贵州北盘江喀斯特地区6种兰科植物结构及其生态适应性
2024-03-08吴天柔安明态余江洪唐煜彬
吴天柔,安明态*,王 琨,余江洪,唐煜彬
(1 贵州大学 林学院,贵阳 550025;2 贵州大学 生物多样性与自然保护研究中心,贵阳 550025)
兰科(Orchidaceae)是有花植物中最大的科之一,很多种具有极高的观赏价值和药用价值,被认为是分类学和系统学发育最多样且进化程度最高的类群,同时在生态系统中拥有不可替代的生态指示作用,多为环境中旗舰物种[1],是生物学研究的热点类群之一。世界约有兰科植物736属28 000种[2],广泛分布于除南北极和干旱沙漠地区以外的陆地生态系统中,尤其是热带地区的兰科植物极为丰富[3]。中国地跨3个气候带(热带、亚热带和温带),地理环境复杂,不仅有各气候带的兰科植物区系,并且还保留着许多原始类群[4],共拥有1 600多种兰科植物,是兰科植物种类最丰富的地区之一[5]。由于兰科植物被过渡采集与繁殖受限导致部分种类濒临灭绝,如今多数已知野生兰科植物均被列入《濒危野生动植物国际贸易公约》(CITES)附录Ⅰ、Ⅱ中[6],占该公约保护植物的90%以上[7],目前对兰科植物保护的研究具迫切性。
兰科植物常生长于土面、石壁、树冠、石缝中,具有不同的生境偏好性,对环境变化较为敏感,常与生境中特定的共生真菌形成较强依赖关系[8]。在漫长的进化过程中,兰科植物已经进化出根、假鳞茎、叶等营养器官的各种内部结构功能和外部形态特征。如叶片有等面叶和异面叶之分,干旱或强光环境中大多数为异面叶,栅栏组织和海绵组织分化、较厚的角质层和复表皮等结构使其更具抗旱特性[9];根系多为粗壮的肉质根,干旱和附生生境中部分兰科植物为气生根结构,除根尖以外,其余部分为死细胞覆盖的根部,能有效避免裸露根系散失水分[10];假鳞茎为兰科植物适应特殊生境的组织构造,由叶节之间的茎膨胀加厚而形成,是兰科植物适应环境变化过程中充当重要的储水和物质储存器官,对兰科植物营养生长和生殖过程发挥着至关重要的作用[11]。各营养器官结构间彼此相互协调适应地生、旱生、附生、岩生以及腐生环境,最大程度降低外界干扰因素对植物本身带来的不利影响,是植物适应环境谋求生存的关键所在,为植物生存发展发挥着重要的生态作用[12]。兰科植物各营养器官结构的存在程度和差异,反映了为适应环境做出的生态策略[13],当前关于兰科植物各营养器官多集中在其叶片结构与抗旱性特征、传粉机制、分类学研究和单种或单器官结构特征研究[9-11,13-17]。综合探究不同生活型兰科植物叶、假鳞茎、根等营养器官结构特征及其与环境间适应关系的相关研究较少,因此探讨兰科植物长期适应环境演化形成营养器官内部结构特征与功能性状值得深入研究。
北盘江是珠江流域西江水系的一级支流,该流域位于中国东南季风与西南季风交汇区,地处云贵高原倾斜地带,为西南喀斯特地貌典型发育区,复杂多样的地形和基岩类型影响植物生境的光照、土壤养分和水分等条件,使其具有高度的异质性、严酷性、脆弱性[18]。
独特的生境内分布249种兰科植物,占据贵州省兰科植物总数的72.59%,是贵州省兰科植物分布最丰富的地区[19],但目前该地区兰科植物营养器官结构的相关研究较少。因此,本研究拟以该地区分布种类较多的6种不同生活型兰科植物营养器官(叶、假鳞茎、根)结构特征为基础,探究不同生活型兰科植物结构特征及其差异,分析种间结构差异与环境之间的适应特性,旨在揭示不同兰科植物响应生境异质的生存策略,为兰科植物研究、保护与发展提供理论支持和参考依据。
1 材料和方法
1.1 试验材料
2021—2022年于北盘江流域喀斯特地貌(24°50′—26°50′N,103°50′—106°15′E)选取分布数量较多或具有代表性的不同生活型6种兰科植物(表1),每种兰科植物采集植株中部健康完整的叶片和假鳞茎,挖取1条离根尖5 cm 长的根,擦拭干净后放置FAA 固定液中保存带回测定。

表1 贵州北盘江喀斯特地区6种兰科植物基本信息Table 1 Basic information of six orchids in Beipan River basin of Guizhou
1.2 试验方法
参照石蜡切片的方法[20],将采集的兰科植物根、假鳞茎、叶从FAA固定液中取出后洗涤干净,用不同浓度的酒精进行脱水、氯仿透明、侵蜡和包埋处理,使用切片机切取根、假鳞茎、叶厚度为8 μm 的横切薄片,番红-固绿染色后用中性树胶封片。使用显微镜进行观察并拍摄横切图,应用CaseViewer、ImageJ软件测量以下指标:(1)叶片结构指标:叶片厚度(LT),上表皮厚度(TU),下表皮厚度(TL),上角质层厚度(TUC),下角质层厚度(TLC),叶肉厚度(MT),维管束直径(DVB1),叶脉厚度(TV),维管束直径占比(PVB1);(2)假鳞茎结构指标:假鳞茎直径(DS),表皮厚度(ET),维管束直径(DVB2),维管束数量(NVB),假鳞茎横截面积(CSAS),维管束密度(VBD),维管束直径占比(PVB2);(3)根结构指标:根直径(RD),根被厚度(RCT),皮层厚度(CT)、维管束直径(DVB3),维管束直径占比(PVB3),叶脉凸起度(VPD)=叶脉厚度(TV)/叶片厚度(LT)×100%。
1.3 数据分析
对于6种兰科植物叶片、假鳞茎、根的结构测量至少选取3个视野,每个视野测量重复5次,所有测量指标均为15个重复。通过SPSS 26.0软件进行数据统计分析,获取均值与标准差,利用单因素方差分析(one-way ANOVA)比较不同兰科植物结构指标之间的差异性,Duncan's多重比较法检验差异显著性(P<0.05)。
通过R语言和OriginPro 2021等软件进行Pearson相关性分析和主成分分析(principal component analysis,PCA)并绘图。
2 结果与分析
2.1 兰科植物结构特征
2.1.1 叶片结构
横切面结果显示,植物叶片结构分为表皮、叶肉、叶脉3个部分(图1)。叶片厚度变化范围为229.89~1 164.05 μm,附生兰(梳帽卷瓣兰,单叶石仙桃)叶厚度差异较大。栗鳞贝母兰与梳帽卷瓣兰为异面叶,表皮细胞均有复表皮结构,为长方形或长圆形细胞,紧密嵌合,无细胞间隙。栗鳞贝母兰有明显的栅栏组织与海绵组织分化,海绵细胞呈圆形或椭圆形,松散排列于栅栏细胞下复表皮之间。梳帽卷瓣兰栅栏组织不明显,海绵细胞呈圆形或类圆形网状,分布于栅栏组织和下表皮之间。莎叶兰、丘北冬蕙兰、兔耳兰和单叶石仙桃为等面叶,均无明显栅栏组织与海绵组织的分化,可明显观察到有较多叶绿体呈串状分布于叶肉细胞中,且细胞排列疏松,有细胞间隙,上下表皮均由单层细胞构成。整体来说不同种间细胞结构存在差异,但同一生活型兰科植物具有一定的相似性,附生兰表现出阳生植物结构特征,地生兰表现出阴生植物特征。
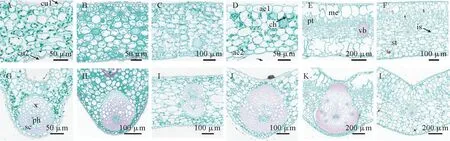
图1 贵州北盘江6种兰科植物叶片(A-F)和主叶脉(G-L)的结构横切面A、G.丘北冬惠兰;B、H.莎叶兰;C、I.兔耳兰;D、J.单叶石仙桃;E、K.栗鳞贝母兰;F、L.梳帽卷瓣兰;cu1.上角质层;cu2.下角质层;ae1.上表皮;ae2.下表皮;me.复表皮;pt.栅栏组织;vb.维管束;st.海绵组织;is.细胞间隙;x.木质部;ph.韧皮部;sc.厚壁组织;ch.叶绿体。Fig.1 Cross section of leaves (A_F) and midrib (G_L) of six orchids in the Beipan River of Guizhou ProvinceA,G.C.qiubeiense.B,H.C.cyperifolium.C,I.C.lancifolium.D,J.P.leveilleana.E,K.C.flaccida.F,L.B.andersonii.cu1,upper cuticle.cu2,lower cuticle.ae1,upper epidermis.ae2,lower epidermis.Me,multiple epidermis.pt,palisade tissue.vb,vascular bundle.st,spongy tissue.is,intercellular space.x,xylem.ph,phloem.sc,sclerenchyma.ch,chloroplast.
丘北冬蕙兰表皮细胞呈三角形或圆形,叶片表面起伏呈波状,叶肉细胞呈圆形或椭圆形,细胞排列疏松且细胞间隙较大,叶绿体均匀分布于叶肉细胞中。上下角质层厚度最大,分别为(11.96±3.05)μm 和(7.80±3.32) μm,上表皮较厚[(25.65±9.86) μm],下表皮较薄[(20.33±4.79) μm],主叶脉厚度最小[(287.53±39.47) μm],叶片整体表现出叶脉细小,叶片机械支撑能力较弱的结构特征。
莎叶兰和兔耳兰在叶片结构特征无显著差异,莎叶兰上下表皮厚度和上角质层最小,分别为(13.55±1.33) μm、(4.40±0.46) μm 和(10.81±1.74)μm,具有较薄的叶片、主脉和下角质层。兔耳兰与莎叶兰相比较,除上表皮细胞较薄[(14.76±3.62)μm]以外,其余指标均较厚。
单叶石仙桃、栗鳞贝母兰和梳帽卷瓣兰3种地生兰叶片存在显著差异(P<0.05)(图2),单叶石仙桃上表皮细胞厚度和主脉凸起度最大,分别为(33.28±5.82) μm 和(220.92±16.87) μm,具有较大的上角质层厚度和主脉凸起度,叶肉细胞和叶片厚度最小,分别为(166.15±17.00) μm 和(229.89±16.04) μm。栗鳞贝母兰主脉维管束直径和下表皮厚度最大,分别为(681.05±13.95) μm和(42.67±2.76) μm,下角质层厚度最小,为(2.47±0.54) μm。
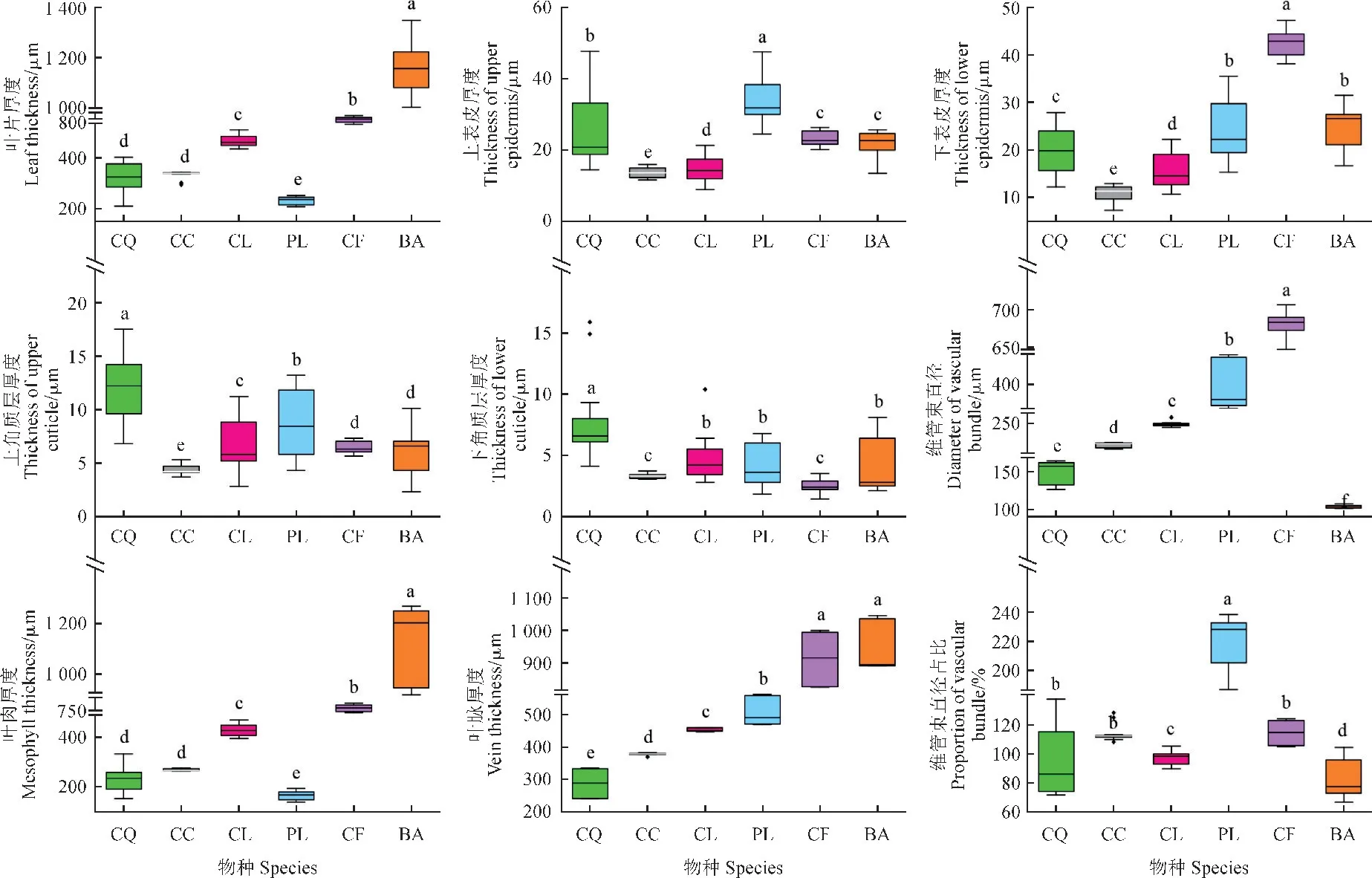
图2 贵州北盘江流域6种兰科植物叶片组织结构指标不同小写字母表示差异显著(P<0.05)。Fig.2 Leaf tissue structure indices of six orchids in Beipan River basin of GuizhouDifferent lowercase letters indicate significant differences (P<0.05).
梳帽卷瓣兰叶片厚度、叶肉厚度和主叶脉厚度最大,分别为(1164.05±104.49) μm、(1128.45±146.41) μm 和(940.51±70.31) μm,主脉维管束直径和主脉凸起度最小,分别仅为(103.10±2.03)μm 和(81.78±12.80) μm,较于单叶石仙桃和栗鳞贝母兰指标差异较为显著。
2.1.2 假鳞茎结构
假鳞茎主要由表皮、基本组织和维管束3部分组成(图3)。表皮细胞体积较小,由1层排列紧密的方形或圆形细胞组成,栗鳞贝母兰表皮最厚,为(23.94±2.94) μm,兔耳兰表皮最薄,为(13.54±1.39) μm。基本组织细胞形态各异,呈圆形、多边形或无规则等。地生兰基本组织细胞均较小且数量多,附生兰基本组织细胞均较大且数量相对较少。维管束呈圆形,半附生兰维管束呈“环状”或“帽状”散生其中,发育程度不同、大小不一、数量差异明显。假鳞茎的维管束均表现出中心发育较成熟,直径较大,边缘逐渐变小且无维管束分布。丘北冬蕙兰假鳞茎横截面积、直径、维管束均为最大,分别为(76.93±0.17) mm2、(9 930.59±343.22) μm、(431.98±95.49) μm,兔耳兰假鳞茎横截面积、直径最小,分别为(9.85±0.02) mm2、(3671.8±128.63) μm。维管束分布数量依次排序为单叶石仙桃(183)>莎叶兰(136)>丘北冬蕙兰(134)>兔耳兰(70)>栗鳞贝母兰(38)>梳帽卷瓣兰(32)。地生兰假鳞茎结构整体表现出较大的维管束,高效输导特征,附生兰基本组织细胞较大,具有较强的储水结构优势。

图3 贵州北盘江6种兰科植物假鳞茎的结构横切面A.丘北冬惠兰;B.莎叶兰;C.兔耳兰;D.单叶石仙桃;E.栗鳞贝母兰;F.梳帽卷瓣兰;ep.表皮;gt.组织细胞;vb.维管束。Fig.3 Cross section of pseudobulb of six orchids in the Beipan River of Guizhou ProvinceA.C.qiubeiense.B.C.cyperifolium.C.C.lancifolium.D.P.leveilleana.E.C.flaccida.F.B.andersonii.ep,epidermis.gt,histocyte.vb,vascular bundle.
2.1.3 根结构
兰科植物根部结构由根被、皮层、维管束3部分构成(图4),根直径和皮层有较大差异,丘北冬蕙兰根直径最大,为(4095.03±208.12) μm,其次为兔耳兰和莎叶兰,直径分别为(3678.66±56.50) μm 和(3361.59±435.45) μm,远大于附生兰。根皮层结构由外皮层和内皮层之间的组织细胞构成,呈圆形、长椭圆形、近方形或不规则形。地生兰和半附生兰根皮层为10~13层细胞组成,相较于附生兰,细胞较小、数量较多,其中丘北冬蕙兰皮层最厚,为(1870.89±946.54) μm。

图4 贵州北盘江6种兰科植物根表皮(A-F)、皮层(G-L)和维管束(M-R)的结构横切面图A、G、M.丘北冬惠兰;B、H、N.莎叶兰;C、I、O.兔耳兰;D、J、P.单叶石仙桃;E、K、Q.栗鳞贝母兰;F、L、R.梳帽卷瓣兰;rh.根被;h.根毛;ex.外皮层;co.皮层;ed.内皮层;vbs.维管束鞘;vb.维管束;pe.中柱鞘;ph.韧皮部;x.木质部;p.髓。Fig.4 Cross section of the roots (A_F),cortex (G_L) and bundle (M_R) of six orchids in the Beipan River of Guizhou ProvinceA,G,M.C.q iubeiense.B,H,N.C.c yperifolium.C,I,O.C.l ancifolium.D,J,P.P.l eveilleana.E,K,Q.C.flaccida.F,L,R.B.a ndersonii.rh,velamem.h,root hair.ex,exodermis.co,cortex.ed,endodermis.vbs,vascular bundle sheath.vb,vascular bundle.pe,pericycle.ph,phloem.x,xylem.p,pith.
单叶石仙桃、栗鳞贝母兰、梳帽卷瓣兰3种附生兰皮层为5~7层圆形细胞组成,细胞大、数量较少,有细胞间隙,厚度分别为(399.71±149.38) μm、(306.01±138.91) μm 和(293.28±133.31) μm,根直径和皮层显著小于地生兰和半附生兰。其中梳帽卷瓣兰皮层厚度最小,但与根直径比值较大(图5)。根被厚度和维管束直径差异不明显,兔耳兰维管束直径最大,为(1099.7±6.94) μm,梳帽卷瓣兰最小,为(300.6±11.19) μm。根被结构特征差异较明显,地生兰和半附生兰特征趋于相似,均由5~9层长方形、五边形或六边形薄壁细胞紧密嵌合排列组成,覆盖在根的最外层,无细胞间隙。丘北冬蕙兰根被厚度最大,为(558.27±60.94) μm,是附生兰根被厚度的2~7倍。附生兰根被为单层或2~5层椭圆形、长方形或圆形薄壁细胞组成,梳帽卷瓣兰由单层长方形细胞紧密排列而成,栗鳞贝母兰根被细胞外壁向外凸起形成根毛结构。

图5 植物根部结构指标Fig.5 Structural indices of plant roots
2.2 植物叶片、假鳞茎、根结构的生态适应性
对6种兰科植物的营养器官22个结构指标进行Pearson相关性分析,结果(图6)表明:各营养器官结构之间存在显著相关关系,叶片结构的TL 与LT、DVB1、MT、TV 呈极显著正相关(P<0.01),MT 和VPD、PVB1 呈极显著正相关,与TLC、DVB2呈显著负相关(P<0.05),TV 与VPD、PVB1、DS、ET 相关性较弱;假鳞茎的DS 和ET、NVB、CSAS呈极显著正相关,PVB2与DVB2呈极显著正相关关系外,与DS、ET、NVB、CSAS均呈极显著负相关,与DVB2呈极显著正相关,与VBD 呈显著正相关;根结构的RD、RCT 和CT 与PVB3呈极显著负相关,CT 和DVB3相关性不明显。
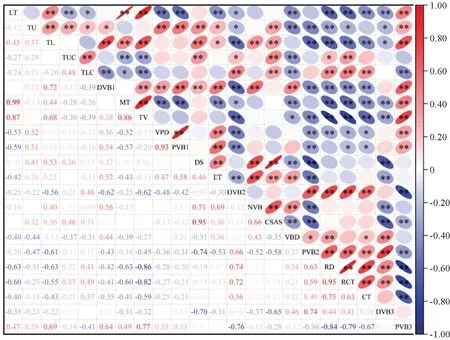
图6 植物结构指标相关关系*表示相关性显著(P<0.05),**表示相关性极显著(P<0.01);LT.叶片厚度;TU.上表皮厚度;TL.下表皮厚度;TUC.上角质层厚度;TLC.下角质层厚度;DVB1.叶维管束直径;MT.叶肉厚度;TV.叶脉厚度;VPD.叶脉凸起度;PVB1.叶维管束直径占比;DS.假鳞茎直径;ET.表皮厚度;DVB2.假鳞茎维管束直径;NVB.假鳞茎维管束数量;CSAS.假鳞茎横截面积;VBD.假鳞茎维管束密度;PVB2.假鳞茎维管束直径占比;RD.根直径;RCT.根被厚度;CT.皮层厚度;DVB3.根维管束直径;PVB3.根维管束直径占比。下同。Fig.6 Correlation of plant structural indices* indicates significant correlation (P<0.05),** indicates extremely significant correlation (P<0.01).LT,leaf thickness.TU,thickness of upper epidermis.TL,thickness of lower epidermis.TUC,thickness of upper cuticle.TLC,thickness of lower cuticle.DVB1,the vascular bundle diameter of the blade.MT,mesophyll thickness.TV,vein thickness.VPD,vein protu-berance degree.PVB1,percentage of the diameter of the vascular bundle of the blade.DS,diameter of pseudobulb.ET,epidermal thickness.DVB2,diameter of vascular bundles of pseudobulb.NVB,number of vascular bundles in pseudobulb.CSAS,cross-sectional area of pseudobulb.VBD,density of vascular bundles in pseudobulb.PVB2,the fraction of the diameter of the vascular bundle of pseudobulb.RD,root diameter.RCT,root coat thickness.CT,cortical thickness.DVB3,the diameter of the vascular bundle of the root.PVB3,the fraction of the diameter of the vascular bundle of the root.The same as below.
整体上叶片持水结构(TU、TL、TUC)与假鳞茎的储水功能结构(DS、CSAS)呈极显著正相关,两者在功能上具有相似性,彼此间存在相互促进协调关系,与PVB2呈极显著负相关;叶片的持水结构与机械支撑结构(LT、TL、DVB1、MT、TV)与根对物质吸收功能(RD、RCT、CT)呈极显著负相关,与根传导结构(PVB3)呈极显著正相关;假鳞茎的输送结构(DVB2、PVB2)与根的对物质吸收功能性状(RD、RCT、CT)间存在极显著正相关,与输送结构(PVB3)呈极显著负相关关系。
通过相关性系数去除数据间共线性较大的指标(绝对值大于0.8),对筛选的结构参数进行主成分分析(PCA),可以将所有指标概括为2个具有代表性的主成分(图7)。PC1和PC2中共解释了植物结构参数变异的33%和20.6%的总方差,PC1主要以TL、DVB1、PVB2载荷较大,分别为82.89%、78.82%和75.62%,这些指标主要是植物叶片的物质传输与机械支撑功能相关因子。PC2 主要以CSAS、TUC、DVB3载荷较大,分别为89.86%、59.18%和59.1%,主要为叶片水分保持与假鳞茎的营养物质储存运输的相关因子。
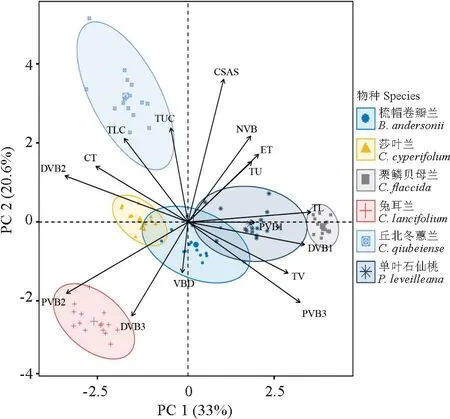
图7 贵州北盘江6种兰科植物根、假鳞茎、叶结构参数主成分分析Fig.7 Principal component analysis of structural parameters of rhizomes and leaves of six orchids in the Beipan River of Guizhou Province
3 讨论
3.1 兰科植物适应环境的结构特征
喀斯特地区生境“干旱”是限制植物生长和繁殖的主要因素之一,环境因素长期影响着植物各器官的发育与演化,因此喀斯特地区植物大多向着保水和提高物质利用效率的结构方向发生适应性变化[21]。本研究对比不同生活型兰科植物结构发现,地生兰(丘北冬蕙兰)与半附生兰(兰兔耳兰、莎叶兰)具有相似的根和假鳞茎结构特征。即肉质根系较粗壮,根被和皮层由多层细小紧密的细胞组成,对深厚土层具有更大的穿透力和生长势能[22]。根系越粗大越能支撑更多“土壤-根际”真菌的生存繁殖,对兰科植物生存生长具有重要的促进意义[23]。Rieger等[24]研究发现,根直径、根皮和根被结构与水分传导速率呈显著负相关,该结构越厚或越紧密越能增加土壤水分横跨进入维管束的距离和阻力,为较缓慢的吸水结构特征。该结构特征是适应水分充足湿润环境的表现,与地生兰和半附生兰温润林下生存环境相一致。但不同的是,丘北冬蕙兰与附生兰(单叶石仙桃、栗鳞贝母兰和梳帽卷瓣兰)具有相似抗旱特征的叶和假鳞茎结构,即较厚的叶片、叶肉细胞和上角质层等阳生叶结构特征和发达的维管束。相关研究表明,表皮和角质层结构能有效反射强光照射、降低叶片温度、减少叶肉水分蒸腾丧失[25-26]。发达的根部维管束和假鳞茎能为植物提供强有力的机械支撑外,还能提高营养物质和水分的输送速率,及时满足植物光合生长和蒸腾作用等对水分的需求,是植物适应强光、高温和干旱环境的结构特征。与丘北冬蕙兰不同的是,单叶石仙桃、栗鳞贝母兰和梳帽卷瓣兰的根系相对细小,较薄的皮层和单细胞根被结构大大减少水分穿越根被和皮层进入维管束向上运输的距离和阻力,能够更高效地获取养分和水资源;而细小的根系能扩大根系对土层探索吸收水分和养分的空间,对环境变化做出迅速的调整反应,具有适应石壁、石缝、树干和腐殖质层等特殊附生生境的结构特征[27]。半附生兰(莎叶兰、兔耳兰)叶片表皮和角质层均较薄,不利于对光的反射与水分的保持,根结构对水分的吸收速率也较低。但根、假鳞茎和叶的维管组织比较发达,对植物起到强有力的机械支撑,避免植物在强光高温环境下受到萎焉损伤。通过高速运输物质和水分,满足植物生长发育过程中对水分的需求,这样的适应结构可能和半附生兰所面临的干旱胁迫强度较弱有关。
尽管相同生活型物种器官间存在趋同适应的结构特征,但局部生境差异长期影响仍然会促使植物内部结构差异化,从而选择进化出不同的资源获取方式[28-29]。本研究对比相同生活型兰科植物结构发现,附生兰(单叶石仙桃、梳帽卷瓣兰和栗鳞贝母兰)中,栗鳞贝母兰和梳帽卷瓣兰具有栅栏组织和海绵组织的分化,可通过栅栏组织调节叶片对光的捕获能力,进一步促进光合作用,是植物增强光合作用和减少水分散失适应喀斯特干旱生境的有效结构[30]。随着植株增大,维管束提高水分和养分的传输效率,并储存于发达的假鳞茎结构中,满足植物干旱时期对水分的需要。而单叶石仙桃叶片性状则与之相反,可能与其长期干旱或间接性降雨的环境条件有关[31]。栗鳞贝母兰叶片和表皮较厚,但角质层较薄,叶片蒸腾作用大,水分散失快,能适应强光、高温和干旱的喀斯特生境,可能与其发达的假鳞茎储水功能结构密切相关。三者的假鳞茎均具有发达的储水结构,为植物补充光合生长过程中散失的水分和营养物质,弥补和协调叶片结构保水性和抗阳性较差的弱点。与朱晓旭等[32]通过对不同生活型物种功能性状特征对环境变化的响应研究结果相似,不同生活型物种适应环境变化的过程中,性状之间相互协调表现出共性结构时,也会在适应环境过程中发生差异性变化。植物结构的差异化是适应环境改变资源获取方式的能力,往往决定植物生存竞争的成功与否,是植物适应环境最直接的策略之一[33]。附生兰假鳞茎中散生的维管束数量和大小虽不呈规律性,但维管束占假鳞茎横截面积的比例均较小。说明附生兰假鳞茎为植物储存水分和养分过程中有严格的占比控制,在维管束大小和数量之间做出生长策略权衡[34],协调其他器官缺陷,满足兰科植物在干旱缺水环境中对水分和养分物质的需求。
3.2 兰科植物结构的生态策略
基于营养器官结构特征对比不同生活型植物间结构差异是揭示植物适应策略的重要方法之一,不同环境条件下,植物对环境的响应不仅仅通过单一器官的结构性状来实现,而是多器官间相互协调,在生理结构和形态上做出调整以此获得适应特殊生境的能力[35]。植物的部分结构性状可能来源于长期进化的遗传基因所决定的稳定性状,受进化时间和自然选择影响较小;而部分结构发生变化主要取决于对局部环境差异的响应[36]。本研究兰科植物在生活型的分类水平上,根部结构比叶片和假鳞茎更具稳定性和趋同适应特征,叶片与假鳞茎结构之间存在密切地协调作用,兰科植物主要通过叶片和假鳞茎结构相互协调促进植物适应局部环境差异。叶片是植物感应外界环境最直接的营养器官,具有较高的可塑性,是植物进行光合作用、蒸腾作用、生产物质的重要场所[37]。吴春梅等[14]对假鳞茎结构的研究表明,假鳞茎作为兰科植物特殊的贮藏器官,不仅具有储存水分和养分的功能,还是碳水化合物的重要补充来源。当叶片的光合作用效率低时,假鳞茎可以储存大量的碳水化合物,为叶片提供能量[38]。在长期干旱的环境下,假鳞茎中储存的水溶性多糖可以被调动并释放,将水分和养分物质转移到其他器官,从而减轻植物叶片含水量的降低和水势下降的速率,帮助植物适应干旱的环境胁迫[39]。假鳞茎和叶片同样具有光合作用和温度调节等相似的生理生态功能,赋予植物在恶劣生境中生存和生长的能力,是协调植物与环境之间相互关系的重要器官,成为兰科植物适应喀斯特干旱生境的关键所在,这一观点在多项研究中得到验证[40-43]。不同兰科植物结构之间呈“此消彼长”的协调规律,与倪榕蔚等[44]的研究结果一致,植物增大某一功能性状上的投入必将减少其他性状的生长资源配比,营养器官之间相互协调共同适应环境。莎叶兰和兔耳兰在根和假鳞茎的结构上表现出较差的吸收与储水能力,同时增强叶片的光合作用、保水能力和养分运输结构,植物对资源的利用方式主要为叶片与环境资源间的相互作用,属于消费型“快投资—收益”策略,协调了根和假鳞茎结构上的弱点。丘北冬蕙兰假鳞茎结构具有较强的储水和运输能力,相较于莎叶兰和兔耳兰更具有抗旱的优势,与附生兰营养器官在功能权衡上具有相似特征。即主要将生长资源投入在高密度组织结构上,促进植物叶、根的养分吸收与传输,加强假鳞茎对资源的储备功能,为资源保守型“慢投资—收益”策略,假鳞茎发达的储水组织是喀斯特地区兰科植物应对季节性干旱和临时性降雨生境高度适应的特殊结构。
4 结论
北盘江兰科植物器官结构特征与植物适应喀斯特干旱环境的策略有密切联系。通过对比不同生活型兰科植物叶片、假鳞茎、根结构特征发现:
(1)兰科植物结构之间存在密切相关关系,具相似功能的结构在不同物种及不同器官间表现出明显协同关系,不同功能结构间具有明显的权衡关系,即“此消彼长”的协调规律。
(2)不同生活型间结构差异主要体现在根对资源获取功能性状上,附生兰植物根结构相较于地生兰和半附生兰更具有获取生存资源能力的优势;物种间叶片和假鳞茎结构特征不呈明显的差异规律,植物主要通过调节叶片和假鳞茎结构功能适应生境。
(3)莎叶兰和兔耳兰结构具有更发达的叶功能结构,更倾向于快速获取资源型的“快投资—收益”策略;丘北冬蕙兰和3种附生兰具有较强的抗旱、储水和输导结构等高密度组织结构特征,更倾向于资源保守型“慢投资—收益”策略。
