谷氨酸的生理功能及其在畜禽生产中的应用研究进展
2024-03-05黄钰琴王力仪单体中
黄钰琴 王力仪 单体中
(浙江大学动物科学学院,浙江大学动物分子营养学教育部重点实验室,杭州 310058)
蛋白质是畜禽生长及生产不可或缺的营养物质,为满足畜禽更高生产效率的需求,生猪养殖业对饲料中蛋白质的营养价值和生理需要也越来越高[1]。氨基酸是蛋白质和小肽的组成成分,根据其能否在动物体内被合成分为必需氨基酸与非必需氨基酸[2]。随着对氨基酸营养的深入研究,发现部分氨基酸是动物生长、发育、免疫、繁殖及维持机体稳态关键代谢途径的重要调节因子,亦被称为功能性氨基酸[3]。
谷氨酸(glutamic acid,Glu)是食物和动物组织中含量最丰富的非必需氨基酸之一,以游离或蛋白质结合形式存在于肉类、乳制品、蔬菜、豆类等产品中[4]。左旋构型的游离Glu具有增味作用,食物在腌制、老化、成熟和烹饪过程中释放的游离Glu可给食物带来鲜味[5]。作为合成谷胱甘肽和谷氨酰胺(glutamine,Gln)的前体物质,Glu可用于合成蛋白质与其他非必需氨基酸,也在介导兴奋性突触传递、维持肠道功能完整性、调控能量稳态中发挥重要作用[6-8]。近年来,Glu在畜禽生产中所发挥的营养作用越来越受关注[7],本文就Glu的理化性质、生理功能及其在畜禽生产上的应用研究进展进行综述,对后续利用营养手段提高生猪生产性能并改善猪肉品质具有重要意义。
1 Glu的理化性质
Glu的化学名称为α-氨基戊二酸,化学式为C5H9NO4,分子内含2个羧基,是一种酸性氨基酸,有左旋体、右旋体和外消旋体3种不同构型。Glu为无色晶体,微溶于水,而溶于盐酸溶液,等电点3.22。可提供负电荷的Glu残基在参与蛋白质合成时可起到稳定蛋白质结构的重要作用,还能以多聚谷氨酰化参与蛋白质翻译后修饰作用[8]。
作为非必需氨基酸,Glu可在动物体内大量合成[9]。膳食中的Glu主要经肠道进行代谢,只有少数膳食Glu会被吸收进入门静脉血中,这也使得血浆Glu水平受膳食Glu水平影响较小[8]。膳食Glu主要在肠道被代谢,机体其他组织的Glu需经内源性合成而来。Glu可在绝大多数细胞内经不同的酶与底物生成,其可由细胞内蛋白质降解所产生,也可由Gln在谷氨酰胺酶(glutaminase,GLS)或果糖-6-磷酸转氨酶的作用下生成;除此之外,Glu的重要前体物质α-酮戊二酸可在支链氨基酸转氨酶(branched-chain aminotransferase,BCAT)、天冬氨酸转氨酶(aspartate aminotransferase,AST)、丙氨酸转氨酶(alanine aminotransferase,ALT)和谷氨酸脱氢酶(glutamate dehydrogenase,GDH)的作用下与支链氨基酸、草酰乙酸、丙酮酸盐和铵离子反应生成Glu[7]。
2 Glu的生理功能
Glu具有广泛的生理功能,它既是重要的神经递质以及肠道与大脑的能量底物,还在维持动物肠道功能完整性和调控机体能量稳态上发挥重要作用[6-8](图1)。
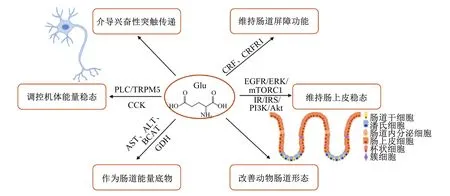
Glu:谷氨酸 glutamic acid;PLC/TRPM5:磷脂酶C/瞬时受体电位M5 phospholipase C/transient receptor potential M5;CCK:胆囊收缩素 cholecystokinin;AST:天冬氨酸转氨酶 aspartate aminotransferase;ALT:丙氨酸转氨酶 alanine aminotransferase;BCAT:支链氨基酸转氨酶 branched-chain aminotransferase;GDH:谷氨酸脱氢酶 glutamate dehydrogenase;CRF:促肾上腺皮质素释放因子 corticotrophin-releasing factor;CRFR1:CRF受体1 CRF receptor 1;IR/IRS/PI3K/Akt:胰岛素受体/胰岛素受体底物/磷酸肌醇3-激酶/蛋白激酶B途径 insulin receptor/insulin receptor substrate/phosphoinositide 3-kinase/protein kinase B pathway;EGFR/ERK/mTORC1:表皮生长因子受体/细胞外调节蛋白激酶/哺乳动物雷帕霉素靶蛋白复合物1信号通路 epidermal growth factor receptor/extracellular regulated protein kinase/mechanistic target of rapamycin complex 1 signaling pathway。
2.1 作为重要的神经递质
Glu是最重要的神经递质之一,是大脑中最丰富的游离氨基酸,在发育中的大脑中通过促进钙离子(Ca2+)转运在神经元分化、迁移和存活中发挥重要作用[10]。Glu还是脊椎动物中枢神经系统的主要兴奋性神经递质,介导中枢神经系统中大多数兴奋性突触传递,在突触可塑性、学习及记忆等认知功能起着关键作用[11]。Glu主要作用于大脑内的突触前神经元、突触后神经元和神经胶质细胞。大脑中大多数兴奋性突触的信息传递由突触前细胞释放Glu介导,在Glu扩散到突触后神经元细胞后,可激活阳离子通道或G蛋白连接受体,从而改变突触后神经元细胞的膜电位或细胞内生物化学水平[12]。为了避免过量Glu造成神经兴奋性毒性并保持大脑神经元能够高速传递信息,星形神经胶质细胞中表达的高亲和力的谷氨酸转运蛋白可从细胞外清除Glu以保证Glu的突触后作用被迅速终止[13],并保持细胞外Glu水平远低于神经元兴奋性毒性水平[14]。
2.2 作为肠道与大脑的能量底物
膳食中的Glu主要经肠腔吸收,胃肠道具有非常高的Glu利用能力,从肠腔吸收的Glu大部分被小肠上皮细胞通过AST、ALT、BCAT和GDH分解后经多种代谢途径利用,包括生成支持肠道运动和功能的能量[15]。作为胃肠道中重要的能量底物来源,约35%的肠黏膜总能量需求来自Glu氧化为二氧化碳(CO2)所产生的能量[4]。肠道以动脉Gln或膳食Glu为底物时,约90%的碳以CO2形式释放;而以葡萄糖作为底物时,只有34%的碳以CO2形式释放,表明膳食Glu是肠道内更为重要的氧化底物[16]。
大脑是中枢神经系统中最大和最复杂的部位,是调节机体功能和各种高级神经活动的物质基础,需要各种营养物质提供能量。因血脑屏障的限制,除了葡萄糖作为大脑的主要供能物质以外,部分脂肪酸和氨基酸也可为大脑提供能量,而Glu也被认为有助于维持大脑能量稳态[17]。如星形胶质细胞可主动吸收细胞外Glu并以GDH依赖性的方式氧化Glu生成ATP[18]。
2.3 维持动物肠道功能完整性
Glu对维持动物肠道形态、肠道屏障及肠道功能完整性具有重要作用[19]。肠道黏膜屏障由肠上皮细胞及细胞间紧密连接蛋白等结构组成,Glu可通过促进肠上皮细胞增殖、减少细胞凋亡和维持膜完整性以维持肠粘膜屏障功能[20]。Glu还可增加小肠绒毛高度、绒毛高度与隐窝深度之比以改善动物肠道形态[21]。Glu可减轻脂多糖(lipopolysaccharide,LPS)诱导的肠道损伤并下调促肾上腺皮质素释放因子(corticotrophin-releasing factor,CRF)、CRF受体1(CRF receptor 1,CRFR1)的mRNA表达水平,表明Glu减轻LPS诱导的肠黏膜屏障损伤可能与调节CRF信号通路有关[22]。体外试验表明,Glu可通过胰岛素受体(insulin receptor,IR)/胰岛素受体底物(insulin receptor substrate,IRS)/磷酸肌醇3-激酶(phosphoinositide 3-kinase,PI3K)/蛋白激酶B(protein kinase B,Akt)途径激活哺乳动物雷帕霉素靶蛋白复合物1(mechanistic target of rapamycin complex 1,mTORC1)信号通路发挥作用并促进肠道干细胞(intestinal stem cells,ISCs)扩张[23];Glu可提高ISCs活性并以表皮生长因子受体(epidermal growth factor receptor,EGFR)/细胞外调节蛋白激酶(extracellular regulated protein kinase,ERK)/mTORC1信号机制依赖性的方式促进ISCs的增殖与分化,进而维持肠上皮的稳态与再生[24];而缺乏Glu则通过下调哺乳动物雷帕霉素靶蛋白(mammalian target of rapamycin,mTOR)/核糖体蛋白S6激酶1(ribosomal protein S6 kinase,S6K1)通路和兴奋性氨基酸转运蛋白3(excitatory amino acid transporter 3,EAAT3)表达抑制猪肠上皮细胞增殖,并导致仔猪肠上皮更新障碍[25]。
2.4 调控机体能量稳态
作为多功能氨基酸,Glu在激活肠脑轴和维持机体能量稳态中发挥着新作用[26]。大鼠长期摄入1%谷氨酸钠(monosodium glutamate,MSG)溶液可通过增加能量消耗来减少体重增加、脂肪沉积和血浆瘦素浓度升高[27]。Glu可调节机体食物的摄入与饱腹感。大鼠口服MSG后通过激活G蛋白偶联受体T1R1/T1R3以磷脂酶C(phospholipase C,PLC)/瞬时受体电位M5(transient receptor potential M5,TRPM5)依赖的方式刺激胆囊收缩素(cholecystokinin,CCK)分泌,进而抑制食物的摄入[28]。MSG与蛋白质一起摄入时可增强餐后饱腹感[29],Glu与5′-核苷酸单磷酸盐特别是5′-单磷酸肌苷(inosine 5′-monophosphate,IMP)结合时具有增强鲜味的协同作用[30],而MSG与IMP的协同作用还可增强饱腹感[31]。
位于脑干中缝的中缝背核(dorsal raphe nucleus,DRN)与食物摄入控制有关,抑制DRN中表达囊泡谷氨酸转运体3(vesicular glutamate transporter 3,VGLUT3)的谷氨酸能神经元(DRNVGLUT3神经元)的活性可显著减少肥胖小鼠的食物摄入[32]。丘脑外侧区(lateral hypothalamic area,LHA)是一个负责协调机体进食的高度保守的大脑区域,LHA中对饱腹状态更为敏感的表达囊泡谷氨酸转运体2(vesicular glutamate transporter 2,VGLUT2)的谷氨酸能神经元(LHAVGLUT2神经元)在机体进食欲望较低时其神经兴奋性更强,高脂饮食会导致LHAVGLUT2神经元兴奋性降低,进而破坏内源性进食抑制系统的功能[33]。
Glu还参与调控机体对糖的摄入。虽然甜味是由舌头检测到,但摄入糖的需求是由肠道产生的。一些被称为神经足细胞的十二指肠细胞可以使用钠-葡萄糖共转运蛋白1(sodium-glucose co-transporter 1,SGLT1)检测到葡萄糖,并将Glu释放到迷走神经传入神经元上,引发兴奋性突触后电流,再进一步调控机体对糖的摄入[34]。
3 Glu在畜禽生产中的应用研究进展
3.1 Glu在猪生产中的应用
3.1.1 Glu对猪生长性能的影响
生长性能是生猪生产的重要性状之一[35]。Rezaei等[36]的研究发现,在21日龄断奶仔猪饲粮中添加1%、2%、4%的MSG,剂量依赖性地增加了断奶后仔猪的体重、日增重和饲料效率,显著提高了仔猪断奶后的生长性能。Liu等[37]的研究发现,在杜长大仔猪饲粮中补充1% Glu后,其平均日增重与平均日采食量显著增加。胡诚军等[38]的研究发现,在77 kg育肥猪基础饲粮中添加1% Glu与1.44%丙氨酸(alanine,Ala),饲喂60 d后显著降低了育肥猪的末重与背膘厚,降低了育肥猪的脂肪率,提高了试验第1~30天的平均日增重,显著降低了试验第1~60天的平均日采食量,并显著提高了试验第1~30天的料重比,提高了育肥猪的生长性能。以上研究结果表明,Glu对改善猪生长性能起着重要作用。
低蛋白质饲粮可通过合理配合饲料及添加适宜种类和数量的工业氨基酸,达到降低饲料成本及减低氮排放的目的[39]。Zimmerman[40]研究发现,在21日龄断奶仔猪低蛋白质饲粮中添加3.36% Glu可使断奶仔猪的日增重增加12%,且不会影响料重比。Upadhaya等[41]的研究发现,在杜长大断奶仔猪减少2%粗蛋白质饲粮中补充Glu后,仔猪的体重、平均日增重和饲料转化率显著提高,表明补充Glu可补偿基础饲粮中降低2%的粗蛋白质。Upadhaya等[42]研究发现,在杜长大生长猪减少2%粗蛋白质的低蛋白质饲粮中添加赖氨酸(lysine,Lys)与Glu,当Lys与Glu的比例达1∶2.71时,即添加6.8 g/kg的Glu可获得与饲喂标准基础饲粮的对照组杜长大生长猪相似的生长性能和胴体性状。甄吉福等[43]的研究发现,在杜长大育肥猪低蛋白质饲粮中添加Glu可显著降低育肥猪的尿氮和总氮排放量,提高蛋白质利用效率,且对育肥猪的生长性能无负面影响。以上研究结果提示,在低蛋白质饲粮中补充适量Glu可补偿基础饲粮中粗蛋白质的降低带来的生猪生长性能的下降。
3.1.2 Glu对猪肉品质的影响
肉品质如肌内脂肪(intramuscular fat,IMF)含量、肉色、嫩度等指标是影响消费者决策的重要因素[44]。胡诚军等[38]的研究发现,在育肥猪基础饲粮中添加1% Glu与1%亮氨酸(leucine,Leu)对其生长性能无显著影响,但显著提高了IMF的含量,改善了猪肉品质。Hu等[45]在杜长大生长育肥猪基础饲粮中添加1% Glu与1%精氨酸(arginine,Arg)发现对其生长性能无负面影响,但显著降低了育肥猪的背膘厚并提高背最长肌中IMF的含量,显著改善了背最长肌的肉色及脂肪酸组成。Glu还可以通过促进背部脂肪组织中脂解相关基因表达及背最长肌中脂肪生成基因表达的形式降低背膘厚并促进肌肉中IMF沉积[46]。Guo等[47]研究发现,添加1% Glu与1% Arg可显著增加IMF含量,改善肉的嫩度和多汁性,提高了猪肉的整体食用品质。杨翠等[48]在育肥猪基础饲粮中补充0.6% Glu及大豆磷脂发现对育肥猪的生长性能和屠宰性能无显著影响,但显著提高了肌肉中总鲜味氨基酸及IMF的含量,提高了猪肉品质。但也有研究发现,在生长猪基础饲粮或高脂饲粮中补充MSG会引起背部脂肪沉积的增加,并影响脂肪酸在机体组织的分布和代谢以及机体感知脂肪酸的能力[49]。以上研究结果表明,Glu在提高猪肉品质上也具有潜在的应用前景。
肌肉发育是影响猪生长性能与肉品质的重要生物学过程,骨骼肌萎缩相关疾病会导致循环LPS水平升高,LPS可诱导肌肉蛋白质合成下降与蛋白质降解增加,进一步诱导肌肉蛋白质损失与肌肉萎缩[50]。研究发现,LPS诱导组断奶仔猪饲粮中补充Glu,饲喂28 d后可促进肌细胞蛋白质合成能力,并通过抑制Toll样受体4(Toll-like receptor 4,TLR4)、核苷酸结合寡聚结构域蛋白(nucleotide-binding oligomerization domain protein,NOD)信号传导相关基因的mRNA表达以及激活Akt/叉头框蛋白O (forkhead box O,FOXO)和mTOR信号通路来缓解肌肉LPS诱导处理引起的肌肉蛋白质损失[51]。以上研究结果表明,Glu在促进肌肉蛋白质合成上亦有着重要作用。
3.1.3 Glu对猪肠道健康的影响
作为机体中重要的消化器官,肠道健康影响猪各个阶段的生长性能[52]。研究发现,口服MSG可增强哺乳仔猪肠道中Glu信号受体和谷氨酸转运蛋白的表达[53],改善哺乳仔猪肠道形态并增加盲肠和结肠中挥发性脂肪酸含量,提高仔猪肠道的吸收能力[21]。Wang等[54]的研究发现,在基础饲粮中补充Gln、Glu与Asp可显著提高仔猪断奶后21 d的体重与平均日增重,改善断奶仔猪的肠黏膜屏障结构并改善肠道能量稳态。Qin等[55]在断奶仔猪无氮饲粮或基础饲粮中补充1% Glu后发现可提高小肠重量并提高饲粮表观消化率,Glu通过与跨膜受体卷曲蛋白7(frizzled 7,FZD7)结合激活Wnt/β-连环蛋白(β-catenin)信号通路以促进仔猪肠道上皮生长并调控ISCs的增殖和分化。AG补充剂是一种含10% Glu与10% Gln的膳食补充剂,Cabrera等[56]研究发现,在14日龄断奶仔猪断奶后前3周的开口料与保育料中添加0.88%的AG补充剂可提高仔猪的饲料转化率,而这可能与其提高仔猪的肠道功能相关。Luise等[57]研究发现,在24~45日龄断奶仔猪饲粮中补充Glu和Gln可通过促进仔猪肠道免疫及屏障功能促进仔猪断奶后的生长;另一研究发现,在28~49日龄断奶仔猪饲粮中补充1%的Glu与Gln显著提高了其体重增长,并降低了腹泻的发生率,Gln与Glu可提高肠道细胞增殖率,促进肠黏膜修复并维持肠上皮完整性,进一步促进肠道营养吸收与仔猪体重的增加[58]。以上研究结果表明,在基础饲粮中补充Glu有利于促进仔猪肠道发育,提高肠道功能,改善肠道营养吸收效率,在提高仔猪生产效率上有着良好的应用前景。
仔猪肠道氧化应激是阻碍仔猪生长性能提高的重要因素[59]。仔猪饲粮中补充Glu可通过激活mTOR信号通路并抑制TLR4和NOD信号通路以减轻LPS引起的仔猪肠道炎症与损伤并促进肠道修复[60];Glu还可抑制具有促炎作用的辅助性T(T helper,Th)17细胞的分化并促进免疫调节剂调节性T(regulatory T,Treg)细胞的功能,通过Th17细胞/Treg细胞平衡相关免疫信号通路增强仔猪的肠道免疫功能,缓解因LPS急性损伤诱导的仔猪肠道炎症[37]。饲粮中补充Glu可缓解除草剂诱导的氧化应激并恢复仔猪体重,提高血清中超氧化物歧化酶活性和总抗氧化能力以及抑制脂质氧化与丙二醛的生成,并提高肠道中氨基酸转运蛋白的表达[61]。在低蛋白质饲粮中补充Glu和Arg可缓解过氧化氢诱导的仔猪肠道氧化应激反应,保护仔猪免受氧化应激诱导的肠道功能障碍[62]。以上结果提示,Glu在缓解仔猪氧化应激反应、维持肠上皮稳态、促进肠道健康与功能上发挥着营养调节剂作用。
饲料霉变会引起猪肠道屏障损伤并引起生长性能下降,且被霉菌污染的饲料含有多种真菌毒素,可对生猪养殖产生更为严重的影响[63]。Duan等[64]在饲喂霉变饲料的生长猪饲粮中补充2% Glu,发现其可改善真菌毒素引起的生长猪抗氧化系统失衡和肠道结构异常,并提高饲料转化效率,对恢复生长猪的生长性能具有积极影响。Wu等[65]研究发现,在仔猪饲粮中补充2% Glu可减轻呕吐毒素(deoxynivalenol,DON)引起的氧化应激与肠道损伤,并降低DON对Akt/mTOR/真核翻译起始因子4E结合蛋白1(eukaryotic initiation factor 4e-binding protein 1,4EBP1)信号通路的抑制作用。以上研究结果表明,Glu可能是一种可改善真菌毒素引起的不良反应的营养调节因子。
3.1.4 Glu对母猪繁殖性能的影响
妊娠早期胎盘发育和功能及妊娠期内子宫容量是限制母猪产仔数的重要因素[7]。N-氨甲酰谷氨酸(N-carbamylglutamate,NCG)由Glu与乙酰辅酶A经乙酰谷氨酸合成酶催化生成[66],母猪妊娠早期补充NCG可提高早期胚胎存活率[67],妊娠及哺乳期补充NCG可提高窝产健仔数,并提高母猪哺乳期采食量及断奶仔猪生长性能[68],提高活产仔猪数与窝重,改善母猪繁殖性能[69],提示Glu及其代谢物在提高妊娠母猪繁殖性能上也发挥着重要作用。妊娠后期胎儿生长迅速,Wu等[70]研究发现,在母猪妊娠90~114 d期间基础饲粮中补充1% Gln可以改善胎儿生长迟缓的现象并提高产奶量,还可降低仔猪断奶前的死亡率。由于机体内Gln与Glu可相互转换,因此Glu也被认为是母猪妊娠和泌乳期间的条件必需氨基酸[71]。
泌乳母猪产奶量随饲粮蛋白质摄入量的增加而增加,饲喂含18%粗蛋白质的基础饲粮的泌乳母猪在泌乳的第1~21天内每天损失约0.5 kg体重的蛋白质,表明饲粮蛋白质的摄入量对于多产母猪的乳蛋白生产来说是严重不足的,其中进入门静脉的膳食Glu只能满足母猪乳汁中Glu产量的8%[72],因此泌乳母猪可能无法合成足够的Glu来支持最大产奶量[73]。Rezaei等[74]在29 d的哺乳期内给泌乳母猪饲粮中补充1%~2% MSG,发现母猪乳汁中游离氨基酸及蛋白质的含量显著提高,其哺乳仔猪采食量分别增加了14%和25%,而体重分别增加了27%和42%,并显著降低了仔猪的死亡率,表明饲粮中添加1%~2%的MSG可以提高泌乳母猪的产奶量并促进所饲喂仔猪的生长。
综上所述,饲粮中补充Glu有助于提高生猪的生长性能,提高蛋白质利用效率并降低氮排放;Glu在降低生猪背膘厚、提高IMF沉积以及促进肌肉蛋白质合成上具有重要作用;仔猪的肠道功能与肠道健康对其生长效率有着重要影响,进一步在生产上的应用发现Glu在改善断奶仔猪与生长猪的肠道功能上也发挥着积极作用;Glu还可促进妊娠母猪胎儿的生长、提高产奶量、改善母猪繁殖性能,提示Glu在提高猪生产效率及肉品质上具有良好的应用前景,但具体的作用机制有待进一步研究。
3.2 Glu在家禽生产中的应用
Glu对家禽的生长性能具有促进作用。Maruyama等[75]研究发现,在1~14日龄肉仔鸡基础饲粮中补充5%~10% Glu可显著促进肉仔鸡的体重增加,且在无Glu的基础饲粮中添加10% Glu可使其日增重增加4倍,显著提高肉仔鸡的生长性能。Bezerra等[76]研究发现,与饲喂基础饲粮的对照组相比,降低1日龄雄性肉鸡基础饲粮中粗蛋白质含量会显著降低其生长性能,但在降低粗蛋白质含量的饲粮中补充1%~2% Glu则对生长性能无显著影响,提示在低蛋白质饲粮中补充Glu对维持肉鸡生长性能是可行的。Shakeri等[77]研究发现,在1~21日龄肉仔鸡基础饲粮中补充0.5%的AG补充剂可显著降低肉仔鸡的死亡率而对生长性能无显著影响;另一研究发现,在1~21日龄肉仔鸡基础饲粮中补充1%的AG补充剂,可显著提高22~42日龄肉仔鸡的体重增加与饲料转化率,并可显著降低肉仔鸡的死亡率,提高生长性能,而这可能与肉仔鸡消化系统的发育完善相关[78]。另有研究发现,在刚孵化的雏鸡低蛋白质饲粮中补充Glu与甘氨酸(glycine,Gly)混合物有助于提高雏鸡的体重和饲料转化率,并降低腹部脂肪的沉积,提高雏鸡的生长性能[79]。马现永等[80]的研究发现,在42日龄黄羽肉鸡饲粮中添加540 mg/kg MSG对其生长性能无负面影响,且可通过提高肌肉中风味氨基酸的含量及促进风味物质形成相关基因的表达提高肉的风味,进而提高肉品质。以上研究结果表明,在饲粮中添加Glu有助于降低肉鸡的死亡率并提高生产效率,还可通过促进风味物质形成以提高肉品质。
产蛋效率及蛋品质是影响蛋鸡的生产效率的重要因素[81]。饲粮补充NCG可促进蛋鸡卵泡发育,并提高蛋鸡的采食量与饲料转化效率[82],还可显著提高产蛋后期蛋鸡的产蛋效率[83]及蛋壳质量[84]。以上研究结果提示,Glu及其代谢产物可促进卵泡发育,提高产蛋效率及蛋品质,在改善蛋鸡生产性能上有着潜在的积极影响。
Glu在提高家禽的生产效率及改善畜产品品质上也具有潜在的应用前景,但Glu对不同阶段家禽的生产性能作用如何仍有待进一步探究,Glu提高家禽的畜产品品质的作用机制仍有待进一步研究。
3.3 Glu在反刍动物生产中的应用
瘤胃是反刍动物最重要的消化器官之一,瘤胃微生物可用含氮化合物合成菌体蛋白以供机体利用,因此反刍动物饲粮中一般不需要补充氨基酸[85]。但热应激条件下,反刍动物饲粮摄入量、养分的吸收与利用能力下降,导致蛋白质的可用性下降[86],使得生长性能降低[87]。Li等[88]的研究发现,在湖羊饲粮中以3 g/(头·d)的水平补充Glu,饲养90 d后改善了热应激湖羊的瘤胃发酵情况,提高了营养物质消化率和代谢率,增强了免疫力,提高了湖羊的生长性能,提示长期补充Glu有助于提高热应激湖羊的生产效率。这表明补充Glu在缓解反刍动物热应激导致的生长性能下降上具有一定的应用前景。
瘤胃微生物对反刍动物的生长、健康水平等有着重要影响[89]。有研究发现,体外培养瘤胃微生物时补充Glu可显著促进其生长,而缺乏Glu则会显著降低瘤胃微生物的生长和效率[90]。进一步研究发现,成年牛及绵羊的瘤胃混合微生物对细胞外Glu的代谢能力有限[91],而Glu在分离的肉牛瘤胃上皮细胞和十二指肠黏膜细胞的三羧酸循环中产生α-酮戊二酸方面发挥着至关重要的作用[92]。作为小肠的重要能量代谢底物及谷胱甘肽的合成原料,Glu可添加于饲粮中以改善反刍动物的肠道功能[93],如另一项研究发现,在奶牛围产期饲粮中添加过瘤胃保护MSG可提高奶牛的消化能力和采食量,并在不影响产奶量的情况下,在产仔后可减少奶牛机体肌肉及脂肪的动员[94]。这提示添加过瘤胃保护的Glu对改善反刍动物的肠道功能并提高生产性能具有潜在的积极作用。
繁殖性能与产奶性能是反刍动物重要的生产性状[95]。饲粮中补充NCG可提高母羊早期胚胎数量与存活率[96]并增加产羔数与活羔数[97]。经产母羊饲粮中补充NCG可提高产双羔率并提高羔羊的生长性能[98]。饲粮中补充NCG可提高奶牛产奶量并改善乳品质[99],还可提高奶牛的泌乳性能及抗高原反应能力[100-101],并通过增加瘤胃微生物多样性以提高生产性能[102],围产期奶牛饲粮补充NCG可显著提高犊牛初生重,改善奶牛繁殖性能[103]。以上研究结果提示,Glu及其代谢产物能够提高胚胎存活率、产仔数及泌乳性能,对提高反刍动物生产性能有着一定的积极影响。
Glu在促进反刍动物瘤胃微生物生长、缓解热应激导致的反刍动物生产性能下降、改善反刍动物肠道功能、提高反刍动物繁殖性能上具有一定的生产应用前景,但具体的Glu改善反刍动物瘤胃发酵、提高生产性能与改善肠道功能的作用机制还有待深入研究。
4 小 结
综上所述,Glu在介导兴奋性突触传递、维持动物肠道健康、调节机体能量稳态、参与蛋白质与氨基酸的代谢等过程中发挥着重要作用,在畜禽饲粮中添加一定的Glu可提高畜禽体重、日增重和饲料转化效率,具有改善肉品质并提高畜禽繁殖性能等作用。但Glu的不同添加形式、添加浓度和饲喂时间对畜禽生产性能、产品品质以及肠道微生态等的影响还有待进一步研究;在实际生产应用过程中,不同畜禽品种间Glu的作用效果及机制是否存在差异以及Glu与其他氨基酸、营养素之间的互作效应也有待于系统研究;而Glu对畜禽生产性能和产品品质的调控机制也仍需进一步探究。随着更多研究的深入开展,Glu在畜禽生产中的功能及作用机制将得到进一步揭示和阐明,并将在畜禽健康养殖中得到更为广泛的应用。
