玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡生产性能、肝脏糖代谢、内源消化酶活性和肠道中葡萄糖转运载体基因表达的影响
2024-03-05常丹萍刘观忠杨博文赵国先张东远冯志华张海华
常丹萍 刘观忠 杨博文 赵国先 张东远 冯志华* 李 茜 张海华 王 珏
(1.河北农业大学动物科技学院,保定 071000;2.中科康源(唐山)生物技术有限公司,迁安 064400;3.河北省畜牧兽医研究所,保定 071000;4.河北科技师范学院,秦皇岛 066000)
近年来,畜禽业发展迅速造成的饲料资源短缺问题日益严重。玉米-豆粕型饲粮是我国的主导饲粮结构,这种结构对豆粕年消耗量巨大。我国大豆年产量约1 640万t[1],国内大豆严重供给不足导致我国大豆对外依存度超80%[2],2022年末豆粕价格同比年初增长39.21%[3],造成饲料成本大幅提高。解决上述问题,有2个途径,一是开发新的饲料资源,二是提高现有饲料养分利用率。
目前很多饲料企业常会使用一部分菜籽粕、棉籽粕等杂粕代替豆粕或鱼粉等常规蛋白质饲料,这样不仅能缓解饲料资源短缺的问题,还可以降低饲料成本。我国具有丰富的杂粕资源,各种农副产品如棉籽饼粕、菜籽饼粕、花生饼粕等产量巨大且价格低廉,2021年棉籽粕、菜籽粕、花生粕的年产量分别为580万、700万、400万t[4],但是杂粕的应用在实际生产中并不广泛,其主要原因有杂粕中氨基酸不平衡、粗纤维与可溶性非淀粉多糖含量高等。杂粕中除了花生粕、芝麻粕的粗纤维含量较低外,其余的粗纤维含量都较高,均在12%左右[5],而粗纤维是植物细胞壁的主要成分,其含量高会影响动物对饲粮中养分的利用。杂粕中的非淀粉多糖主要有纤维素、果胶、阿拉伯木聚糖、木聚糖、β-葡聚糖等,这些物质是细胞壁的主要组成成分[6],单胃动物尤其是鸡自身不能分泌消化这些物质的酶,使得细胞壁无法充分打开,导致饲料养分利用率低[7]。因此,破坏细胞壁是提高饲料养分利用率的有效手段。破坏细胞壁的方法有很多,目前研究较为热门且效果显著的就是添加外源性酶制剂。由于细胞壁的结构很复杂,单一的酶对细胞壁的破坏效果并不理想,需要一系列酶协同作用才能破坏。研究表明,在蛋鸡饲粮中添加复合酶制剂可以显著改善其生产性能、蛋品质及养分利用率等相关指标[8-9]。由羧甲基纤维素酶、β-葡萄糖苷酶、木聚糖酶、外切葡聚糖酶等一系列酶复合而成的复合纤维素酶,针对细胞壁结构设计,与其他酶制剂相比所含酶种类和配合比例理论上更加科学。但该复合纤维素酶在蛋鸡杂粕型饲粮中的应用效果及其作用机理尚未见报道。为此,本试验通过在玉米-杂粕型饲粮中添加不同水平的复合纤维素酶,研究其对蛋鸡生产性能、蛋品质、肝脏中糖代谢相关酶活性、内源消化酶活性以及肠道中葡萄糖转运载体基因表达的影响,并探讨其作用机理,明确蛋鸡杂型饲粮中复合纤维素酶的应用效果并筛选出适宜添加量,为复合纤维素酶在蛋鸡杂粕型饲粮中应用提供试验基础和数据支撑。
1 材料与方法
1.1 试验材料
复合纤维素酶是基于里氏木霉与黑霉酶系的高效协同作用通过混菌发酵与高通量筛选技术获得,其组成与酶活性如下:羧甲基纤维素酶活性20 389.6 U/g、木聚糖酶活性4 054.4 U/g、外切葡聚糖酶活性3 284 U/g、β-葡萄糖苷酶活性10 528 U/g、淀粉酶活性56 U/g、蛋白酶活性28 U/g。
1.2 试验设计
采用单因素试验设计,选取288只健康状况良好的53周龄海兰褐蛋鸡,随机分为4组,每组6个重复,每个重复12只鸡。各组平均体重、产蛋率无显著差异(P>0.05),按照常规饲养方式进行笼内饲养。4组蛋鸡分别饲喂添加0(对照组)、200、400、600 mg/kg复合纤维素酶的玉米-杂粕(豆粕-棉籽粕-菜籽粕)型饲粮。饲养试验共计91 d,其中预试期7 d,正试期84 d。玉米-杂粕型饲粮组成及营养水平见表1。饲粮中粗蛋白质、钙、总磷、氨基酸含量分别参照GB/T 6432—2018、GB/T 6436—2018、GB/T 6437—2018和GB/T 18246—2019测定。

表1 玉米-杂粕型饲粮组成及营养水平(风干基础)
1.3 饲养管理
试验采用阶梯式笼养,每笼3只鸡,各个重复均匀分布,试验鸡只全部饲养于同一鸡舍内。每天加料2次(08:00、15:00),终日保持饲料器内有料。乳头式饮水器自由饮水,每天16:00—17:00清粪和打扫卫生1次,每周对鸡舍进行带鸡喷雾消毒1次,定期对鸡舍周围进行喷雾消毒。鸡群的免疫程序按照常规程序进行。
1.4 测定指标及方法
1.4.1 生产性能
以重复为单位记录每周耗料量,每天16:00收蛋,记录各重复产蛋数、总蛋重,整理计算以下指标。
产蛋率=[总蛋数/(试验鸡数×试验天数)]×100; 平均蛋重=每个重复总蛋重/每个重复产蛋总数;平均日采食量=(每周每个重复喂料量-每周每个重复剩料量)/(重复内鸡只数×每周天数);料蛋比=试验期内每个重复耗料总量/试验期内每个重复产蛋总重。
1.4.2 蛋品质
试验结束当天,每个重复随机取3枚鸡蛋并标号,进行蛋品质检测。
蛋黄重:电子天平测量,精确到0.01 g。
蛋白高度:全自动蛋品质分析仪(EMT-5200 Robotmation公司,日本)测定。
蛋壳强度:蛋壳强度测定仪(EFG-0503,Robotmation公司,日本)测定。
蛋壳厚度:螺旋测微器分别测量除去内外壳膜的蛋壳钝端、中部、锐端3部位的厚度,取平均值,精确到0.01 mm。
蛋黄颜色:全自动蛋品质分析仪测定。
蛋形指数:用全自动蛋品质分析仪测量鸡蛋纵径和横径,纵径与横径比值即为蛋形指数,精确到0.1 mm。
哈氏单位:全自动蛋品质分析仪测定。
1.4.3 肝脏中丙酮酸激酶及磷酸果糖激酶活性
于试验结束当天,每个重复随机选取1只蛋鸡,颈动脉放血致死后立刻解剖,取其肝脏组织至冻存管,液氮速冻后转移至-80 ℃超低温冰箱中保存待测。肝脏丙酮酸激酶及磷酸果糖激酶活性测定具体步骤均按照试剂盒说明书进行,检测试剂盒(试剂盒编号:A076-1-1、A129-1-1)均购自南京建成生物工程研究所。
1.4.4 十二指肠、空肠中α-淀粉酶及胰蛋白酶活性
于试验结束当天,每个重复随机选取1只蛋鸡,颈动脉放血致死后立即解剖,轻轻挤出十二指肠、空肠食糜至冻存管,液氮速冻后转移至-80 ℃超低温冰箱中保存待测。十二指肠、空肠中α-淀粉酶及胰蛋白酶活性测定具体方法参照试剂盒说明书进行,检测试剂盒(试剂盒编号:016-1-1、A080-2-2)购于南京建成生物工程研究所。
1.4.5 十二指肠、空肠中葡萄糖转运载体mRNA相对表达量
于试验结束当天,每个重复随机选取1只蛋鸡,颈动脉放血致死后立即解剖,分离小肠,沿纵向剖开,用生理盐水冲洗,吸水纸吸干。剪取十二指肠、空肠中段约3 cm作为组织样品,置于液氮中速冻,取出后于-80 ℃冷冻保存。取100 mg组织抽提总RNA,通过研磨、离心、孵育等操作以及添加三氯甲烷、异丙醇、乙醇等溶液最终将提取的RNA稀释至200 ng/μL;配制逆转录反应体系(5×SweScript All-in-One SuperMix for qPCR 4 μL、gDNA Remover 1 μL、RNA 10 μL、RNase free water 20 μL),轻轻混匀并离心,设置逆转录程序,于普通PCR仪上完成逆转录;取0.1 mL PCR反应板,配制反应体系[2×SYBR Green qPCR Master Mix (None ROX) 7.5 μL、2.5 μmol/L基因引物(上游+下游)1.5 μL、反转录产物(cDNA)2.0 μL、Water Nuclease-Free 4.0 μL],每个反转录产物配制3管;点完样用PCR封板膜配合封膜仪完成封膜,于荧光定量PCR仪上完成扩增。以β-肌动蛋白(β-actin)作为参比基因,采用2-ΔΔCt法计算目的基因钠-葡萄糖协同转运蛋白-1(SGLT-1)和葡萄糖转运蛋白-2(GLUT-2)的mRNA相对表达量。PCR引物信息见表2。

表2 PCR引物信息
1.5 数据统计与分析
试验数据用Excel 2019进行整理汇总,采用SPSS 22.0统计软件进行单因素方差分析(one-way ANOVA),并采用Duncan氏法进行组间多重比较。结果用平均值±标准误(mean±SE)表示,P<0.05为差异显著。
2 结果与分析
2.1 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡生产性能的影响
由表3可知,与对照组相比,400 mg/kg组产蛋率显著提高了5.71%(P<0.05),料蛋比显著降低了8.90%(P<0.05),其他试验组无显著变化(P>0.05)。与对照组相比,400 mg/kg组与600 mg/kg组平均日采食量分别显著降低了3.26%与3.09%(P<0.05)。各组的平均蛋重无显著差异(P>0.05)。
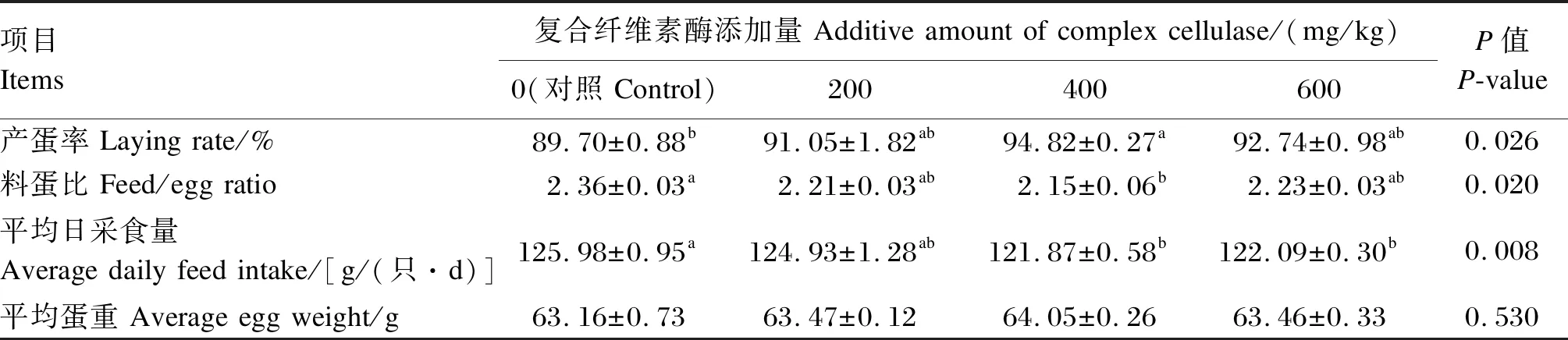
表3 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡生产性能的影响
2.2 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡蛋品质的影响
由表4可知,与对照组相比,玉米-杂粕型饲粮中添加不同水平复合纤维素酶对蛋鸡的蛋壳厚度、蛋壳强度、蛋白高度以及哈氏单位等各项蛋品质指标均无显著影响(P>0.05)。
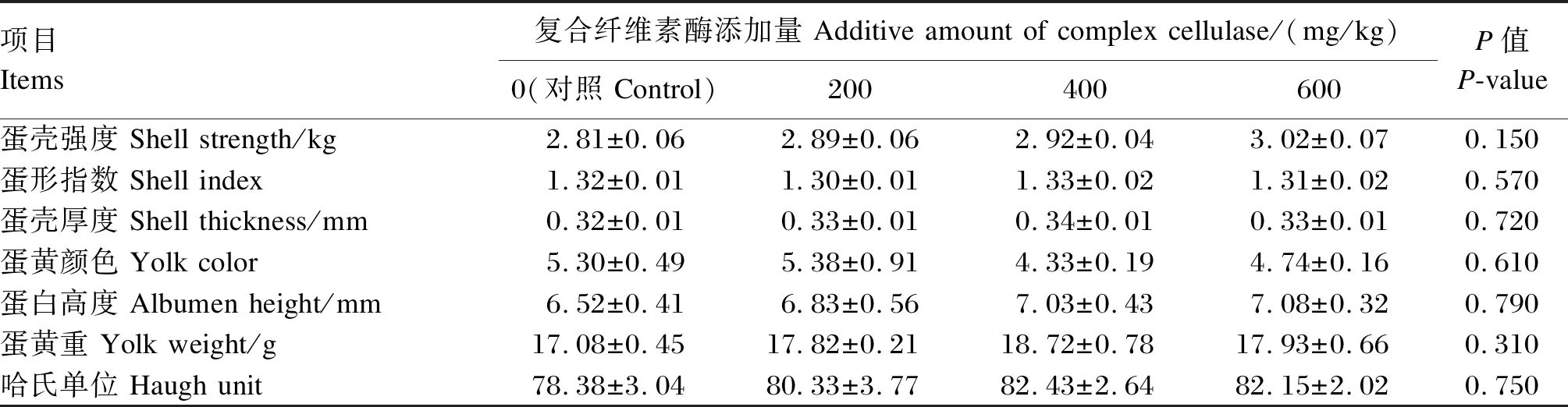
表4 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡蛋品质的影响
2.3 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肝脏中糖代谢相关酶活性的影响
由表5可知,与对照组相比,400 mg/kg组和600mg/kg组肝脏中丙酮酸激酶活性分别提高70.71%、53.04%,差异显著(P<0.05)。各试验组肝脏中磷酸果糖激酶活性与对照组相比均有所提高,但差异均不显著(P>0.05)。

表5 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肝脏中糖代谢相关酶活性的影响
2.4 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡内源消化酶活性的影响
由表6可知,与对照组相比,玉米-杂粕型饲粮中添加400 mg/kg复合纤维素酶可以显著提高空肠中淀粉酶活性(P<0.05)。各试验组十二指肠中淀粉酶活性与对照组相比差异均不显著(P>0.05)。与对照组相比,各试验组十二指肠和空肠中胰蛋白酶活性均有所提高,其中400 mg/kg组空肠胰蛋白酶活性比对照组提高160.69%,差异显著(P<0.05)。
2.5 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肠道中葡萄糖转运载体基因表达的影响
由图1可知,与对照组相比,玉米-杂粕型饲粮中添加400或600 mg/kg复合纤维素酶均可以显著提高蛋鸡空肠中SGLT-1 mRNA相对表达量(P<0.05);各试验组蛋鸡十二指肠中SGLT-1 mRNA相对表达量与对照组相比差异不显著(P>0.05);此外,各组蛋鸡空肠中SGLT-1 mRNA相对表达量整体比十二指肠高。与对照组相比,玉米-杂粕型饲粮中添加不同水平复合纤维素酶对蛋鸡空肠、十二指肠中GLUT-2 mRNA相对表达量均无显著影响(P>0.05),但在数值上以400 mg/kg组较高。
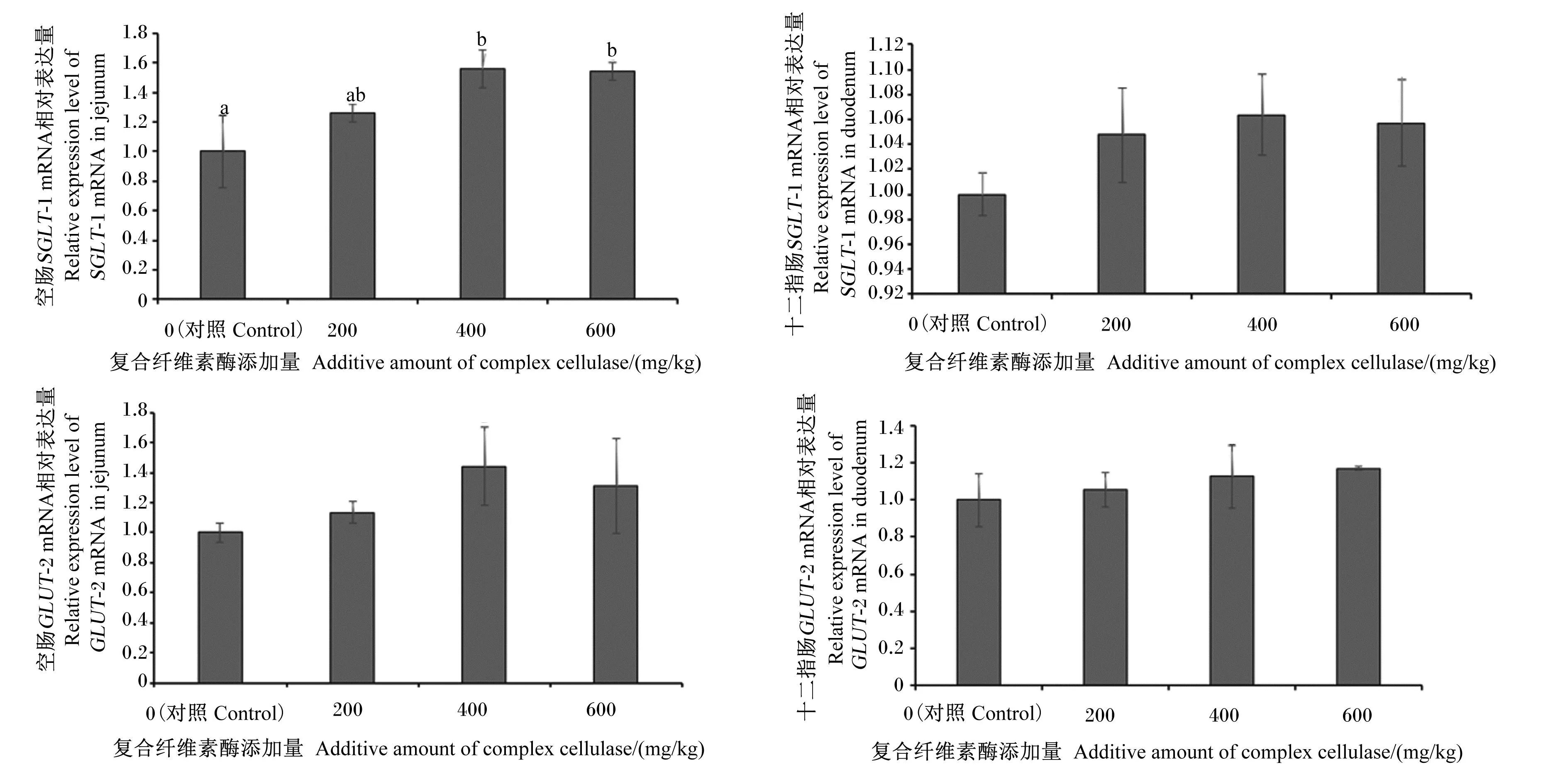
数据柱形标注不同小写字母表示差异显著(P<0.05),无字母或相同字母表示差异不显著(P>0.05)。
3 讨 论
3.1 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡生产性能和蛋品质的影响
目前畜禽大多使用植物性饲料,其共同特点是有大量细胞壁。由于鸡自身不能分泌分解细胞壁的酶,导致细胞壁内养分不能被充分利用,不仅造成饲料资源的浪费,同时还增加了养殖成本。本试验使用的复合纤维素酶由一系列针对分解细胞壁的酶复合而成,可以通过破坏细胞壁结构,使细胞壁内养分充分释放,提高饲料利用率,进而提高蛋鸡生产性能。焦莉等[10]的研究表明,低营养水平饲粮中添加复合酶可以显著降低蛋鸡的料蛋比;Oba等[11]的研究表明,添加复合纤维素酶可以显著提高蛋鸡的产蛋率。本试验结果与上述研究结果一致。Bederska-ojewska等[12]研究显示,在黑麦饲粮中添加复合酶制剂对蛋鸡的产蛋率无显著影响。本试验结果与其不同,其原因可能是使用的酶制剂不同导致结果不同,本试验选用的复合纤维素酶可以有效破坏细胞壁中的β-1,4糖苷键等化学键[13]。海存秀等[14]研究发现,在杂粕型饲粮中添加复合酶制剂可以显著降低蛋鸡的采食量与料蛋比。本试验结果与上述研究结果相同,由于家禽具有为能而食的特点,添加复合纤维素酶可以提高饲粮中养分的释放量,降低采食量与料蛋比。
饲粮中添加酶制剂会从不同方面影响蛋品质。张胜[15]的研究表明,蛋鸡饲粮中添加饲用酶制剂可以一定程度上提高蛋壳厚度,降低破蛋率。黄靓[16]的研究显示,复合酶制剂对海兰褐蛋鸡蛋黄颜色、蛋壳强度、蛋壳厚度均无显著影响,本试验结果与该研究结果一致。
3.2 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肝脏糖代谢的影响
肝脏是动物机体重要的代谢器官,是糖、脂代谢的主要场所,在动物营养物质代谢过程中发挥着重要作用[17]。动物机体主要的糖代谢途径有糖酵解、糖异生、三羧酸循环、磷酸戊糖途径等,其中糖酵解是动物机体分解葡萄糖最主要的途径之一。丙酮酸激酶可催化磷酸烯醇式丙酮酸和ADP生成ATP和丙酮酸,磷酸果糖激酶可催化果糖-6-磷酸和ATP生成果糖-1,6-二磷酸和ADP,二者都是糖酵解途径中的限速酶。由于鱼类对葡萄糖利用能力低下,因此目前关于饲粮中碳水化合物水平对肝脏糖代谢影响的研究主要集中在鱼类上,在蛋鸡方面的研究较少,但二者吸收利用葡萄糖的机制类似[18]。有报道指出,饲粮中碳水化合物水平与肝脏丙酮酸激酶活性关系密切[19-21]。Zhou等[17]研究发现,随着饲粮中碳水化合物水平提高,金鲚鱼肝脏中丙酮酸激酶活性显著提高。陆游等[22]研究表明,控制饲粮中脂肪水平为5%时,随着饲粮中淀粉含量的增加,大黄鱼肝脏中丙酮酸激酶活性显著提高。本试验结果与上述结论基本一致。本试验通过添加复合纤维素酶使饲粮中养分的释放量增多,可被蛋鸡利用的有效养分增多,即酶的底物增多,肝脏中丙酮酸激酶活性随之提高。聂琴等[23]研究发现,大菱鲆肝脏中磷酸果糖激酶活性不受饲粮中碳水化合物水平的影响。与上述结论一致,在玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肝脏中磷酸果糖激酶活性无显著影响。有研究报道,磷酸果糖激酶非常保守,其活性不受饲粮中碳水化合物水平的诱导[24],这可能是肝脏中磷酸果糖激酶活性变化不显著的原因。由于添加酶制剂对蛋鸡肝脏中丙酮酸激酶活性影响的研究鲜有报道,其具体原因有待进一步研究。
3.3 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡内源消化酶活性的影响
小肠是家禽消化吸收饲粮中养分的主要场所,肠道中的淀粉酶和胰蛋白酶将饲粮中的淀粉和蛋白质分解成葡萄糖和多肽等小分子物质,有助于机体利用。
温超[25]通过试验发现,饲粮中添加复合酶制剂后产蛋后期蛋鸡各肠段中淀粉酶活性以及空肠中胰蛋白酶活性有所提高,但差异不显著,而十二指肠中胰蛋白酶活性则有下降趋势。本试验结果与其不同,其可能是由于使用的酶制剂具体组成成分不同造成最终试验结果不同,温超[25]的试验中使用的复合酶制剂主要由纤维素酶、植酸酶和木聚糖酶复合而成。陈加敏[26]研究发现,底物食糜的浓度与内源酶活性呈正相关。White等[27]研究发现,外源酶对内源酶的作用机制不仅是直接提高内源酶分泌量,并且可以间接通过调节底物的量对内源酶进行调控。本试验中,外源性添加的复合纤维素酶通过分解细胞壁,释放更多壁内养分,增加了肠道内可消化底物浓度,进而刺激内源消化酶分泌,提高内源消化酶活性。随着复合纤维素酶添加量的增加,各个肠段的内源消化酶活性不断提高,达到最大值后又出现降低的趋势。本试验结果与Rebolé等[28]的研究结果一致,分析原因为当复合纤维素酶添加量不足时,导致细胞壁降解不完全,可消化底物浓度低,因此消化酶活性低,当复合纤维素酶添加量过高时,达到底物消化的饱和量,会产生负反馈抑制,影响养分的消化吸收,生产性能等指标也印证了该趋势。
3.4 玉米-杂粕型饲粮中添加复合纤维素酶对蛋鸡肠道中葡萄糖转运载体基因表达的影响
进入动物机体的碳水化合物会在小肠被淀粉酶等消化酶分解为单糖,而单糖需要相应的转运载体转运才能最终被机体吸收[29]。小肠黏膜上皮细胞吸收的单糖包括葡萄糖、半乳糖和果糖,其中葡萄糖占80%,被禽类吸收的单糖中葡萄糖的占比达98%以上[30]。有研究表明,小肠上皮细胞中营养物质转运载体的表达量直接影响养分吸收[31]。单糖转运载体主要有SGLT-1和GLUT-2,主要转运葡萄糖、果糖、半乳糖。SGLT-1在葡萄糖转运过程中起主导作用,主要分布在近端小肠上皮细胞刷状缘膜上,通过激活蛋白激酶C(PKC)依赖性通路调节GLUT-2对葡萄糖的转运[32]。mRNA作为指导蛋白质合成的模板,其相对表达量增多意味着葡萄糖转运载体增多,被机体吸收利用的养分增多。王修启等[33]研究发现,饲粮中添加木聚糖酶可以显著提高肉仔鸡十二指肠中SGLT-1 mRNA相对表达量,对空肠中SGLT-1 mRNA相对表达量的影响不显著,该结果与本试验结果不同。Barfull等[34]研究表明,SGLT-1 mRNA相对表达量和鸡日龄与肠腔面积有关,且均呈正相关。本试验选用的蛋鸡比肉仔鸡肠腔面积更大,营养物质与肠道接触面积更大,因此在本试验中添加复合纤维素酶对蛋鸡空肠中SGLT-1 mRNA相对表达量有显著影响。SGLT-1 mRNA相对表达量受底物葡萄糖浓度的影响[35]。本试验中,添加400 mg/kg复合纤维素酶组蛋鸡空肠中淀粉酶活性较对照组显著提高,因此其空肠中SGLT-1 mRNA相对表达量显著高于对照组。王修启等[33]研究发现,与对照组相比,饲粮中添加木聚糖酶对肉鸡十二指肠和空肠中GLUT-2 mRNA相对表达量无显著影响。本研究结果符合前人研究结论。有报道表明,GLUT-2不仅分布在小肠,在肝脏、肾脏、胰脏、大脑中均表达[36-37],这可能是添加复合纤维素酶后蛋鸡肠道中GLUT-2 mRNA相对表达量不显著的原因。周力等[38]的研究发现,精料比例越高,青海黑藏羊小肠中SGLT-1 mRNA相对表达量越高。复合纤维素酶可分解细胞壁降低饲粮纤维含量,提高饲粮中养分释放量,因此各试验组肠道中葡萄糖转运载体mRNA相对表达量均高于对照组,与前人研究结果一致。
4 结 论
在玉米-杂粕型饲粮中添加复合纤维素酶可以通过提高饲粮中营养物质释放量提高蛋鸡小肠中消化酶及肝脏中糖代谢相关酶活性,还能上调肠道中葡萄糖转运载体基因的表达,进而提高生产性能。本试验条件下,蛋鸡玉米-杂粕型饲粮中复合纤维素酶的最适添加量为400 mg/kg。
