森林木质残体微生物群落构建机制研究进展*
2024-03-12赵杼祺胡振宏黄志群
赵杼祺 胡振宏, 何 鲜 黄志群
(1. 西北农林科技大学资源环境学院 农业农村部西北旱地农业绿色低碳重点实验室 杨凌 712100; 2. 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室 杨凌 712100; 3. 福建师范大学地理科学学院 福州 350007;4. 湿润亚热带山地生态国家重点实验室培育基地 福州 350007)
木质残体,涵盖了从碎根残片、小枯死树枝的细木质残体到枯立木及倒木的粗木质残体(郭剑芬等,2011;Baniet al., 2018;蒋雨芮等, 2020),是森林生态系统的重要碳库(杨玉盛等, 2005),占森林植被碳储量的14%及森林植物残体碳储量的61%(Panet al.,2011;Magnússonet al., 2016;刘世荣等, 2011)。木质残体分解是森林生态系统中物质循环的基本过程之一,对森林土壤肥力、植被组成、生产力有重要影响。分解时,木质残体会将自身80%~90%的碳释放至大气中(Cornwellet al., 2009),同时归还大量有机碳和养分元素至土壤(Baniet al., 2018)。此外,木质残体为动植物提供养分和栖息地,如某些蕨类植物只能在木质残体中发芽(Turneret al., 2005;Browninget al.,2010)。
影响分解的关键因素包括气候、基质质量及微生物(Swiftet al., 1976),微生物对分解速率影响较大,只有真菌可通过分泌氧化还原酶和水解酶大幅度降解木质素(Boddyet al., 2008;魏玉莲, 2021)。细菌的作用也不容忽视,如杉木树桩分解后期碳损失速率更多受细菌的影响(Huet al., 2017)。以上研究均表明木质残体微生物群落结构对木质残体分解及养分循环具有重要意义。然而,由于缺乏统一的理论指导,木质残体微生物群落影响因素的相关研究结果仍存在分歧。
基于Web of Science核心数据库以及CiteSpace软件,本文对国内外木质残体微生物相关的文献进行了检索及统计分析。数据搜索主题为woody debris和microbial OR microbe OR microorganism,文献类型为“Article”,检索截止时间为2022年11月20日,对检索结果进行去重后得到215篇参考文献,利用CiteSpace软件对文献进行了关键词时间序列分析,分析结果展示了各聚类之间的关系及其在时间序列上的变化(陈悦等, 2015)。时间可视化图谱(图1)表明,该领域高频词大部分出现在1993—2005年之间,其中,两大高频词“粗木质残体(coarse woody debris)”与“分解(decomposition)”均出现在2000年前,随着时间推进,新的高频词不断出现,说明该研究领域仍在起步阶段。同时,该领域不同聚类间各关键词节点联系较弱,如“微生物群落(microbial community)”与“气候变化(climate change)”。此外,现今研究普遍认为树种属性是影响真菌群落构成的最重要因素(Jaroszewiczet al., 2021;Baberet al., 2016;Ferreret al.,2003),但也有研究表明真菌群落多样性在不同树种间的差异较小,树种对真菌群落的组成和多样性的解释度较差(Joneset al., 2019;Baldrianet al., 2016)。同时,近年研究发现,土壤微生物与土壤环境对木质残体微生物群落可能具有重要影响(图1)(Purahonget al., 2018a;Makipaaet al., 2017;Crowtheret al.,2012a),然而目前相关研究没有将土壤与木质残体微生物群落形成过程很好地联系起来。
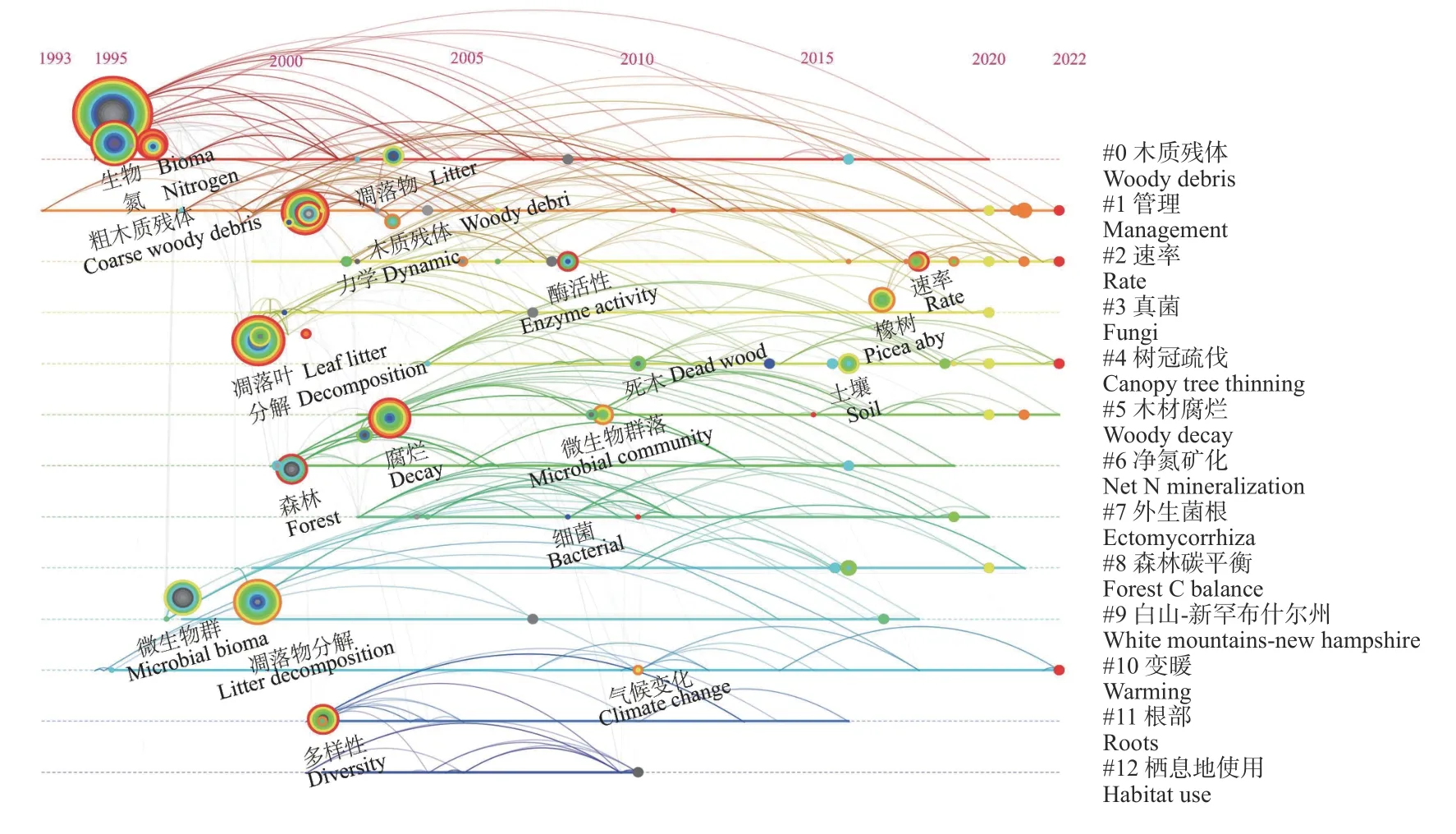
图1 关键词时间序列聚类图谱Fig. 1 Timeline cluster map of keywords
本文首先通过对国内外木质残体微生物相关的文献进行关键词时间序列分析,说明该研究领域的研究现状。在此基础上,探讨了木质残体微生物的来源及其对木质残体的定殖过程的影响,其次围绕土壤、木质残体特性、气候展开,讨论了3者影响下木质残体微生物群落的形成机制(图2)。最后对森林木质残体微生物群落构建机制进行了归纳,并对研究中存在问题进行了展望。
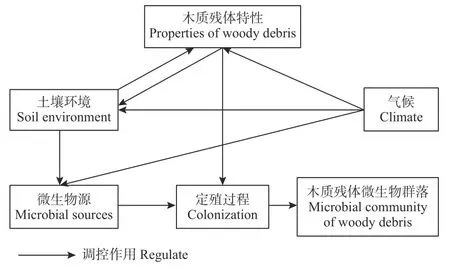
图2 木质残体微生物群落形成机制的概念框架Fig. 2 Conceptual framework of microbial community constructing mechanism in woody debris
1 微生物来源
木质残体的微生物来源为内生菌以及外部环境(如空气、土壤)微生物(Boddy, 2001)。这些定殖者会在木质残体中通过竞争达到群落的动态平衡,进而塑造木质残体微生物群落(Boddyet al., 2016;Boddy, 2001)。
1.1 木质残体
内生菌指栖息在活体植物组织中而不对其造成明显损害的真菌、细菌,包含部分附生物种(Zhengetal., 2016)。内生真菌是木质残体分解早期微生物来源之一,被称为内部定殖者(Fukasawaet al., 2009b)。宿主死亡后,组织发生改变,部分内生真菌菌丝体可以穿透宿主在其表面定殖。此外,内生菌会转变营养模式,成为“先锋降解菌”(Promputthaet al., 2010):如一部分子囊菌(Ascomycota)会在植物死亡后由内生菌转变为腐生菌,从而在分解初期影响来自其他介质的微生物定殖(Rajalaet al., 2012;Promputthaet al.,2007)。但由于大多数内生真菌依赖于更易获得的糖及更易降解的化合物,因此这部分可转变营养模式的内生菌只在分解早期存在,对后续真菌群落的形成与构建影响较小(Songet al., 2017;van der Walet al.,2016;Osonoet al., 2009;Carrollet al., 1983 )。
1.2 大气
大气作为一种传播媒介,微生物含量较低。大气中一些真菌需先产生真菌碎片或孢子才可定殖木质残体(Goodellet al., 2020;Baldrian, 2017)。大气对微生物孢子的传播通常与孢子性质和传播距离有关。短距离内传播通常更有效(Peayet al., 2012;Edmanet al., 2004)。大气传播具有一定的不确定性,但仍被认为是木质残体细菌传播的主要方式(Johnstonet al.,2016),且对稀有真菌的生长以及多样性的维持至关重要(Stenlidet al., 2001)。
1.3 土壤
土壤中微生物多样性丰富、群落组成多样且与木质残体保持稳定接触,因此被认为对木质残体微生物有重大影响(Rajalaet al., 2015)。已有研究发现,木质残体中的微生物有30%以上均来自土壤(Rajalaet al., 2015)。Makipaa等(2017)在挪威云杉(Picea abies)木质残体分解的不同阶段均发现了与土壤真菌种类相同的木质残体真菌,且随着分解的进行,木质残体真菌群落组成与土壤趋于相似。此外,木质残体中部分真菌可以进入土壤。这些真菌进入土壤,改变了土壤微生物结构,为后续木质残体分解中微生物的群落形成提供了更多的可能性(Boddy, 1993)。
2 定殖过程
定殖指微生物在某一环境下可长期生存、繁殖(王以燕等, 2022;Ferreiraet al., 2012)。木材分解的各个阶段均会涉及微生物定殖。植物死亡前,木质残体可能就已被内生真菌定殖,内生真菌被视为第一批定殖木质残体的微生物,即内部定殖者。随时间推移,会有外部定殖者进入木质残体。最初的定殖者会成为后续群落结构和功能的主要决定因素(Lindneret al., 2011; Parfittet al., 2010)。早期资源获取会导致最初的殖民者抢占生态位,进而竞争性排斥后续到达的物种,形成“优先效应”(Vannetteet al., 2014;Chase, 2003;van der Walet al., 2016)。此外,定殖涉及到对资源(即木质残体)的争夺,这种争夺可以分为对初级资源(未被定殖的木质残体)和次级资源(已被微生物定殖的木质残体)的捕获(Boddy,2000),资源的可得性会通过直接影响到达基质的微生物来放大或抑制“优先效应”的强度(Clineet al.,2015)。同时,定殖过程中还存在定殖—竞争的动态平衡。根据竞争排斥原则,物种必须具有不同的生态功能才能共存(Hardin, 1960)。然而,实际微生物群落存在一定功能冗余,缓解了微生物在激烈竞争下多样性的锐减(Smithet al., 2018)。
3 影响木质残体微生物群落形成的主要因素
3.1 土壤环境对木质残体微生物的影响
3.1.1 土壤环境对木质残体微生物源的影响 土壤微生物是木质残体微生物定殖的重要来源,对温度非常敏感(Curtinet al., 2012),一般低温高湿环境更有利于微生物的生长与繁殖(Fravoliniet al., 2018;Liuet al., 2009),但在水分不受限制的地区,土壤温度的升高在一定程度上会增加微生物的生物量(Macdonaldet al., 1995)。pH值同样是影响土壤微生物的重要因素,pH值下降在一定程度上会促进真菌的生长与活性(Ascheret al., 2012;Heinemanet al., 2015;Fravoliniet al., 2016)。研究表明,周围植物根系分泌物的组成和浓度会改变土壤养分浓度,影响土壤微生物的群落组成(Evineret al., 2003;Seneet al., 2012;Beareet al.,1997)。木质残体、凋落物等有机质投入同样会改变土壤养分条件及微生物群落组成(Ayreset al., 2009;Vivancoet al., 2008;Kogel-Knabner, 2002)。
3.1.2 土壤环境对微生物定殖过程的影响 土壤环境会通过影响木质残体的理化性质进而影响定殖过程。如帚石楠(Calluna vulgaris)地块土壤中对微生物生长有抑制作用的多酚物质会扩散到该土壤的木质残体中,改变其化学性质,降低木质残体微生物的丰度(van der Walet al., 2007)。然而,土壤环境是否会通过影响木质残体理化性质改变定殖过程还存在一定的疑问,有研究发现,插入土壤中的木质残体比放置在土壤上方的木质残体分解更快(Rischet al., 2013)然而仍不明确土壤环境是通过影响木质残体理化性质还是通过影响土壤微生物进而影响木质残体微生物的分解的(van der Walet al., 2007;Fravoliniet al., 2016)。
3.2 木质残体对其微生物群落的影响
木质残体特性(理化性质、解剖特征)会影响木质残体微生物群落(Cornwellet al., 2009;Boddyet al.,2016),也会影响定殖过程。木质残体死亡初期,由于木质残体特性以及微生物定殖和分解能力的差异,突破基质界限进入木质残体的微生物种类和数量在时间上会有变化(魏玉莲, 2021;Rajalaet al., 2012)。
3.2.1 木质残体N含量与C/N 自然条件下,木质残体中N含量通常为0.5~13.5 mg·kg-1,但不同树种间N含量差异较大,就裸子植物与被子植物而言,前者的N含量可以达到13.5 mg·kg-1,而后者的N含量最高在3 mg·kg-1左右(Cornwellet al., 2009)。木材的高C∶N(一般为200~1200∶1)会限制作为先驱物种的真菌活动,因为其降解酶是富N的,平均C∶N为3∶1(Rinneet al., 2019;Cornwellet al., 2009)。木质残体N含量与微生物丰度通常呈正相关,N含量的增加尤其会促进细菌丰度(Pastorelliet al., 2020),但具体关系受环境影响。一般温带森林木质残体微生物群落受氮限制更明显,而热带森林则较小(Chenet al.,2016)。此外,基质中N含量增加也可能会抑制微生物的丰富度,如有研究表明,温带森林中N含量与真菌丰富度呈现负相关,作者解释为缺N环境中资源配置优化(Purahonget al., 2018b)。
3.2.2 木质残体纤维素、木质素、单宁、密度与解剖特征 纤维素、木质素、单宁均为木质残体重要化学组成(Harrison, 1970)。纤维素主要分解者为担子菌(Basidiomycota)和子囊菌(tursováet al., 2012)。高纤维素含量会促进真菌生物量的增长(Talbotet al.,2012)。木质素有具有复杂的三维结构,极难被微生物分解(Kirket al., 1987),其主要分解者为白腐菌中的部分担子菌。密度对微生物群落的影响的通常是其余理化性质的改变而引起的(Huanget al., 2022)。例如,木质残体密度的降低通常伴随着含水量的增加,进而促进细菌的生长(Arnstadtet al., 2016;Huet al., 2017)。
就裸子植物与被子植物的木质残体而言,二者植物导管、木质素、单宁与密度均存在差异(表1),被子植物木质残体更利于微生物生长(Weedonet al.,2009)。其细胞管道和连接程度优于裸子植物,一些大直径菌丝在其中受限较小(Cornwellet al., 2009;Choatet al., 2008)。裸子植物木质素浓度通常在23.3%~38.5%间,高于被子植物的14%~38%(Weedonet al.,2009;Loganet al., 1987),同时,由于裸子植物只含浓缩单宁,对微生物生长的抑制作用会更加强烈(Kogel-Knabner, 2002)。木质残体分解速率与密度呈现负相关,但有研究发现,相同环境下裸子植物比被子植物分解更慢,这可能是裸子植物木质残体中浓度更高、结构更复杂的木质素、单宁等难分解物质造成的(Jaroszewiczet al., 2021;Weedonet al., 2009),说明相较于物理特性,木质残体的分解与微生物的组成可能更多受其化学特性的影响。
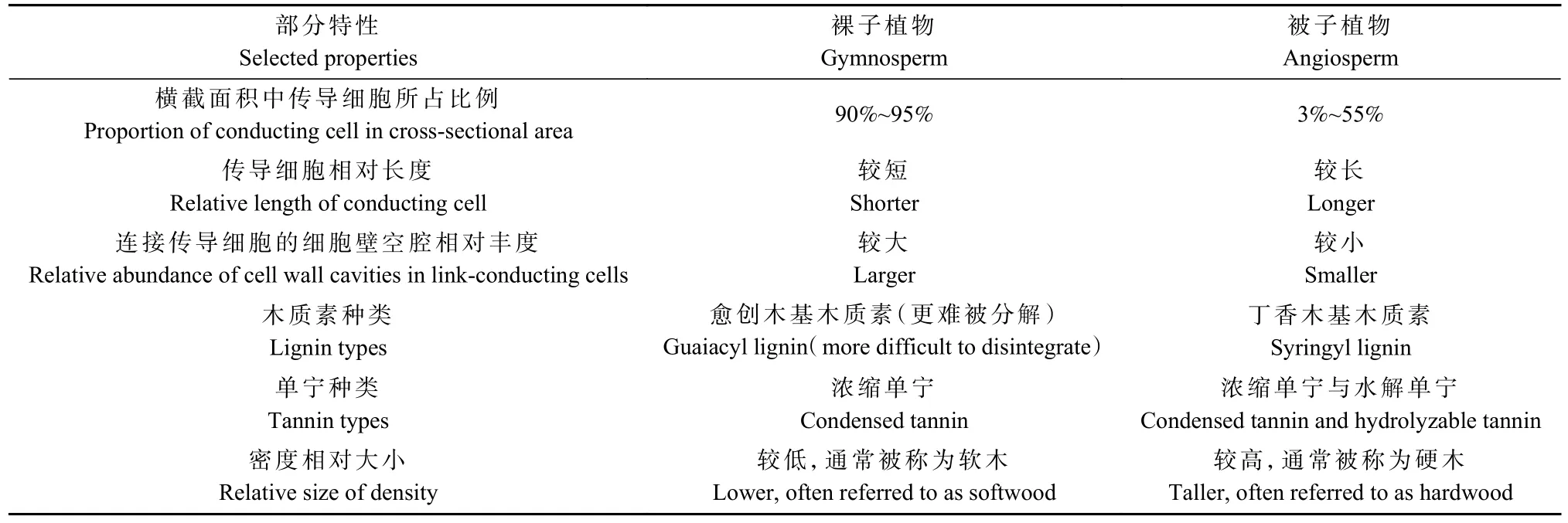
表1 裸子植物与被子植物木质残体部分解剖特征、化学特性及物理特性差异Tab. 1 Differences in anatomical characteristics, chemical and physical properties of woody residue parts of Gymnosperms and Angiosperms
3.2.3 木质残体pH值 pH值是微生物群落构成的重要驱动因素(Mollet al., 2018)。木质残体的pH值普遍在4~6之间,适宜于大部分微生物生长(表2)。木质残体微生物的群落构成与微生物对酸碱性的偏好有关(Mollet al., 2018; Rousket al., 2010)。有学者发现,pH值更低(3.8~4.1)的木质残体分解速度比pH值稍高的(4.3~5.0)木质残体快,因为酸性条件有利于真菌生长,进而促进了木质残体分解(Bewleyet al.,1985; Gomez-Brandonet al., 2017)。木质残体pH值会因微生物活动改变。裂褶菌(Schizophyllum commune)和变色栓菌(Trametes versicolor)等真菌以及梭状芽孢杆菌(Clostridium)、克雷伯氏菌(Klebsiella)等细菌可将木材的pH值改变在适合其生长的水平上(Humaret al., 2001;Johnstonet al., 2016),微生物对木质残体pH值的改变也会诱发后续微生物群落的变化(Gomez-Brandonet al., 2017)。

表2 细菌、真菌与pH值关系Tab. 2 Bacterial and fungal relationships with pH
3.2.4 木质残体直径、体积与物理位置 木质残体直径与体积通过水热条件和比表面积影响微生物定殖(Nordenet al., 2004)。体积较大的木质残体可为微生物提供更多的空间,促进定殖(Baessleret al., 2010)。大直径(大于30 cm)木质残体内具有更稳定的水热条件,而小直径(10~30 cm)木质残体每单位体积的表面积更大,可给真菌留有更多生存空间(Heilmann-Clausenet al., 2004),但同时养分稀缺,一些真菌如黑边近木层孔菌(Phellinus nigrolimitatus)会减少对小直径木质残体的定殖(Stoklandet al., 2004)。
木质残体的物理位置(枯立木与倒木)通常会间接影响其微生物群落。枯立木比与地面接触面积较大的其余木质残体分解更慢。这主要由于枯立木的水分通常小于20%,较低的水分可用性使枯立木中真菌丰富度下降(Hararuket al., 2020; Bradfordet al.,2021)。此外,由于与土壤接触面积较小,土壤微生物定殖速度也更为缓慢(Boddy, 2001; Přívětivýet al.,2018; Shorohovaet al.2014)。
3.2.5 木质残体分解阶段对微生物群落的影响 木质残体的分解会释放养分并提供新的生态位,促进更多微生物种群的建立(Pastorelliet al., 2020;Purahonget al., 2018a;Hoppeet al., 2015)。在不同分解阶段,木质残体特性差异主要表现在质量(主要指化学性质)上(Swiftet al., 1976)。例如随分解进行,N含量会因对大气N的吸收、真菌的转移、细菌固氮而升高(Melilloet al., 1982;Buetleret al., 2007;Fukasawaet al.,2009a),C/N则会由于微生物对木质素、纤维素等物质的分解而下降(Harmonet al., 1986)。但木质残体分解过程中质量变化并不确定,有研究发现,温带森林中欧洲角树(Carpinus betulus)、橡树(Quercus)等木质残体N含量在分解中呈现出下降或较为稳定的状态,而挪威云杉、欧洲落叶松(Larix decidua)等木质残体的C/N比值显著增加(Purahonget al., 2018b;Boddyet al., 2008)。分解初期第一批定殖的微生物会使木质残体更易与其他微生物接触,一定程度上促进后续分解中微生物丰富度增加(Pastorelliet al.,2020)。子囊菌通常在分解前期占主导地位,这是由于其主要参与纤维素等易分解物质的分解,可迅速定殖木质残体。随分解进行,易分解物质被不断消耗,木质素等难分解物质浓度增加,真菌群落会转变为担子菌为主的菌群,并出现菌根真菌(Baniet al.,2018;Rajalaet al., 2012)。同时,真菌群落的变化会引起细菌群落的变化,细菌群落会逐渐转变为适应于真菌环境的细菌为主导(Johnstonet al., 2016)。
参与木质残体早期分解的微生物被认为具有较大的随机性,但同时又对后续群落发展具有重要影响(Dickieet al., 2012;Magnússonet al., 2016;Fukamiet al., 2010),然而有研究发现这种影响持续的时间有限,随分解进行,不同木质残体微生物群落结构及活跃真菌种类均趋于相似(Makipaaet al., 2017;Rajalaet al.,2011)。这是由于其理化性质均与土壤属性逐渐相似(Rajalaet al., 2011)且木质残体特性对微生物群落作用超越了早期殖民者。
3.3 气候对木质残体微生物群落的影响
气候既会影响木质残体微生物的微生物源,又会影响其定殖过程。气候会通过土壤环境影响土壤微生物群落组成,极端温度也会导致空气中孢子的死亡(Norroset al., 2015)。同时,气候也会改变木质残体的水温条件,进而影响微生物的定殖过程(Eichenberget al., 2017;Chazdonet al., 1984)。
3.3.1 2种主要气候因子 气候主要通过湿度与温度影响木质残体微生物(表3)(A'Bearet al., 2014;Olajuyigbeet al., 2012;Boddy, 1983)。真菌对水分可用性依赖小于细菌,因此在基质含水量较低时会处于优势地位(Brayet al., 2012; Liuet al., 2016),而随含水量升高,微生物群落会逐渐转变为细菌为主导(Huet al., 2017)。

表3 两种气候因子变化与微生物的响应Tab. 3 Changes in two climate factors and microbial response
微生物最佳活动温度取决于物种。一般情况下,真菌生长的最适范围在20~40 ℃,一些担子菌温度高于35 ℃时会停止生长(Jannaet al., 2010;Boddy, 1983),此外,真菌对低温的耐受性比细菌强。
3.3.2 大气候与小气候 气候变化带来的湿度与温度的差异在不同温度带下表现明显,纬度变化下微生物群落结构与生物量也会变化(Wuet al., 2009),如Zhou(2011)等发现,从中国寒带森林到亚热带与热带森林,多孔菌(Polyporaceae)的种类及丰富度逐渐增高。热带真菌多样性在所有气候带中最高,其微生物丰富度及稀有真菌种类均高于高纬度地区(魏玉莲, 2021;Větrovskýet al., 2019),这是由于温暖潮湿区域基质可用性更高,可以为木腐真菌提供更加适宜的生存条件(Gomez-Hernandezet al., 2012)。海拔梯度是大尺度下微生物群落变化研究的常用系统,研究表明,随海拔升高,真菌与细菌生物量的比例逐渐增加(Wagaiet al.,2011)。而就多样性而言,微生物多样性通常会随着海拔升高而下降(Gomez-Hernandezet al., 2012;Zhanget al., 2010)。
一些树种通过遮荫、拦截降水等方式形成的小气候也会影响木质残体微生物群落组成。有学者研究发现,大拟射脉菌(Phlebiopsis gigantea)、肉色隔孢伏革菌(Peniophora incarnata)等真菌对遮荫环境具有较为强烈的倾向(Lindheet al., 2004)。而由于微生物的不同响应,微生物群落可能会转变为以适宜严苛小气候下生长的微生物为主导,进而改变微生物群落结构(Krahet al., 2018;Horaket al., 2016)。
3.3.3 全球气候变化 全球气候变化主要表现为全球气候变暖以及极端气候事件(Joneset al., 2022;Tiedjeet al., 2022)。全球气候变化会导致微生物多样性丧失,改变物种分布(Banerjeeet al., 2020)。气候变暖会影响木质残体中早期定殖者,改变后续群落组成(Crowtheret al., 2012b;Hiscoxet al., 2016)。Edman等(2021)发现,外部环境温度的增加加强了早期殖民者对后续微生物群落的影响,使对照与变暖处理下微生物群落具有显著差异。极端天气所引起的自然灾害同样会影响木质残体微生物。森林火灾会直接改变土壤的温度,引起土壤pH值增加、水分含量降低,改变土壤微生物群落,进而影响其对木质残体的定殖(Hartet al., 2005)。在干旱发生时,木质残体含水量的降低会迫使微生物改变资源分配策略(Manzoniet al., 2012;Schimelet al., 2007)。一般认为真菌比细菌耐旱性更强,这是因为真菌可以通过菌丝在土壤中获取水分,因而其丰度也会增加(Schimelet al., 2007)。而在长期干旱的环境条件下,具有与碳氮获取有关功能基因的真菌会具有更强竞争力(Tresederet al., 2018)此外,真菌更易受到冷冻伤害,土壤的冻融会使细菌占据主导,重复冻融会使微生物的生物量显著下降(Nazihet al., 2001;Larsenet al.,2002)。
4 结论与展望
4.1 结论
木质残体的微生物源主要包括残体自身、土壤以及大气,这些微生物对木质残体的定殖塑造了木质残体微生物群落。定殖指微生物在木质残体中增殖或持续存在的现象,定殖过程与木质残体微生物竞争及对资源争夺有关。土壤、木质残体、气候是影响木质残体微生物群落形成的重要因素。土壤理化性质改变会影响土壤微生物群落,也会改变与其接触的木质残体特性。木质残体特性会特性差异会引起定殖与竞争结果差异,较高的N含量、纤维素含量及较大的导管与体积等通常有利于其生长繁殖;较高的密度、C/N、木质素含量、单宁含量通常与微生物的生长呈现负相关。由于细菌与真菌对pH值的适应性差异,pH值升高会促进细菌活动而抑制真菌的活动。此外,木质残体物理位置会通过影响水分可用性影响木质残体微生物群落,与土壤接触面积更大的木质残体具有更丰富的微生物。在不同分解阶段,木质残体特性变化会改变微生物群落。随分解进行,早期的“优先效应”会减弱。大气候、小气候及全球气候变化主要通过温度与湿度影响土壤微生物及大气中孢子传播,改变微生物生长环境的水热差异,造成木质残体微生物群落差异。
4.2 展望
综上所述,未来森林木质残体微生物群落的研究仍需关注以下几个方面:
1) 关注早期定殖者及不同分解阶段土壤微生物的作用。早期群落形成中存在“优先效应”, “优先效应”的影响会持续多久?土壤微生物群落与木质残体微生物群落之间联系密切,但相关研究仍存在空白。在后续的研究中需通过人工模拟和自然模拟实验衡量不同分解阶段土壤微生物的重要性,探究土壤微生物与木质残体微生物的交换机制。
2) 加强木质残体资源变化情况与对应微生物群落中优势物种功能情况的研究。木质残体资源会随分解改变,这与微生物有关。在分解过程中,木质残体理化性质与微生物之间存在相互作用,对二者相互作用的理解需要建立长时间分解实验,研究不同分解阶段中关键物种,将微生物生态功能与分解过程联系起来。此外,当前研究对微生物功能表述模糊,需要细分其功能性状。
3) 将木质残体分解和养分循环与全球气候变化联系起来。木质残体在分解过程中向土壤提供稳定的有机质输入,而全球气候变化则对木质残体分解具有重要影响。在后续研究中需阐明微生物、木质残体分解、全球养分循环的关系,寻找对木材分解、碳氮转移具有重要关键作用的功能微生物,讨论全球气候变化对三者相互作用的影响。同时,木质残体碳储量及分解对全球气候变化响应的研究较少。全球气候变化背景下,木质残体分解加速,而碳储量降低,能否通过木质残体的分解调控全球气候变化背景下的碳循环?这需要未来通过生态模拟过程探究木质残体分解对全球气候变化的响应和反馈机制。
4) 选择更精确的技术辅助研究。如采取单细胞筛选和靶向宏基因组联用等技术更精确地识别微生物功能,获取木质残体微生物群落中的关键种和稀有种资源(Liet al., 2022);采用植物模型系统(可控制变量)类似技术定量研究木质残体主要理化性质与分解过程中主要微生物种类的具体对应关系(Talbotet al., 2012),目前微生物筛菌和培养技术已有较大发展,可将之利用至木质残体分解研究。通过筛选出特定功能微生物,设定不同的控制条件,定量分析功能微生物的定殖和演替过程对群落形成和分解的影响,将微生物群落变化的因素具体化、清晰化。
