针对该物种的新抗生素或策略[4]。在过去的几十年中,抗生素挽救了无数生命[5],但广谱抗生素在医学和畜牧业中的过度使用导致了细菌的快速进化以及超级细菌的出现[6-7]。同时抗菌素耐药性(antimicrobial resistance,AMR)的出现和全球流行更是对全球公共卫生构成了严重威胁[8]。然而,开发新药不仅成本高,研究耗时长[9],且对抗生素的研发速度远远不及细菌耐药性的产生速度[10],因此需要开发新的抗菌策略来应对AB 日益严重的耐药性。
CRISPR-Cas 系统是指聚簇规则间隔的短回文重复序列和相关Cas 蛋白,它是细菌的固有免疫系统,可识别并防御侵袭的外源核酸。由于其可以识别特异性位点并靶向切割DNA 双链,CRISPR-Cas系统已成功被应用作为一种基因编辑工具来预防和控制细菌耐药性问题[11]。在过去的研究中,II 型CRISPR-Cas 系统已成为基因组工程应用的首选。它只需要在tracrRNA(反式激活CRISPR RNA)以及crRNA(CRISPR RNA)的介导下表达Cas9 蛋白,并将Cas9 引导到入侵DNA 中的靶位点后就可以进行识别和切割。目前将两个RNA 进一步改进为嵌合的单向导RNA(sgRNA),使得基于Cas9 的基因组编辑变得更加简单[12-13]。Junaid 等[14]发现基于CRISPR-Cas 系统的基因编辑技术在克服AMR 和包括鲍曼不动杆菌在内的不同细菌菌株中发挥了至关重要的作用。Yao 等[15]利用鲍曼不动杆菌的天然I-F型CRISPR-Cas 系统进行基因组编辑,使adh4 基因转录水平被抑制了900 倍。本课题组在前期研究中发现1 株具有完整I-F 型CRISPR-Cas 系统的鲍曼不动杆菌AB43,通过全基因组分析与其耐药表型发现CRISPR-Cas 系统可能通过内源性基因调控抑制耐药基因表达[16-17]。这些结果提示运用CRISPR-Cas 系统来抑制鲍曼不动杆菌耐药性的表达具有可行性。
细菌外排泵是降低抗生素、杀菌剂敏感性的关键机制之一,并导致细菌多药耐药表型的出现[18]。在革兰阴性菌中,与临床相关性最高的是RND 外排泵家族[19]。RND 外排泵由内膜转运蛋白、膜融合蛋白和外膜因子组成,并跨越细菌内外膜,起主动外排药物的作用,这导致多重耐药(multidrug resistance,MDR)鲍曼不动杆菌的出现[19-20]。因此本研究基于RND 外排泵基因adeG 构建耐药模式菌株,拟通过构建CRISPR/Cas9 系统重组质粒,在大肠杆菌中对RND 外排系统带来的耐药性进行抑制,为抑制鲍曼不动杆菌的多重耐药提供新的思路。
1 材料与方法
1.1 材料
1.1.1 菌株及质粒来源 13 株临床鲍曼不动杆菌来自盱眙人民医院,鲍曼不动杆菌参考菌株ATCC 17978 及pCas9 质粒由本实验室保存。
1.1.2 主要试剂 药敏纸片购自杭州微生物试剂有限公司;T-Vecter 试剂盒、全基因组DNA 提取试剂盒购自宝日医生物技术有限公司;PNK 酶、DNA 快速连接试剂盒购自碧云天生物试剂有限公司;Bsa I酶、2×Taq plus Master Mix、大肠埃希菌DH5α 购自南京诺唯赞生物科技有限公司;抗生素均购自阿拉丁生化科技股份有限公司。PCR 引物由南京擎科生物科技股份有限公司合成。
1.2 方法
1.2.1 adeLFGH 基因的PCR 筛选 通过煮沸法粗提13 株鲍曼不动杆菌DNA,并于-20℃保存。PCR引物序列及长度见表1。扩增体系及条件:2×Taq plus Master Mix 12.5 μL,上下游引物各1 μL,模板DNA 1 μL,双蒸水9.5 μL,共25 μL,瞬时离心混匀。95℃预变性5 min,循环参数为:95℃变性30 s,退火30 s,按照1 min/kb 进行72℃延伸,共进行35 次循环,再72℃延伸7 min。1%琼脂糖凝胶电泳检测PCR 产物。

表1 本实验使用引物Table 1 Primers used in this study
1.2.2 药物敏感实验检测AB 耐药性 采用微量肉汤稀释法对收集鲍曼不动杆菌进行药敏检测。随机选择13 株具有完整adeLFGH 的菌株以及参考菌株ATCC 17978 进行药敏实验。所使用药物及缩写为哌拉西林(PRL)、替卡西林(TIC)、头孢他啶(CAZ)、美罗培南(MEN)、多黏菌素E(PE)、庆大霉素(GEN)、妥布霉素(TOP)、四环素(TET)、环丙沙星(CIP)、左氧氟沙星(LVX),根据美国临床和实验室标准化协会(CLSI)2020 指南,检测其对抗生素的MIC。
1.2.3 基于adeG 的RND 外排系统模式菌构建 选择1 株具有完整adeLFGH 且具有多重耐药性的临床鲍曼不动杆菌(AB7),提取其全基因组DNA 进行二代测序并扩增adeG 基因。设计adeG 上下游引物,引物序列及长度见表1。以上述菌株DNA 为模板,PCR 扩增目的基因,扩增产物使用产物纯化试剂盒进行纯化回收。回收后的产物根据T-Vecter 试剂盒说明书进行操作,体系及条件为:T-Vector pMD19 1 μL,纯化目的基因2 μL(约0.23 pmol),双蒸水2 μL,轻轻混匀后加入等体积DNA Ligation Kit,充分混匀,16℃孵育12 h 后转化DH5α 大肠杆菌,并在含有50 mg/L 氨苄青霉素的LB 平板上筛选,挑单克隆PCR 鉴定。将鉴定正确的大肠杆菌命名为adeG-E.coli,药敏实验验证adeG-E.coli 模式菌株的耐药性。
1.2.4 pCas9-sgRNA 重组质粒构建及鉴定 利用sgRNA 设计工具CHOPCHOP[22]设计了3 对靶向adeG 基因的sgRNA 序列。合成的sgRNA 序列长度为20 bp,根据Bsa I 酶切位点,在sgRNA 两端增加黏性末端。添加在正向寡核苷酸链的开头为AAAAC,而添加在反向寡核苷酸链的开头和末端序列分别为AAAC 和G。然后用PNK 酶将配对的sgRNA 寡核苷酸链磷酸化,并使用T4 DNA 连接酶连接到Bsa I 酶解的pCas9 上,最终分别产生pCas9-sgRNA1(adeG)、pCas9-sgRNA2(adeG)、pCas9-sgRNA3(adeG)。质粒构建过程如图1 所示。
将重组质粒通过质粒热转法转化入感受态大肠杆菌DH5α,转化后的重组菌在37℃孵育过夜,并在含有40 mg/L 氯霉素的LB 平板上筛选。使用每对sgRNA 下游引物为DocR,将位于pCas9 上sgRNA插入位点上游处的一段特异性序列设计为通用上游引物DocF。对筛选出的重组菌落进行PCR 鉴定,确认后保存。
1.2.5 CRISPR/Cas9 介导的adeG 基因敲除大肠杆菌的构建 甘油法制备adeG-E.coli 模式菌感受态,pCas9-sgRNA(adeG)质粒电转入感受态,37℃培养4 h 后涂布于预先配置好的双抗(40 mg/L 氯霉素,50 mg/L 氨苄青霉素)LB 平板上。
1.2.6 CRISPR/Cas9 编辑效率及药敏测定 挑取上述平板上的单菌落进行扩增培养。首次培养时在液体LB 中添加氯霉素和氨苄青霉素,药物浓度同上。扩增培养12 h 后转接至液体MH 培养基中培养至OD600为0.5。将培养菌液进行药物敏感性筛查。与adeG-E.coli 相对比,测定CRISPR/Cas9 编辑效率。同时通过PCR 鉴定adeG-E.coli 中adeG 基因的降解。
2 结果
2.1 adeLFGH基因检测
PCR 结果显示13 株临床分离鲍曼不动杆菌adeL、adeF、adeG、adeH 的携带率均为100%。图2展示了adeLFGH 的PCR 电泳结果。
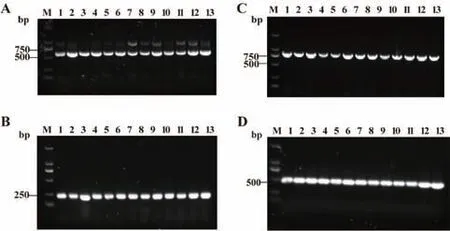
图2 adeLFGH PCR 电泳图Fig.2 PCR gel electrophoresis of adeLFGH
2.2 药敏结果
药敏结果显示(表2,图3),13 株拥有完整adeLFGH 外排泵系统的临床鲍曼不动杆菌对多黏菌素E 敏感,对左氧氟沙星耐药率较低,仅为38.5%,而对头孢他啶的耐药率为92.3%,对妥布霉素耐药率为84.6%,对其他临床常见药物耐药率均为100%,提示具有很强的耐药性。
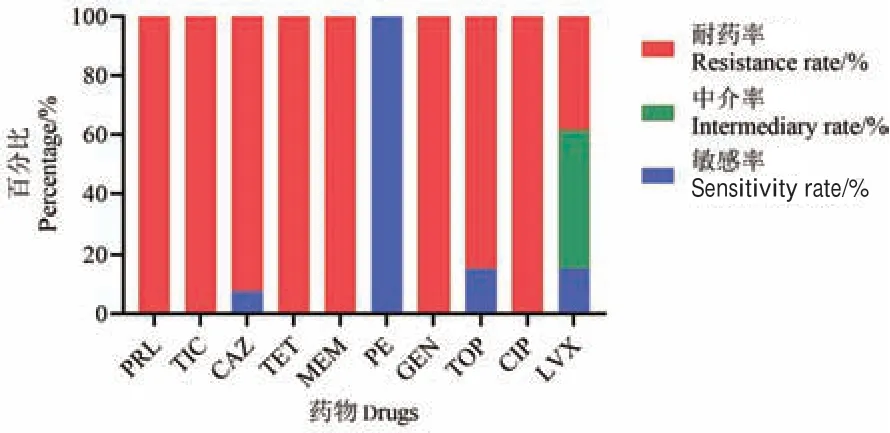
图3 13 株鲍曼不动杆菌对临床常见抗菌药物的耐药百分比Fig.3 Resistance rates of 13 strains of A.baumannii to common clinical antibiotics
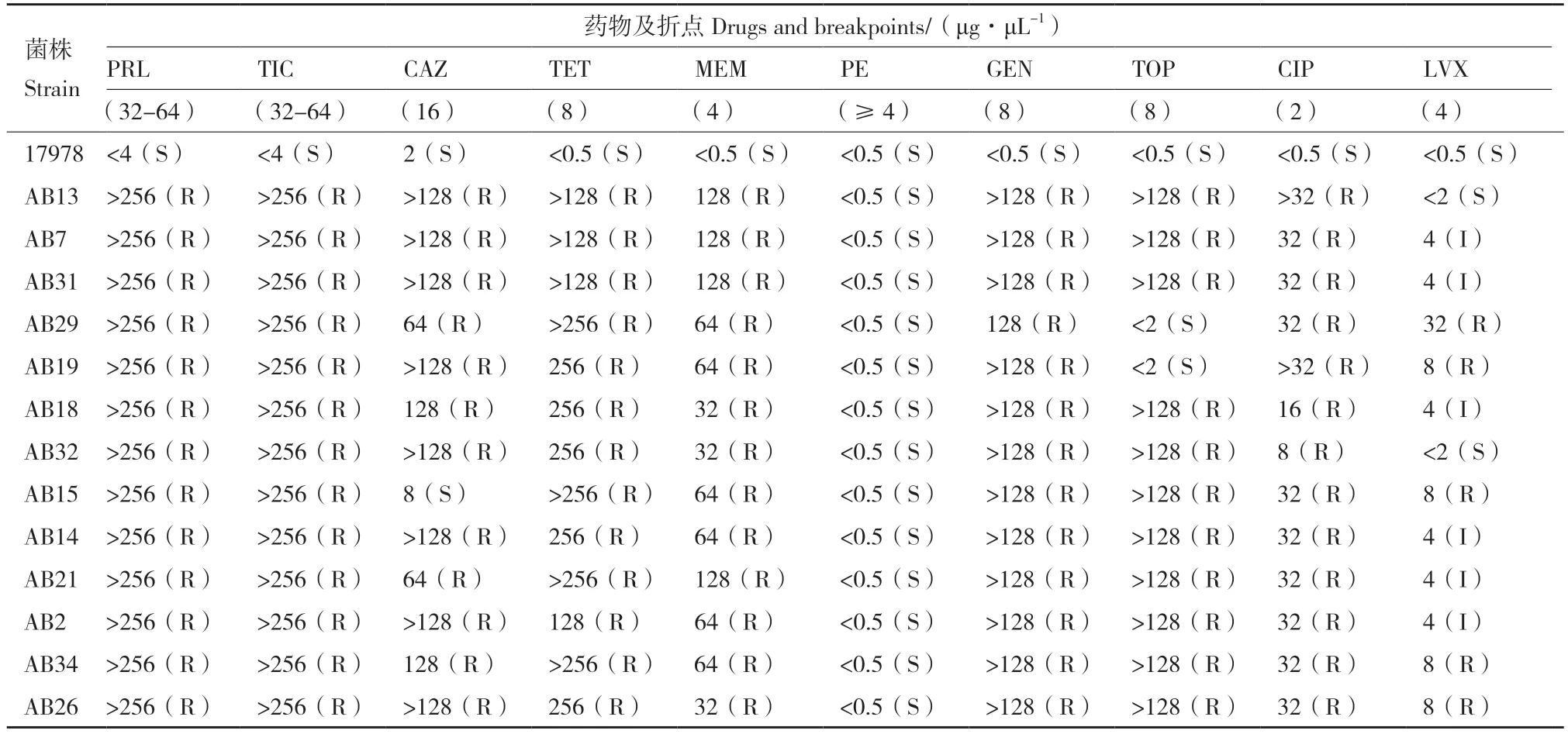
表2 ATCC 17978 和13 株鲍曼不动杆菌对临床常见抗菌药物的药敏结果Table 2 Sensitivity results of ATCC 17978 and 13 strains of A.baumannii to common clinical antibiotics
2.3 基于adeG的RND外排系统模式菌构建及耐药性鉴定
PCR 扩增耐药鲍曼不动杆菌(AB7)的adeG 基因,目的片段大小约为3.3 kb,纯化回收后与T 载体连接,构建T-adeG 质粒。重组质粒通过PCR 鉴定,电泳结果如图4 所示。将其转化至DH5α 后对其耐药表型进行药敏纸片鉴定(图5,表3)。与空白对照DH5α 对比,哌拉西林、替卡西林/克拉维酸以及哌拉西林/他唑巴坦由敏感转为耐药,氯霉素、美罗培南、米诺环素由敏感转为中介。
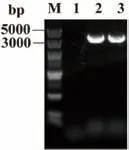
图4 adeG-E.coli 质粒构建PCR 鉴定Fig.4 PCR identification of constructed adeG-E.coli plasmid
2.4 pCas9-sgRNA(adeG)重组质粒构建及鉴定
将合成的sgRNA 磷酸化后与pCas9 质粒连接,转化后挑单克隆进行PCR 鉴定,将鉴定正确的pCas9-sgRNA(adeG) 质粒分别命名为pCas9-sgRNA1(adeG)、pCas9-sgRNA2(adeG) 和pCas9-sgRNA3(adeG)。结果如图6 所示。
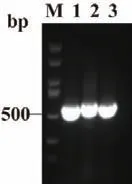
图6 pCas9-sgRNA(adeG)质粒构建Fig.6 Construction of pCas9-sgRNA(adeG)plasmid
2.5 基于CRISPR/Cas9系统的adeG基因编辑效率药敏测定
对pCas9-sgRNA1(adeG)、pCas9-sgRNA2(adeG)和pCas9-sgRNA3(adeG)进行药物敏感性筛查,结果如图7 所示(药敏变化见表4)。在对哌拉西林、替卡西林/克拉维酸、哌拉西林/他唑巴坦3 种纸片进行验证时发现,在pCas9-sgRNA1(adeG)质粒作用下,adeG-E.coli 对于哌拉西林/他唑巴坦(图7-A 中箭头标注)的耐药性得到了逆转,对妥布霉素、四环素、多西环素、米诺环素的耐药性(图7-B、表5)也有不同程度的逆转。另外pCas9-sgRNA2(adeG)和pCas9-sgRNA3(adeG)使adeG-E.coli 对哌拉西林/他唑巴坦由耐药恢复到了中介;而pCas9-sgRNA2(adeG)和pCas9-sgRNA3(adeG)重组质粒对其他药物的耐药性逆转效果并不显著(未展示)。通过PCR 验证adeG-E.coli 中adeG 基因结果(图8)表明,pCas9-sgRNA(adeG)质粒不能完全降解adeG。
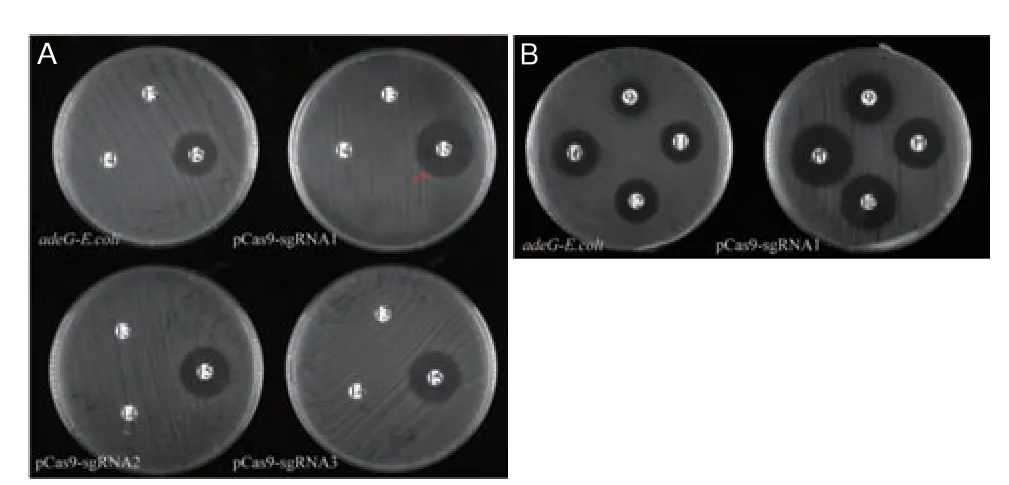
图7 pCas9-sgRNA(adeG)对部分药敏纸片结果Fig.7 Results of pCas9-sgRNA(adeG)on some drug sensitivity paper slides
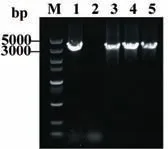
图8 pCas9-sgRNA(adeG)质粒靶向adeG 基因型鉴定Fig.8 Identification of pCas9-sgRNA(adeG)plasmid targeted adeG gene

表4 pCas9-sgRNA 在哌拉西林/他唑巴坦中的药敏变化(与adeG-E.coli 相比)Table 4 Drug sensitivity changes of pCas9-sgRNA in piperacillin/tazobactam(compared to adeG-E.coli)
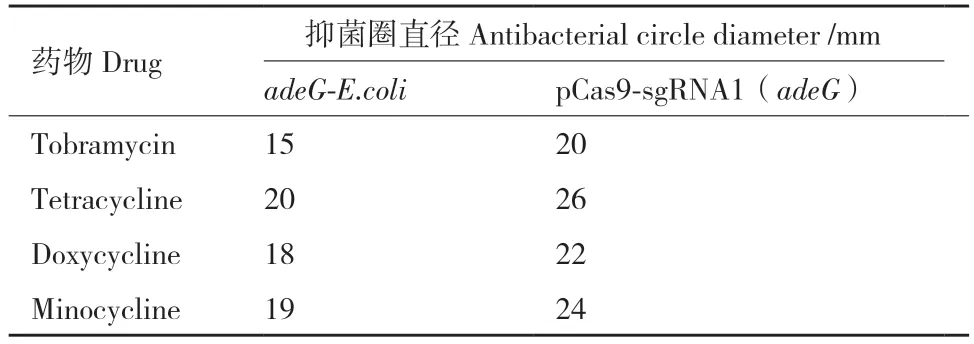
表5 pCas9-sgRNA1(adeG)的部分药敏变化(与adeGE.coli 相比)Table 5 Partial drug sensitivity changes of pCas9-sgRNA1(adeG)(compared to adeG-E.coli)
3 讨论
迄今为止,在鲍曼不动杆菌中已经发现多种类别的外排泵,如RND 外排系统[23]、MFS 外排泵[24]、MATE 家族[25]以及SMR 家族[26]。其中RND 外排系统对于鲍曼不动杆菌多重耐药起到较为积极的作用。RND 外排系统又可分为adeABC、adeIJK[27]和adeFGH 三类[28]。adeABC 受adeRS 双组分系统的严格调控[29];adeIJK 对鲍曼不动杆菌多重耐药作用较小,但与adeABC 具有协同作用[30];而adeFGH 受到其上游LysR 型转录因子adeL 调节,过表达后会赋予细菌多重耐药性[20,31]。鲍曼不动杆菌中adeIJK具有物种特异性,可以稳定表达,而adeABC 和adeFGH 操纵子的检出率分别为80%和90%[20,32]。由于adeFGH 药物外排机制报道较少,且Coyne等[31]对adeG 转运体的跨膜拓扑结构进行了预测并证实其具有RND 蛋白的典型特征。因此在本研究中将来自多重耐药鲍曼不动杆菌的单独外排泵基因adeG 克隆至大肠杆菌DH5α,并观察到了其耐药性的转变,这不仅表明adeG-E.coli 模式菌株构建成功,也进一步证明adeG 对于多重耐药鲍曼不动杆菌的重要性。
pCas9 质粒中的crRNA 与tracrRNA 翻译后形成复合物,可特异性靶向目的基因并募集Cas9 蛋白。Cas9 蛋白是一种RNA 引导的核酸内切酶,它由核酸酶(NUC)瓣和α 螺旋识别(REC)瓣组成[33]。其中NUC 瓣有3 个关键结构域,分别是PI 结构域、HNH 核酸酶结构域和RuvC 样核酸酶结构域。PI 结构域与靶基因中原间隔邻基序(PAM 序列)互补,有助于Cas9 蛋白对靶位点的识别。当Cas9 蛋白识别PAM 序列后,HNH 核酸酶结构域识别并切割与sgRNA 互补的靶链,RuvC 样核酸酶结构域切割对应的非靶链,从而完成Cas9 对靶基因的双链DNA切割,并刺激DNA 修复途径。而REC 瓣在此过程中主要起支撑作用。因此在CRISPR/Cas9 系统靶向敲除中,sgRNA 的选择尤为重要。通过sgRNA 设计网站对AB7 的adeG 基因序列进行分析,选择靶位点位于基因中上游,脱靶率低,预测效率高,且GC 含量处于40%-80%之间的最佳序列[22]。经网站预测及初步筛选获得了85 条sgRNA 序列,根据sgRNA 的设计原则又最终确定了3 条。实验结果证明携带特异性sgRNA 的pCas9 质粒可以对adeG 带来的耐药性进行一定程度的抑制,PCR 结果验证了pCas9-sgRNA(adeG)重组质粒对adeG 的不完全降解。其中pCas9-sgRNA1(adeG)的靶向效率较好,证明通过CRISPR/Cas9 系统敲除细菌耐药基因具有可行性,但要增强敲除效果还需要进一步优化才能与文献报道一致[34-35]。在本实验中,仅pCas9-sgRNA1(adeG)重组质粒对adeG-E.coli 带来的耐药性具有防治效果,证明在sgRNA 的设计上还可以再进行优化,尤其是降低sgRNA 在识别过程中的脱靶率从而增加编辑效率[36]。而在sgRNA 的5'末端添加两个鸟嘌呤核苷酸[37],或者改变sgRNA 与Cas9 的浓度比例[38]也被认为是增强编辑效率的关键。另外,当靶基因存在于细菌基因组中,细菌必须基于同源重组进行修复,换言之,只有基因组被修复的细菌在编辑后才能存活。而本研究构建了基于T-adeG 质粒表达的RND 外排系统模式菌株,避免了由CRISPR/Cas9 系统切割导致菌株死亡从而无法反选编辑成功菌株的问题,同时可以进一步探究CRISPR/Cas9 系统对靶基因表达的抑制能力[39]。但这同时也为CRISPR/Cas9 系统编辑效率带来一些不利因素,如在没有抗生素选择的情况下,质粒会对宿主产生适应性成本[40],可能造成质粒在宿主自然传代下的丢失。或是由于pCas9 质粒与T 载体相比在遗传稳定性、拷贝数差异[41]等影响下带来的竞争劣势,从而最终导致pCas9-sgRNA(adeG)重组质粒基因编辑效率的差异性。Tagliaferri 等[41]研究表明CRISPR/Cas9系统靶向携带blaTEM-1的pSB1A2 质粒后可以明显逆转其基因表型,但并不能完全消除靶基因。这可能解释了为什么pCas9-sgRNA(adeG)重组质粒可以对adeG 带来的部分耐药表型进行逆转,但没有彻底降解adeG。
由于临床鲍曼不动杆菌日益严重的耐药性,以及原生菌株与获得性质粒在多个时间维度上存在相互作用、生长和竞争关系[40],使得直接应用CRISPR/Cas9 系统来防治鲍曼不动杆菌变得困难。在接下来的研究中,将对pCas9 质粒元件进行优化,使其可以真正成为防治多重耐药性鲍曼不动杆菌的有力工具。在本研究中,建立含有鲍曼不动杆菌耐药相关基因的模式菌株对逆转AB 的耐药性进行了初步探索,诚然在DH5α 工程菌中无法很好地还原鲍曼不动杆菌的复杂耐药环境,但这种方式为进一步高效抑制鲍曼不动杆菌的耐药性提供了实验基础,从而帮助更快实现利用CRISPR/Cas9 系统对强耐药鲍曼不动杆菌的有效防控。
4 结论
本研究通过构建鲍曼不动杆菌adeG 耐药基因模型,对CRISPR/Cas9 系统特异性敲除耐药基因进行探索。结果显示鲍曼不动杆菌耐药基因adeG 使DH5α 产生了对哌拉西林、替卡西林/克拉维酸和哌拉西林/他唑巴坦的耐药性,用特异性pCas9-sgRNA(adeG)重组质粒靶向治疗后,部分耐药性发生了改变,并恢复到了敏感。
