不同种源花榈木种子表型性状变异分析
2024-03-06李天翔肖亚琴吴琴香吴小丽曹基武
李天翔,肖亚琴,杨 骏,吴琴香,吴小丽,曹基武
(1.中南林业科技大学,湖南 长沙 410004; 2.湖南省林业科学院,湖南 长沙 410004)
花榈木(OrmosiahenryiPrain)又名红豆树、臭桶柴、花梨木等,为豆科(Leguminosae)蝶形花亚科(Papilionoideae)红豆属(Ormosia)常绿乔木,是国家二级保护野生植物[1]。花榈木木材质地坚硬,花纹美观,是制作家具的上等材料[2]。花榈木适应性较强,在酸性、中性土壤中均能正常生长[3]。其根有固氮菌[4],能改良土壤、增加土壤肥力[5];萌芽力强,耐高温日灼,砍伐或火烧后仍可萌发。同时,花榈木树形优美,树干笔直,树冠浓荫,四季常绿,花色淡雅,荚果吐红,是“材貌双全”的园林绿化树种和防火树种[6-9]。此外,花榈木还具有重要的药用研究价值:花榈木的根、茎及叶可入药,主治跌打损伤、腰肌劳损、风湿关节痛等;现代医学研究表明,花榈木提取物在治疗生理和心理疾病上具有巨大的应用价值和潜力[10];叶片中含有黄酮类化合物,可以有效抑制抑郁行为,是具有强抗抑郁作用的天然潜在候选物[11-12];根皮中分离得到2种化合物,具有一定的抗氧化和抗癌活性[13-14]。
种子性状是遗传物质和环境因子共同作用的结果,而丰富的种源变异是林木优良种源筛选的必要条件[15-18]。目前关于花榈木的研究主要集中于种子休眠[19-20]、苗木繁育[21-24]、生理胁迫[25-27]及人工造林[28-29]等方面,而对于花榈木种源遗传变异的研究较少。本研究对花榈木8个种源种子的长度、宽度、千粒重、种形指数及其生长的地理环境因子进行关联分析,研究4个表型性状的遗传变异情况,以期为花榈木种质资源的保护提供参考,为提高花榈木的苗木生产及其适应能力提供理论依据,为优良种源筛选奠定理论基础。
1 材料与方法
1.1 试验材料
2021年10月,花榈木种子成熟时,分别在浙江省的杭州市、宁波市和湖南省的长沙市、湘潭市、株洲市、邵阳市采种。选取群落中长势健壮的母株采集种实,将种实去除果荚及杂质,晾干后置于4℃冰箱内冷藏备用。种源地的地理坐标和主要气候因子见表1。
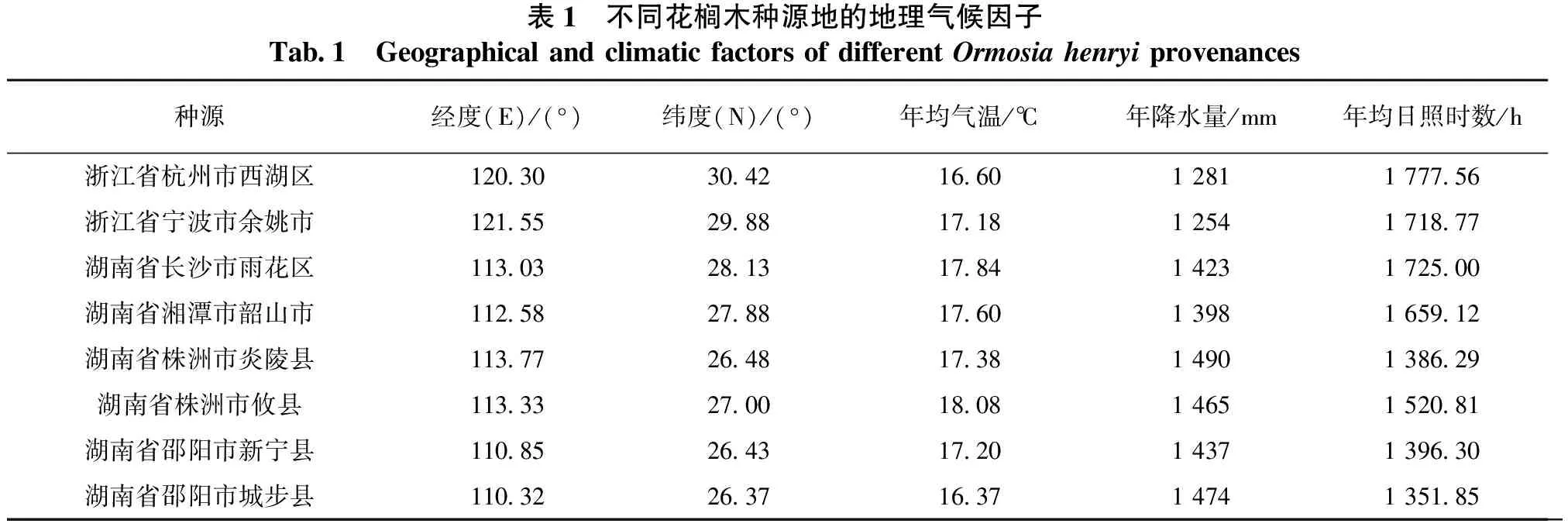
表1 不同花榈木种源地的地理气候因子Tab.1 Geographical and climatic factors of different Ormosia henryi provenances种源经度(E)/(°)纬度(N)/(°)年均气温/℃年降水量/mm年均日照时数/h浙江省杭州市西湖区120.3030.4216.601 2811 777.56浙江省宁波市余姚市121.5529.8817.181 2541 718.77湖南省长沙市雨花区113.0328.1317.841 4231 725.00湖南省湘潭市韶山市112.5827.8817.601 3981 659.12湖南省株洲市炎陵县113.7726.4817.381 4901 386.29湖南省株洲市攸县113.3327.0018.08 1 4651 520.81湖南省邵阳市新宁县110.8526.4317.201 4371 396.30湖南省邵阳市城步县110.3226.3716.371 4741 351.85
1.2 测量方法
每个花榈木种源采用“四分法”随机选取种子30粒,用电子游标卡尺分别测量每组种子的长度和宽度(精确度0.01mm),重复3次。千粒重测量参考《林木种子检验规程》(GB/T 2772—1999)[30]。分别采用以下公式计算种子种形指数[31-33]、广义遗传率[34]:
种形指数=种子长度/种子宽度
(1)
广义遗传率=1-1/F
(2)
式(2)中:F为方差分析中的F检验值。
1.3 数据处理
采用Excel 2019软件处理原始数据,采用SPSS 25.0软件进行数据方差分析、相关分析和聚类分析。采用Origin 2021软件作图。
2 结果与分析
2.1 不同种源花榈木种子表型性状差异分析
对不同种源花榈木种子表型性状进行分析可知,8个种源花榈木种子的长度、宽度、千粒重和种形指数均在种源间存在极显著差异(P<0.01)(见表2)。8个种源的种子长度平均值为9.39mm,变化范围为8.67~10.53mm,其中韶山市种源的种子长度最大(10.53mm),城步县种源的种子长度最小(8.67mm)。8个种源的种子宽度平均值为7.58mm,变化范围为6.91~8.56mm,其中雨花区种源的种子宽度最大(8.56mm),炎陵县种源的种子宽度次之(8.23mm),新宁县种源的种子宽度最小(6.91mm,为雨花区种源的81%);雨花区种源种子的千粒重最大(277.10g),约为新宁县种源种子千粒重(197.50g)的1.4倍。雨花区种源的种子种形指数为1.08,种子外形为扁圆形;新宁县种源的种子种形指数为1.39,种子为长椭圆形。
8个种源花榈木种子性状变异系数为6.78%~10.24%。种子千粒重的变化范围为197.50~277.10g,变异系数最大(10.24%);种子宽度和种形指数的变化范围分别为6.91~8.56mm、1.08~1.39,他们的变异系数次之,分别为8.01%和7.99%;种子长度的变异系数最小,为6.78%。研究结果表明不同种源花榈木种子表型性状存在丰富的变异,且由广义遗传率可知,种子长度、宽度和千粒重的遗传率分别为0.955、0.957和0.996,说明花榈木的表型性状受到极强的遗传控制,不同种源的花榈木种群间种子表型性状遗传率稳定。
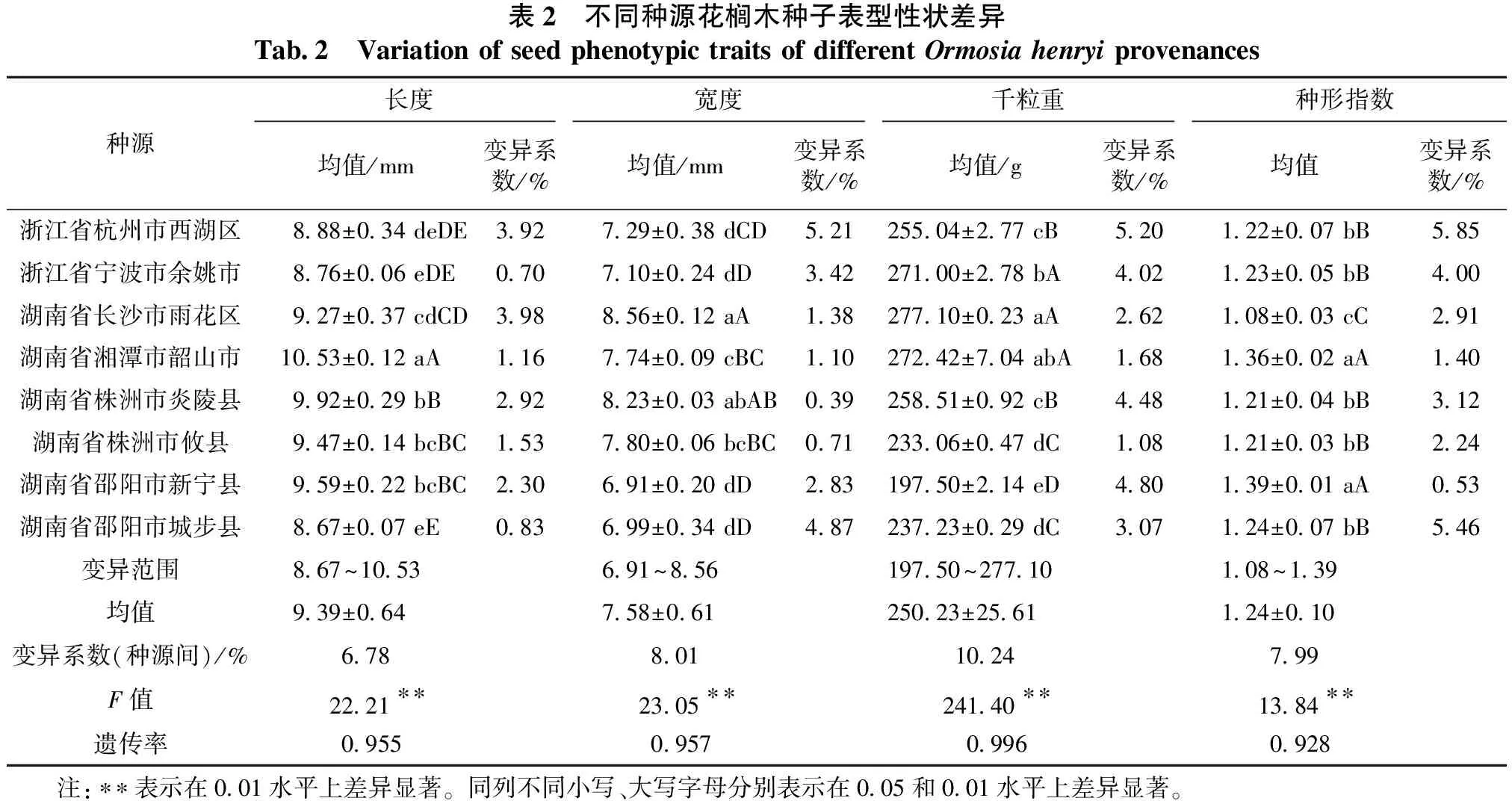
表2 不同种源花榈木种子表型性状差异Tab.2 Variation of seed phenotypic traits of different Ormosia henryi provenances种源长度宽度千粒重种形指数均值/mm变异系数/%均值/mm变异系数/%均值/g变异系数/%均值变异系数/%浙江省杭州市西湖区8.88±0.34 deDE3.92 7.29±0.38 dCD5.21255.04±2.77 cB5.20 1.22±0.07 bB5.85浙江省宁波市余姚市8.76±0.06 eDE0.70 7.10±0.24 dD3.42271.00±2.78 bA4.02 1.23±0.05 bB4.00湖南省长沙市雨花区9.27±0.37 cdCD3.98 8.56±0.12 aA1.38277.10±0.23 aA2.62 1.08±0.03 cC2.91湖南省湘潭市韶山市10.53±0.12 aA1.16 7.74±0.09 cBC1.10 272.42±7.04 abA1.68 1.36±0.02 aA1.40湖南省株洲市炎陵县9.92±0.29 bB2.92 8.23±0.03 abAB0.39258.51±0.92 cB4.48 1.21±0.04 bB3.12湖南省株洲市攸县9.47±0.14 bcBC1.53 7.80±0.06 bcBC0.71233.06±0.47 dC1.08 1.21±0.03 bB2.24湖南省邵阳市新宁县9.59±0.22 bcBC2.30 6.91±0.20 dD2.83197.50±2.14 eD4.80 1.39±0.01 aA0.53湖南省邵阳市城步县8.67±0.07 eE0.83 6.99±0.34 dD4.87237.23±0.29 dC3.07 1.24±0.07 bB5.46变异范围8.67~10.536.91~8.56197.50~277.101.08~1.39均值9.39±0.647.58±0.61250.23±25.61 1.24±0.10变异系数(种源间)/%6.788.0110.247.99F值22.21∗∗23.05∗∗241.40∗∗13.84∗∗遗传率0.9550.9570.9960.928 注:∗∗表示在0.01水平上差异显著。同列不同小写、大写字母分别表示在0.05和0.01水平上差异显著。
2.2 花榈木种子表型性状间的相关性分析
花榈木种子各表型性状之间的相关性分析见表3。由表3可知,花榈木种子宽度和千粒重与种形指数之间有较弱的负相关关系,相关系数分别为-0.622和-0.534;千粒重越大,种形指数越小,表明花榈木种子有从椭圆形到圆形变异的趋势。另外,种子宽度与千粒重有较弱的正相关性,相关系数为0.558,但相关性未达到显著水平。花榈木的种子长度、宽度及千粒重之间并无显著相关性。
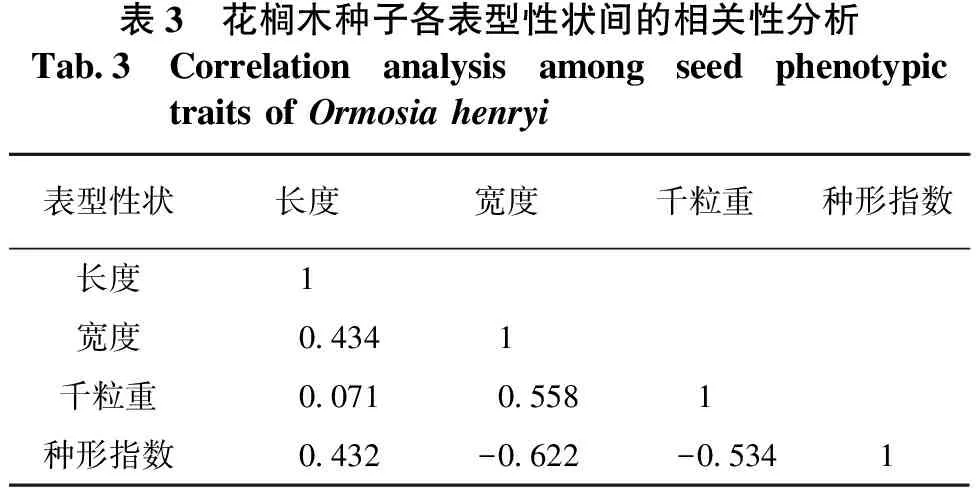
表3 花榈木种子各表型性状间的相关性分析Tab.3Correlation analysis among seed phenotypic traits of Ormosia henryi表型性状长度宽度千粒重种形指数长度1宽度0.4341千粒重0.0710.5581种形指数0.432-0.622-0.5341
2.3 花榈木种子表型性状与地理气候因子的相关性分析
花榈木种子表型性状与种源采集地的地理气候因子的相关分析见表4。相关性分析发现,种子宽度与年均气温成显著正相关,相关系数为0.679;种子千粒重与年均日照时数成显著正相关,相关系数为0.665;种子长度与经度、纬度均成负相关关系,相关系数分别为-0.411,-0.370,但相关性未达到显著水平。
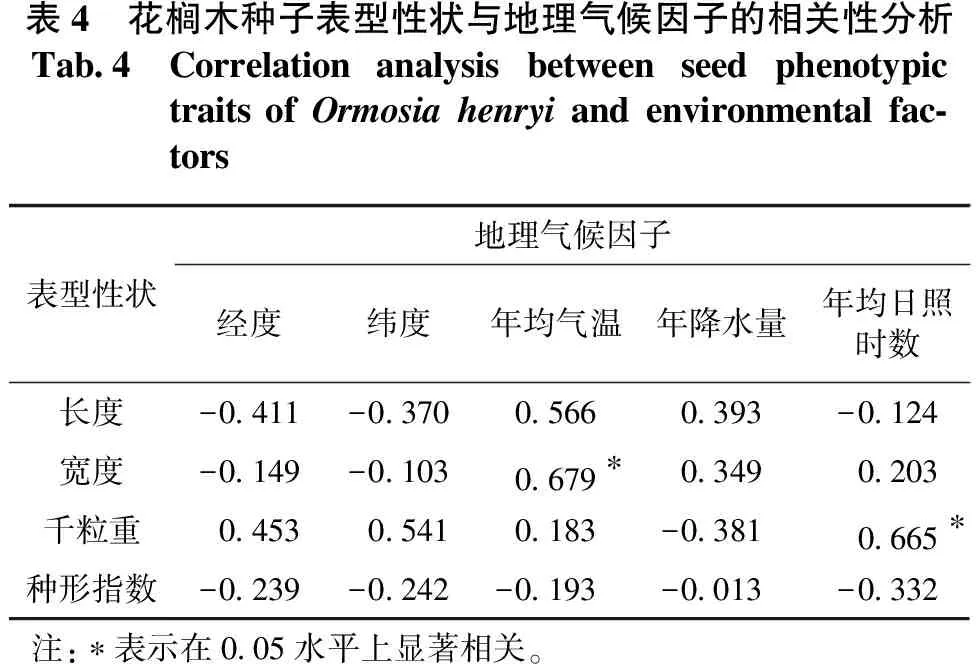
表4 花榈木种子表型性状与地理气候因子的相关性分析Tab.4Correlation analysis between seed phenotypic traits of Ormosia henryi and environmental fac-tors表型性状地理气候因子经度纬度年均气温年降水量年均日照时数长度-0.411-0.3700.5660.393-0.124宽度-0.149-0.1030.679∗0.3490.203千粒重0.4530.5410.183-0.3810.665∗种形指数-0.239-0.242-0.193-0.013-0.332注:∗表示在0.05水平上显著相关。
2.4 花榈木种源的聚类分析
对8个种源花榈木种子表型性状进行系统聚类分析,以欧式距离10为阈值时,可将8个花榈木种源划分为5个类群(见图1):类群Ⅰ包括三个种源,分别是余姚市、城步县和西湖区种源,类群Ⅲ为炎陵县和攸县种源,类群Ⅱ、Ⅳ、Ⅴ分别是新宁县、雨花区和韶山市种源。从地理位置来看,同来源于浙江省的杭州西湖区种源和宁波余姚市种源被划分为同一类群,同来源于湖南株洲的炎陵县种源和攸县种源也被划分为同一类群,表明花榈木种子的表型变异具有区域板块性;而同来源于湖南邵阳的新宁县种源和城步县种源被划分为两个亲缘关系较远的类群,说明种子表型变异还具有随机性。综上可知,由于地理隔离、自然选择与基因漂流等多因素的影响,花榈木种子表型性状具有地域变异性和随机变异性。

图1 不同种源花榈木聚类分析Fig.1 Cluster analysis of different provenances of Ormosia henryi
3 结论与讨论
花榈木种子表型性状具有丰富的遗传多样性,且表型性状具有稳定的遗传率,种子长度、宽度、千粒重和种形指数对应的遗传率分别为0.955、0.957、0.996、0.928。花榈木种子平均长度为9.39mm,平均宽度为7.58mm,平均千粒重为250.23g,其中长沙市雨花区种源的种子宽(8.56mm)和千粒重(277.10g)最大,在种源选择时可优先考虑。花榈木种子的发育与种源地的气候因子息息相关,种子宽度与种源地的年均气温有显著正相关关系,种子千粒重随着日照时长的增长有不断增大的趋势。聚类分析中花榈木种源可分为5个类群,由不同类群种源的地理位置可知,种子表型性状的变异具有地域性和随机性。
种质资源是物种遗传改良的重要物质基础,是新品种选育和品质创新的坚实后盾[35]。种子表型性状是种子生产、苗木繁育等植物生命周期活动中的关键指标[36],其变异系数和多样性指数反映了种质间性状的离散程度和多样性。对植物表型性状遗传多样性进行分析,是合理利用植物种质资源和创制新种质的前提和基础[37]。花榈木不同种源的种子在长度、宽度、千粒重等方面存在极显著差异,各性状变化范围分别为8.67~10.53mm、6.91~8.56mm、197.50~277.10g;其中千粒重的变异系数最大,为10.24%,表明花榈木的种质资源具有较大的遗传变异性,与天山花楸(Sorbustianschanica)[31]、栝楼(Trichosantheskirilowii)[38]、文冠果(Xanthocerassorbifolia)[33]种子的研究一致。本研究中花榈木种子表型遗传率比较稳定,遗传率均在0.92以上。黄振等[39]研究杉木(Cunninghamialanceolata)遗传率时发现,球果表型性状的遗传率也稳定在0.9以上,但幼苗的株高、地径的遗传率显著下降,种子的遗传率和幼苗的遗传率偏差较大,因为种子采自不同地理环境和气候条件,人工创造同一个环境下进行苗木繁育,气候环境的干扰可能会使幼苗的生长产生较大的差异,因此仅靠种子表型性状的研究,还不足以确定优良的种质资源。因此,下一步考虑对不同种源花榈木幼苗生长的遗传多样性进行研究,综合种子和幼苗的遗传率筛选出更加优质的种质资源。
同一树种在不同地理环境和气候因子的条件下,种子表型性状会发生一定程度的变异,例如:蒙古栎(Quercusmongolica)种子的宽度与海拔成反比[32];四川牡丹(Paeoniadecomposita)种子的千粒重随纬度的增大而增大[40];防风(Saposhnikoviadivaricata)种子的千粒重与经度存在显著正相关关系[41];降香黄檀(Dalbergiaodorifera)种子的千粒重与海拔呈显著正相关[42]。在相关性分析中,花榈木种子千粒重与年均日照时数存在显著正相关关系。千粒重反映了种子的大小和饱满程度,花榈木属于温带阔叶树种,日照时间的增长有利于植株营养物质的积累,从而促进种子的发育。
通过聚类分析,将花榈木8个种源划分为5个类群,其中浙江杭州西湖区种源和宁波余姚市种源被归为同一个类群,株洲市炎陵县种源和攸县种源也被归为同一个类群,但同属邵阳市的城步县种源和新宁县种源却被划分到两个不同的类群,且亲缘关系最远,表明花榈木种子表型性状的变异具有地域性、随机性,这与王旭军等[34]对红榉(Zelkovaschneideriana)种子的研究结果相似。因此,综合种子的地理变异规律和遗传分化的差异,对于林木优良种质资源的筛选具有重大指导意义。
