Lineaperpetua gen.nov.: a new diatom genus in the Thalassiosirales supported by morphology and molecular data*
2024-02-27PanYULinYANGQingminYOUJohnPatrickKOCIOLEKKangyuWANGYonghongBIQuanxiWANG
Pan YU, Lin YANG, Qingmin YOU, John Patrick KOCIOLEK, Kangyu WANG,Yonghong BI, Quanxi WANG,**
1 College of Life Sciences, Shanghai Normal University, Shanghai 200234, China
2 Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China
3 Museum of Natural History and Department of Ecology and Evolutionary Biology, University of Colorado, UCB 218, Boulder CO80309, USA
Abstract Based on a combination of morphology and molecular data of ribosomal DNA genes, a new diatom genus Lineaperpetua gen.nov.Yu, You, Kociolek & Wang is described.The features that help define Lineaperpetua at the level of genus include: a tangentially undulated valve face; continuous cribra areolae on the valve interior consisting of pores arranged as strips; single rimoportula located inside the ring of marginal fultoportulae.Additionally, phylogenetic analysis based on nuclear small subunit (SSU)rDNA sequences and nuclear large subunit (LSU) rDNA gene placed the three strains of L.lacustris in a single, monophyletic clade at a considerable sequence distance from the other genera (Thalassiosira,Conticribra, Planktoniella, Shinodiscus, and other genera) belonging to Thalassiosirales.Despite the similarities with some species of Thalassiosira, Conticribra, and Spicaticribra, the suite of features found in Lineaperpetua differentiate it from these other genera.These molecular data and morphological characters suggest an affinity of the new genus to the Thalassiosiraceae.
Keyword: diatom; morphology; new genus; phylogeny; taxonomy; Thalassiosiraceae
1 INTRODUCTION
The Thalassiosirales including roughly 500 species,and belongs to roughly 30 genera (Niu et al., 2022),and these genera are usually distinguished by morphological features.Although there have been various schemes to organize the diversity in the group, the diatoms that possess fultoportulae were identified as a natural group (Theriot and Serieyssol,1994), and they have been recognized as the Thalassiosirales.This interpretation agrees with phylogenetic analyses based on molecular data(Medlin and Kaczmarska, 2004; Niu et al., 2022).The Thalassiosirales have been shown to be comprised of four families, the Lauderiaceae, Skeletonemataceae,Thalassiosiraceae, and Trochosiraceae (Glezer and Makarova, 1986; Round et al., 1990; Kaczmarska et al., 2006).The former family broadly includes some freshwater genera (such asContricribra) and the latter one mostly marine taxa (includingThalassiosira,Skeletonema,Detonula, etc.), though both groups have species that ‘leaked’ in estuarine and even freshwater environments.
The diatom genusThalassiosirawas established by Cleve for those radially symmetrical diatoms which have specialized pores (now known as larger rimoportula and small fultoportulae) on the valve face (Round et al., 1990).The genus was emended by Hasle (1973a), noting the processes fultoportulae on the valve face and mantle.In addition, in this genus, most species have only one rimoportulae(Niu et al., 2022).These features helped to define the genusThalassiosira, but also a broader group of diatoms that include marine, estuarine, and freshwater taxa (Hasle, 1973b, 1978; Fryxell, 1975; Fryxell and Hasle, 1979a, b).
Attempts to understand the diversity withinThalassiosirayielded descriptions of many new taxa(e.g., Fryxell and Hasle, 1972; Fryxell, 1975; Hasle,1985), and better details on the morphological breadth among its representatives.For instance, a group ofThalassiosiraspecies (both in Recent and fossil, mostly freshwater environments), was shown to have tangentially undulate (‘plicate’) valve faces, leading Julius and Tanimura (2001) to produce a phylogeny of these taxa.While manyThalassiosiraspecies have cribra over individual internal areolar openings (which may be dome-like or flat, e.g., Hasle, 1978; Fryxell and Hasle, 1979b),others have a covering spread over the entire valve interior (Hasle and Fryxell, 1977; Kociolek and Khursevich, 2001; Prasad et al., 2011).These dichotomies in morphology suggested to some they might be divided into separate genera (Prasad et al.,2011).Makarova (1994) described new genusRoundiabased onThalassiosiracardiophora, it having indistinct areolae on the internal openings, internal tubes short and very small basal flanges of the fultoportulae, outward extensions of the fultoportulae that have five wings and as well as a group of slightly large exit tubes curved inwards towards the valve center.Makarova (1994) described new genusTakanoabased onThalassiosirabingensis, which has indistinct areolae on the internal openings.Alverson et al.(2006) proposed the genusShionodiscus,for those species formerly inThalassiosirabut having rimoportulae on the valve face and short inward and reduced outward extensions of the fultoportulae.Genera with the coverings across the openings in the valve interior have been established,includingConticribra(Stachura-Suchoples and Williams, 2009) andSpicaticribra(Johansen et al.,2008).
Phylogenetic analysis of the Thalassiosirales using nuclear (SSU rDNA and partial LSU rDNA)and chloroplast (rbcL and partialpsbC) markers by previous studies (Kaczmarska et al., 2006; Alverson et al., 2007; Theriot et al., 2010; Li et al., 2018,2020a, b; Downey et al., 2021) showed that both the Stephanodiscaceae and Thalassiosiraceae were nonmonophyletic, and that ecological boundaries related to salinity (thought to help diagnose these groups) had been crossed numerous times by taxa in the Order.In the case of the Stephanodiscaceae, the genusCyclotellaas typified byC.distinguendais shown more closely related to species ofThalassiosirathan part of a monophyletic lineage of other Stephanodiscaceae genera and species.This same analysis showed thatThalassiosira, as typified byT.nordenskioeldii, was non-monophyletic, with species previously assigned to the genus spread across the Thalassiosirales.Alverson et al.(2007) suggested their work, however, “… offer[s] little towards the taxonomic uncertainties surrounding the large and vaguely defined genusThalassiosira”.
In the present report, we provide sequence data for 3 strains of one of the plicated, freshwaterLineaperpetuaspecies,L.lacustris, and include it in the overall phylogeny of the Thalassiosirales.Based on its phylogenetic position we propose to assign it to a new genus,Lineaperpetua, and discuss the further possible taxonomic approaches to the taxa currently assigned toThalassiosira.
2 MATERIAL AND METHOD
2.1 Sample collection, isolation, and culture
Diatom samples were collected from middle reaches of the Changjiang (Yangtze) River, China, in April 2021, including Qingjing River (30°24′12″N,111°24′48″E) on Yidu City, Hubei, and Yunyang Pier (30°56′4″N, 108°41′2″E) on Yunyang County,Chongqing.They were collected using toothbrushes from the natural substrates.Single diatom cells were isolated using a Pasteur pipette (Yang et al., 2022).Cells were isolated and cultured in 24-well cell plates and each well contained 2-mL CSI medium.Nonaxenic unialgal cultures were maintained in CSI medium at 24 ℃ in a growth chamber under a 12-h:12-h light/dark photoperiod (Yang et al., 2022; Yu et al., 2022).Nucleotide sequences of SSU rDNA and LSU rDNA of clone CJ2D3, QJ, and CJ deposited in GenBank (NCBI), with accession numbers OM906790,OM906791, OM906787, OM906792, OM906789,and OM906793, respectively.
2.2 Light microscopy (LM) and scanning electron microscopy (SEM) analysis
Samples were digested using concentrated nitric acid (Parr et al., 2004; Yu et al., 2022), and modified as described in Yu et al.(2019).The cleaned diatoms were then mounted in Naphrax®to obtain permanent slides for the LM analysis (Yang et al.,2022).LM examinations were performed with a 100× objective lens (1.4 numerical aperture) using a ZEISS AXIO Imager A2 microscope (Yu et al.,2022).SEM examination was made using a Hitachi SU 8010 SEM (2 kV) at Shanghai Normal University,Shanghai, China (Yang et al., 2022).Images were compiled with Adobe Photoshop CS6 (Adobe Photoshop CS6 Extended).All diatom samples and permanent slides are housed in the Biology Department Diatom Herbarium, Shanghai Normal University (SHTU).
2.3 Phylogenetic analysis
DNA extraction was performed as described in Yang et al.(2022).The method for amplifying fragments of SSU rDNA (~1 750 nt) and LSU rDNA(~600 nt) followed Yu et al.(2022).Newly determined sequences and GenBank sequences(Supplementary Table S1) of 67 separate centric diatoms were used in this study.Bellerochea malleuswas selected as the outgroup taxon to root the trees.Sequences were aligned using the Clustal W option in the BioEdit sequence analysis software(Thompson et al., 1997; Hall, 1999; Yang et al.,2022).The ModelTest v3.7 Software was used to choose the most appropriate evolutionary model(Posada, 2006) (Table 1).Finally, Sequence Matrix was constructed using SSU rDNA and LSU rDNA sequences (Vaidya et al., 2011; Yang et al., 2022).PHYML software was used to generate Maximum likelihood (RAxML) trees and the bootstrap analysis was conducted using 1 000 replicates (Felsenstein,1981; Guindon and Gascuel, 2003).Bayesian analyses(BA) were conducted using MrBayes v3.1.2 (Ronquist and Huelsenbeck, 2003).The resulting phylogenetic trees were edited using FigTree v1.4.2 and Adobe Illustrator CS6.
3 RESULT
3.1 Morphological analysis
LM observations (Fig.1a-h) show valves are round, tangentially undulate, with coarse radial striation composed of single rows of large areolae.Valve diameter ranges from 11.5 to 39.5 μm (n=50).One marginal ring of fultoportulae is regularly spaced on the valve mantle, and an inner ring oftubular, irregularly-spaced processes (occluded processes) are present.Valve face fultoportulae are not evident among the coarse areolae in LM.One rimoportula is present slightly inside the outer ring of marginal fultoportulae.
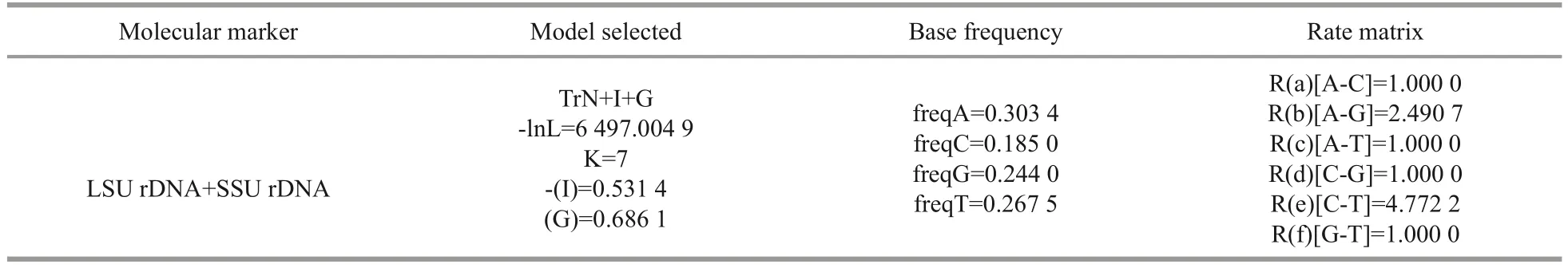
Table 1 Parameters of the nucleotide substitution model estimates using Modeltest v3.7
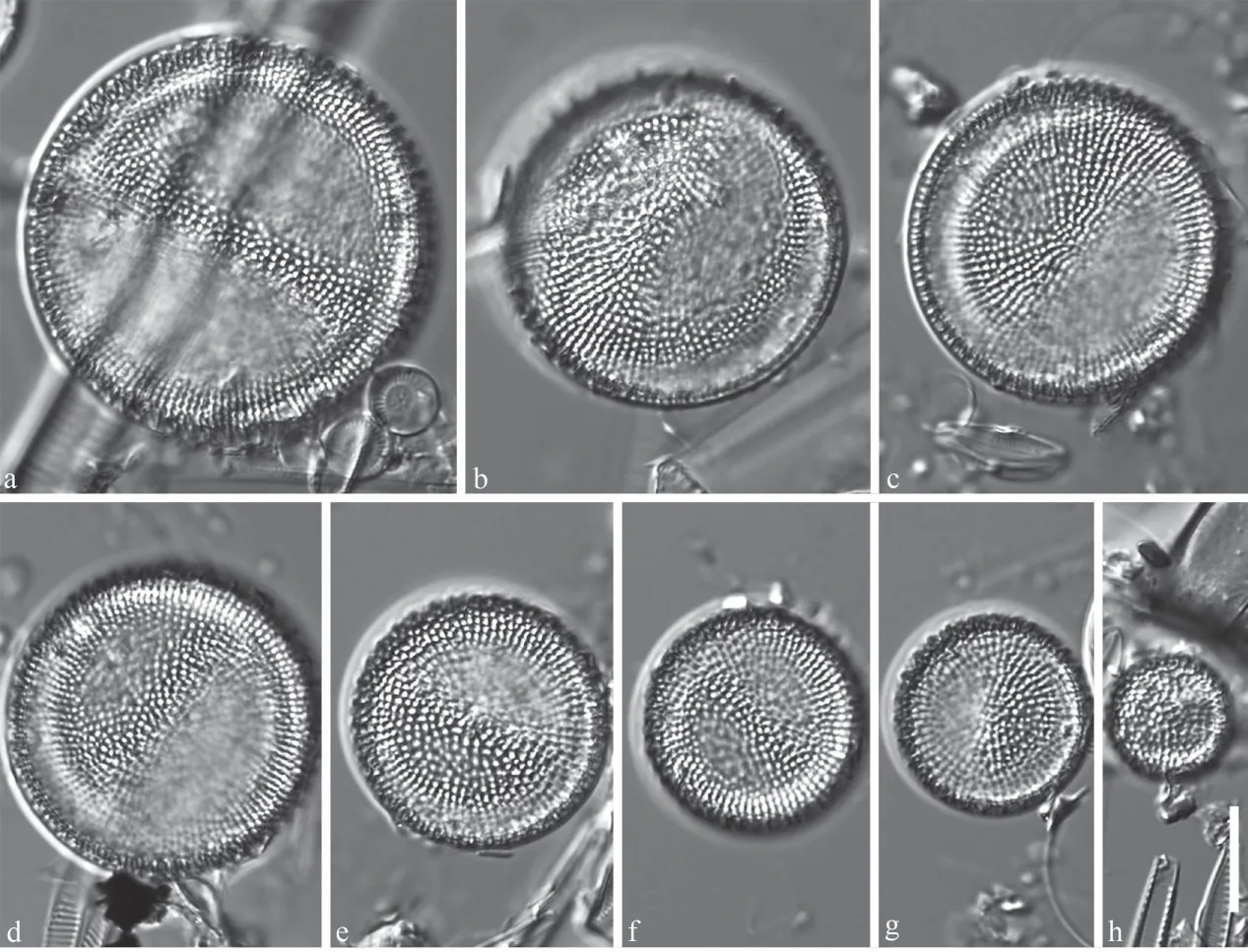
Fig.1 LM valve views of Lineaperpetua lacustris
In the SEM the valve exterior (Figs.2-5) has areolae, open to the outside by irregularly circular foramina that are radially arranged (Fig.2a-b), and thin silica granules on the surface of the areolae(Fig.2a-f).Areolae are composed of complex frameworks, composed of at least two layers(Fig.2e-f).The marginal ring of fultoportulae is located on the fairly steep mantle.They number of marginal fultoportulae from 4 to 7/10 μm(Figs.2a-b, 3a-d, 4a-b, & 5a).The external tubes of the marginal fultoportulae have long tubular extensions and are surrounded by several thin siliceous wrinkles and thin silica granules (Fig.2c-d).One ring of occluded processes is located on the border toward the flattened part of the valve.The number of occluded processes was approximately half than that of marginal fultoportulae (Fig.2a-b).The external tubes of the occluded processes have thin siliceous wrinkles and thin silica granules and appear to have narrow tube-like cavities (Fig.2c-d).The ring of valve face fultoportulae is arranged in an irregular circle and located in the nearly the half between the valve center and the margin (Fig.2a-b).The valve face fultoportulae are shorter than the marginal fultoportulae (Fig.2e-f).A prominent rimoportula was present on the valve face, close to the margin and its external tube is longer and larger than that of fultoportulae (Fig.2a-b).

Fig.2 Lineaperpetua lacustris SEM, external valve view
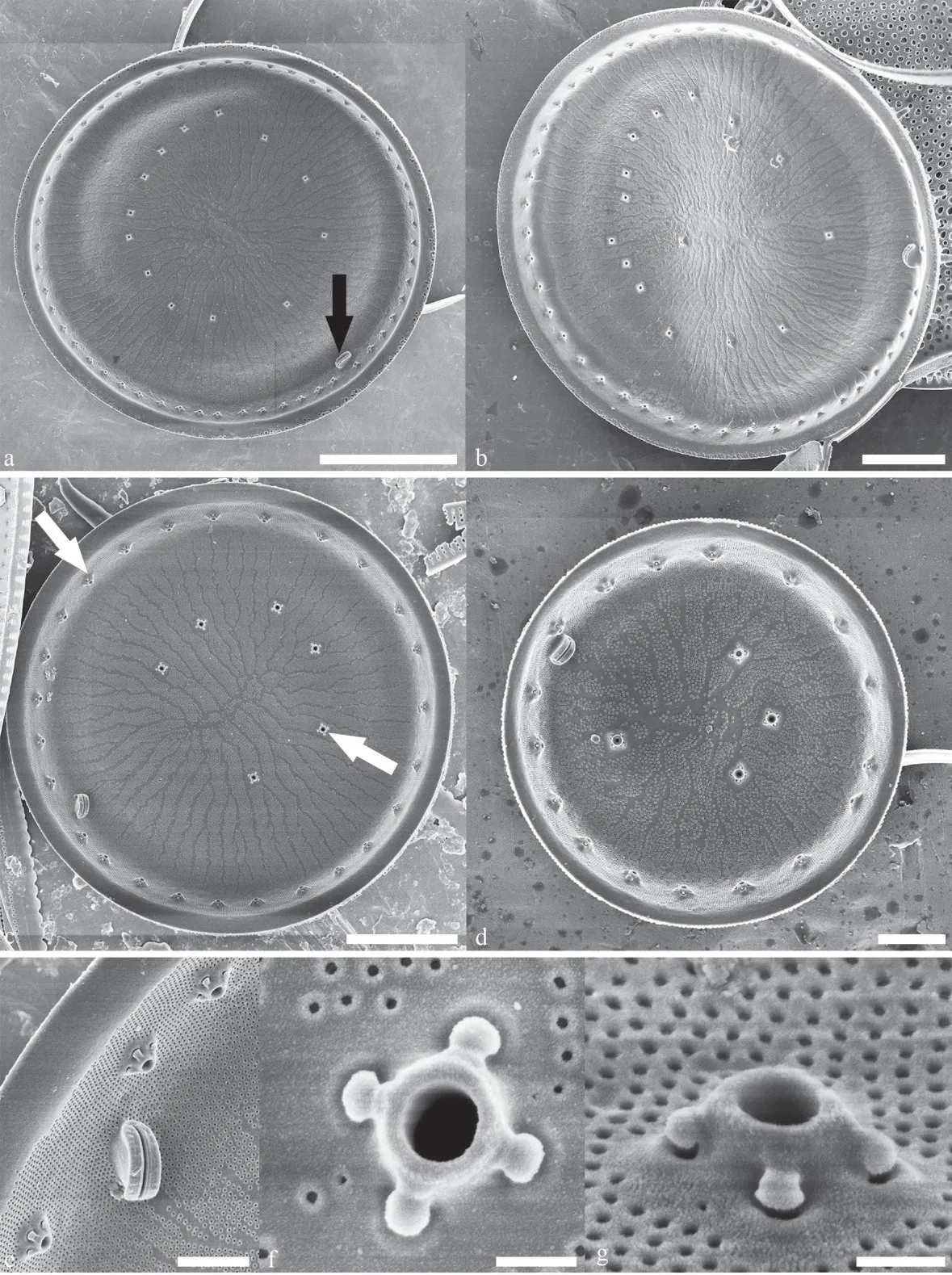
Fig.3 Lineaperpetua lacustris SEM, internal valve view

Fig.4 Lineaperpetua lacustris SEM, whole frustule in girdle view
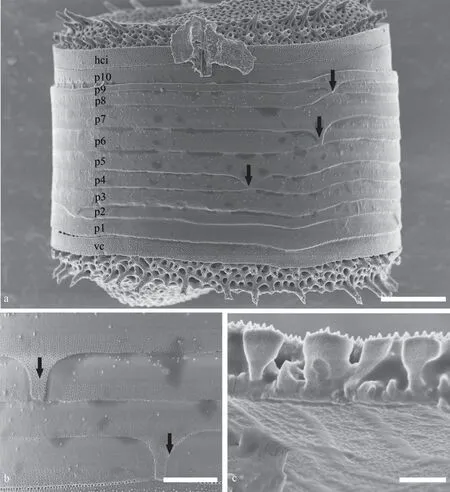
Fig.5 Lineaperpetua lacustris, SEM, the fine structure of girdle
Internally, the cribra are continuous, consisting of radial strips with individual pores evident, and the pattern center is elongated with many interstriae arising perpendicular to the long axis and annules(Fig.3a-d).Continuous cribra were in radial anastomosing patterns which continued onto the valve mantle (Fig.3d-e).The marginal fultoportulae and valve face fultoportulae both have four satellite pores and satellite pore covers (Fig.3f-g).A single rimoportula is located towards the valve center of the open part of the semicircle of valve face fultoportulae.The rimoportula is thick-lipped, shortstalked, slightly curved, with a slit that is parallel to the valve margin or oblique to the margin(Fig.3a-e).In an innermost layer, inner sides of the loculate areolae are covered by continuous cribra in the valve face (Fig.5c).
The frustules are drum-shaped and have a strong undulation on the valve face evident in girdle view(Figs.4a-b & 5a).At the base of the mantle the valve has a marginal thickened flat rim to which the valvocopula is attached, the cingulum is comprised of 7 to 12 open bands, and all bands have the openings in a dextral pattern (Figs.4a-b & 5a).The valvocopula has thin silica granules (Figs.4b & 5a).Moreover, the adjacent pleura band has fine pores with an antiligula in the advalvar part, and on the pleura part minute granules are lacking (Fig.5b).
3.2 Molecular analysis
77 centric diatom strains and three outgroup taxa was used in this study, including 1 113 characters, of which 344 (30.9%) were variable and 226 (20.3%)were parsimony-informative sites.There were 769(69.1%) conserved sites.The average percentages of T, C, A, and G were 26.8%, 18.5%, 30.3%, and 24.4%, respectively.ML and BI analyses for phylogenetic analysis was shown (Fig.6).The topology result of the phylogenetic trees obtained by both ML and BA was consistent.
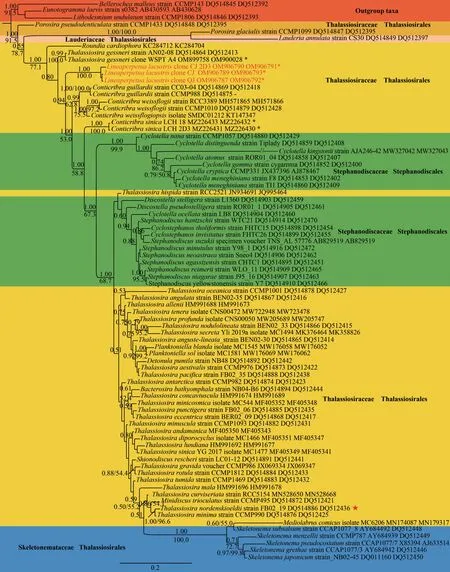
Fig.6 Bayesian tree of Lineaperpetua gen.nov.(in red) constructed from a concatenated alignment of 67 partial SSU rDNA and LSU rDNA sequences of 1 113 characters
All Thalassiosirales species were divided into three clades.Clade I included twoPorosiraspecies,andPorosiraglacialiswas sister to the genusLauderia; Clade Ⅱ contains the new genusLineaperpetua,Roundia,Thalassiosiragessneri,andConticribraspecies; in Clade Ⅲ, most species belong toThalassiosira, besides some genera(Planktoniella,Detonula,Bacterosira,Shionodiscus,Mediolabrus) inserted into this large clade.With this result, the Stephanodiscaceae are non-monophyletic.All of theConticribraspecies were found in two clades, so the genus is non-monophyletic.
Three strains were isolated and used in phylogenetic tree construction.The sequences of the three strains wereLineaperpetualacustriscomb.nov.(≡Thalassiosiralacustris).In the phylogenetic tree,Lineaperpetuawhich will be described as a new genus formed a single, well-supported(BPP/ML 1.00/100.0) branch within the big clade with genera belonging to Thalassiosiraceae,Stephanodiscaceae, and Skeletonemataceae.
The pairwise uncorrectedp-distances based on the partial SSU rDNA gene showed that threeLineaperpetualacustrisstrains had ap-distance of 0.00, indicating that they exhibited 100% similarity.However, they had an evolutionary distance of 0.02-0.10 with other Thalassiosirales species based on SSU rDNA gene.ThreeL.lacustrisstrains had ap-distance of 0.02 withThalassiosira,Conticribra,Shionodiscus,Planktoniella,Detonula,Minidiscus,andRoundia, suggesting a close genetic relations(Table 2).The pairwise uncorrectedp-distances based on the partial LSU rDNA gene showed that threeL.lacustrisstrains had ap-distance of 0.00-0.01, indicating that they exhibited 99%-100%similarity.Similarly, threeL.lacustrisstrains had closer genetic relations withThalassiosira,Conticribra,Planktoniella,Detonula, andMinidiscus, and it had ap-distance with those genera (Table 3).
Evidence from morphology and molecules suggest thatLineaperpetualacustrisforms a unique monophyletic group, well separated fromThalassiosiraas typified byT.nordenskioeldii.Based on thisfinding we propose a new genus forL.lacustris:

Table 2 Pairwise uncorrected p-distances of 20 strains based on the partial SSU rDNA gene

Table 3 Pairwise uncorrected p-distances of 20 strains based on the partial LSU rDNA gene
Class Bacillariophyceae Haeckel emend.Medlin and Kaczmarska (2004)
Subclass Thalassiosirophycidae Round & Crawford(Round et al., 1990)
Order Thalassiosirales Glezer & Makarova(Round et al., 1990)
Family Thalassiosiraceae Lebour (Round et al.,1990)
LineaperpetuaYu, You, Kociolek & Wang, gen.nov.
Description: Frustules drum-shaped.Valve circular, with a tangentially undulated (plicated)valve face.Striation composed by single rows of areolae are well seen.Areolae with a continuous cribra on the valve interior, and composed of strips arranged pores.Central area have valve face fultoportulae, and their number increases with the diameter of the frustule.Marginal fultoportulae arranged in regular ring, external opening as a long tube, internal opening slightly raised.Single rimoportula located inside the ring of margin fultoportulae, its external tube is longer and larger than that of fultoportulae.
Etymology: The name refers to the uninterrupted lines of cribra on the valve interior.
Type species of the genus:Lineaperpetua lacustris(Grunow) Yu, You, Kociolek & Wang comb.nov.
Lineaperpetualacustris(Grunow) Yu, You,Kociolek & Wang comb.nov.
Basionym:CoscinodiscuslacustrisGrunow in Cleve and Grunow (1880): Beiträge zur Kenntniss der arctischen Diatomeen.Kongliga Svenska Vetenskaps-Akademiens Handlingar 17(2): 114.
Homotypic synonym:Thalassiosiralacustris(Grunow) G.R.Hasle in Hasle and Lange (1989):Freshwater and brackish waterThalassiosira(Bacillariophyceae): taxa with tangentially undulated valves.Phycologia, 28: 120-135.
4 DISCUSSION
Lineaperpetuais characterized by the tangential undulation valve and strips of continuous cribra.Table 4 documents other genera that show some similar combinations of morphological features:ThalassiosiraCleve (Round et al., 1990),Cyclotella(Kützing) Brébisson (Downey et al., 2021), andConticribraStachura-Suchoples & Williams (Stachura-Suchoples and Williams, 2009).The valves ofLineaperpetuaare tangentially undulated (plicated),butCyclotellahas undulate or flat valve face,Conticribrahave flat valves, and taxa assigned toThalassiosiracan have predominantly plicated except for the central concavity.The cribra ofLineaperpetuais continuous and arranged in radial strips, while inCyclotella, the areolae are poroidal,closed internally by cribra or continuous.In addition, the cribra of mostThalassiosiraare individual, each with a flat to domed cribrum and are of equal size on the valve face and mantle, and inConticribrathey are (semi) continuous.Furthermore, the external tube of the marginal fultoportulae ofLineaperpetua,Thalassiosira, andConticribraare long tubular extensions, while inCyclotellathis extension is slightly raised.The internal marginal fultoportulae ofLineaperpetuaandConticribraare slightly raised with 4 satellite pores,while inThalassiosirathey are slightly raised with 2-5 satellite pores, and inCyclotellathey are long tubes with 2-3 satellite pores.Valve face fultoportulae are present in the nearly the half between the valve center and the margin inLineaperpetua, each with 4 satellite pores, but inCyclotellathey are absent or present each with 3 satellite pores, while inConticribrathey are present each with 2-4 satellite pores.Moreover, inThalassiosira, have one or more valve face fultoportulae positioned in the valve center.Moreover,LineaperpetuaandConticribrapossess one rimoportula, but rimoportula located within the ring of marginal fultoportulae ofLineaperpetua, while the rimoportula is located on the valve mantle that replaces a marginal fultoportulae ofConticribra, and the rimoportula is closer to the margin than the fultoportulae ofCyclotella, and the rimoportula is also located at the junction of the valve face and margin, but its precise position varies.In addition, other genera have only one ring of marginal fultoportulae and no occluded process, while the new genusLineaperpetuahas a ring of occluded processes.Whether this occluded processes can be used as the identification feature ofLineaperpetuastill needs more research to prove.At present, there is only one species ofLineaperpetua,and it is hoped that more species ofLineaperpetuacan be observed in the future to determine whether this feature is the generic combination ofLineaperpetua.
Khursevich and Kociolek (2012) and Gallo-Sánchez et al.(2015) proposed thatT.lacustrisshould be considered for transfer to the genusCyclotella(Spicaticribraspecies were recently transferred intoCyclotellaby Downey et al.(2021),because it has radial, zig-zag paths of continuouscribra on the internal surface).However,T.lacustris,when compared with those species, reveals some significant differences.First, the external tube of the marginal fultoportulae inT.lacustrisis a long tubular extension, and internally it is slightly raised by four satellite pores and satellite pore covers,while in ‘Spicaticribra’ species, marginal fultoportulae have slightly raised external and elongated internal extensions with 2-3 satellite pores with well developed ‘cowlings’.Second, the rimoportulae are located near the marginal fultoportulae in‘Spicaticribra’ species, but located inside the ring of margin fultoportulae inT.lacustris.Furthermore,compared withT.lacustris, in the external valve, the valve face areolae are usually smaller (3-6 times)than central areolae in ‘Spicaticribra’ species(Johansen et al., 2008; Stachura-Suchoples and Williams, 2009; Tuji et al., 2012).Finally, phylogenetic analysis shows thatT.lacustrisand ‘Spicaticribra’are in two independent branches that are separated from other species ofThalassiosira(see Fig.6 herein,also Alverson et al., 2006; Downey et al., 2021).Based on the combination of morphology and molecular data of ribosomal DNA genes,T.lacustrisshould be separated fromThalassiosiraas an independent genus, and this conclusion is also recognized by Prasad et al.(2011) and Julius and Tanimura (2001).
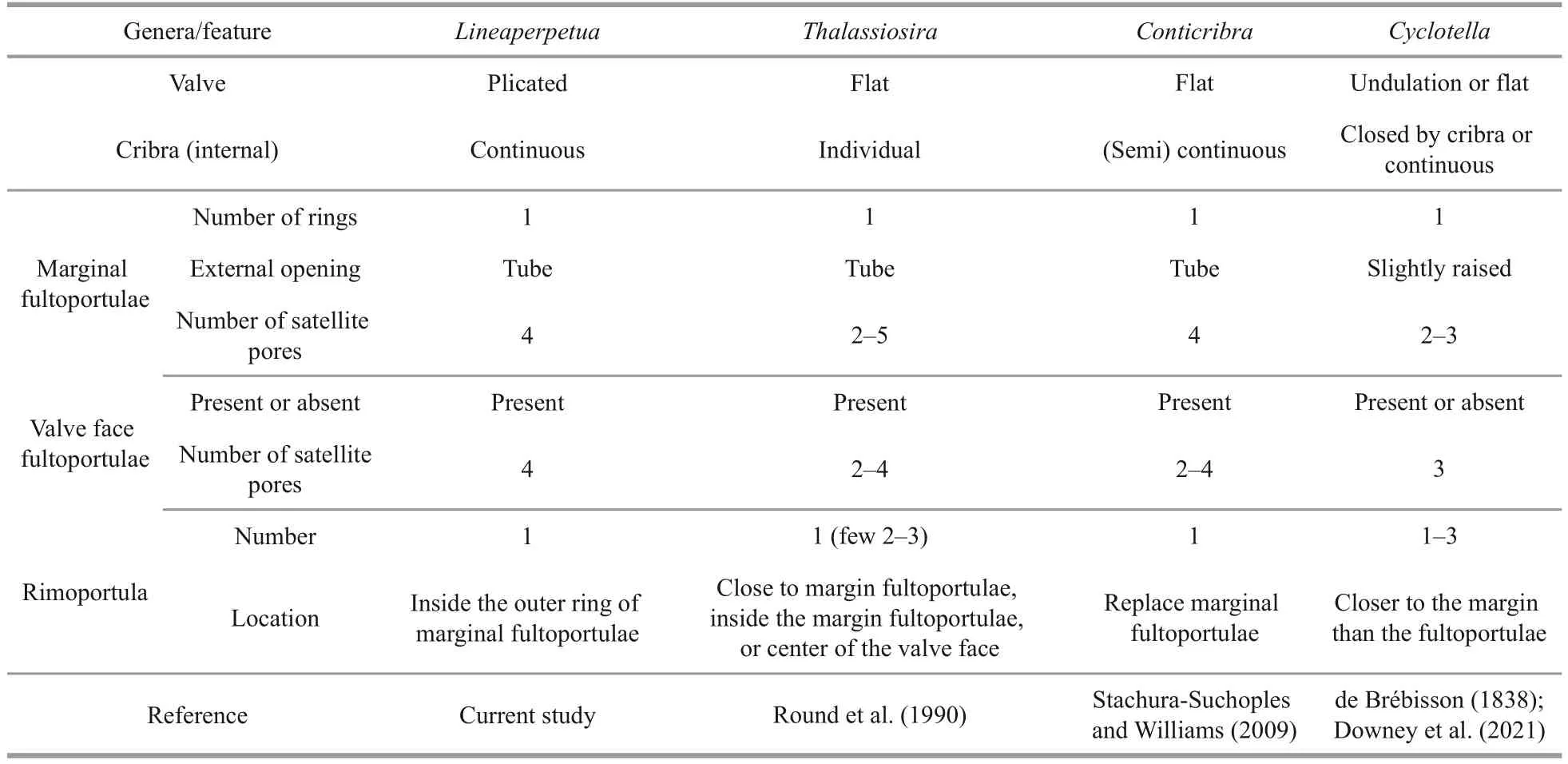
Table 4 Comparison of morphological characteristics of Lineaperpetua gen.nov.and other similar genera
Combining data on morphology and molecules,the concept ofThalassiosiraproposed by Hasle and Lange (1989) has been modified (Alverson et al.,2007).Species with strip-like continuous cribra,tangentially undulated (plicated) valve face, and rimoportula located in inside the ring of margin fultoportulae belong to the new genusLineaperpetua.Kociolek and Khursevich (2001)indicated thatT.kilarskiKaczmarska possess internal cribra openings, which are similar to those inLineaperpetualacustris, and this might suggest this fossil taxon may belong to this new genus.In addition, there are four species similar toL.lacustris, includingT.australiensis(Grunow) Hasle and Lange (Hasle and Lange, 1989),T.bramaputrae(Ehrenberg) Håkansson & Locker (Håkansson and Locker, 1981),T.livingstoniorumPrasad, Hargraves and Nienow (Prasad et al., 2011), andT.gessneriHustedt (Hustedt, 1956) that might be included in the new genus.However, due to the lack of detailed information on the characteristics such as cribra pattern on the inner valve and the position and shape of rimoportula, it is uncertain whether they should be transferred to the new genus at this time.Future research is necessary to ascertain their systematic placement.Moreover, in our study, althoughT.gessnerihas tangentially undulated (plicated) valves and continuous cribra on the valve interior, it differs fromLineaperpetuain that the rimoportula ofT.gessneriis between the two marginal fultoportulae,andT.gessnerihave the much finer areolae and the larger rimoportula than inLineaperpetua; the fultoportulae are operculate but differed fromLineaperpetuain that the marginal processes usually have five and those on the valve face have four satellite pores, and the rimoportula is short-necked,somewhat twisted, more elongated and compressed with narrower lips than inLineaperpetua.In addition, the external tubes of the marginal fultoportulae are surrounded by several thin siliceous wrinkles and thin silica granules inLineaperpetua, but smooth inT.gessneri(Hasle and Lange, 1989; Kiss et al., 2002).In our phylogenetic analysis,T.gessneriis clustered in an independent branch separate from bothLineaperpetuaandThalassiosira(Alverson et al., 2007; Downey et al.,2021).
There are several subspecific taxa ofL.lacustrisworthy of review as well.They includeThalassiosiralacustrisvar.cassiospinuaCai and Xie (Xie and Cai, 1981),T.lacustrisvar.hyperborea(Grunow) Qi (Qi et al., 1980),T.lacustrisvar.nipponica(Skvortzow) Qi (Qi et al., 1980),T.lacustrisvar.proximus(Makarova) Makarova(Makarova, 1983),T.lacustrisvar.septentrionalis(Grunow) Qi (Qi et al., 1980), andT.lacustrisvar.sinica(Skvortsov) Qi (Qi et al.1980).Due to the loss of type material and insufficient available illustration to accurately diagnose these taxon, more work is needed to re-collect samples from the type localities of these taxa and re-evaluate their taxonomic placement.Therefore, we suggest these taxa remain inThalassiosiraeven though the nominate variety is included in the new genus, until these taxa are confirmed as separate and can be placed within a monophyleticLineaperpetua.
In this study, most of the sequences for taxa in the Thalassiosirales were obtained from GenBank.Nuclear SSU rDNA sequences and LSU rDNA sequences were applied in the phylogenetic analysis of centric diatoms of Thalassiosirales.The result show that taxa belonging to Thalassiosirales are clustered into several large clades, which are separated from other genera of the order(Kaczmarska et al., 2006; Alverson et al., 2007; Li et al., 2018, 2020a; this study).The results of these previous analyses and in this current work show that the genusThalassiosirais non-monophyletic.Phylogenetic trees based on SSU rDNA and LSU rDNA sequences showed thatLineaperpetuais monophyletic group in the Thalassiosirales.Differences in both morphological features and molecular analyses support recognizingLineaperpetuaas a new genus.Meanwhile our molecular investigation also showed thatThalassiosira gessneriis clustered in a single branch but separated from other mostThalassiosiraspecies, and its phylogenetic data are congruent with results by previous studies (Alverson et al., 2007; Park and Lee, 2014; Li et al., 2018, 2020b; Niu et al., 2022).Based on these results, it seems reasonable to suggest thatT.gessneriis separated from the genusThalassiosira.T.gessnericould be a new genus, or a member ofLineaeperpetua, and further studies are needed for a solid conclusion.
We can also use the results of this analysis to examine the possible character evolution of features described within the Thalassiosirales.For example,the tree topology presented here suggests that tangentially undulate valves evolved at the base of tree, and may be diagnostic for the entire order, and that they were subsequently lost and then regained elsewhere in the large clade that includes most of the species currently assigned toThalassiosira.Thus, the ‘plicated Thalassiosiraceae’ do not form a monophyletic group as assumed by Julius and Tanimura (1991).Also, the feature of internal cribra cover types, both domed or flat over individual areolae and strip-like, appear to be within the Thalassiosirales.
Finally, treatment of the large clade containing most of the species currently assigned toThalassiosirarequires further review.It would be possible to treat the group as a single large genus.Within this clade are several monophyletic groups.One of these groups includes the type species ofThalassiosira,T.nordenskioeldii, and this group could represent a narrower interpretation of the genus.Another clade within this group contains species that have been assigned to the genusShionodiscusAlverson et al.(2006).In their subsequent analysis, Alverson et al.(2007) retained species within a broad concept ofThalassiosira.However, to acceptShionodiscuscreates the situation where the genusThalassiosirais nonmonophyletic.Finally, a third monophyletic group,consisting ofT.concaviusculathroughT.aestivalis,may require a name separate fromThalassiosiraat the rank of genus.Keeping these taxa withinThalassiosirarendersThalassiosiranon-monophletic if taxa such asShionodiscusare retained.
The results presented herein are largely aligned with the results of Alverson et al.(2007) and Downey et al.(2021).Alverson et al.(2007)’s conclusion that their work “ … offer[s] little towards the taxonomic uncertainties surrounding the large and vaguely defined genusThalassiosira”appears in hindsight to be too humble.It is our hope that with the proposal here of a new genus forT.lacustris, addressing nomenclatural ramifications of the consistently identified monophyletic groups(now recovered for over a decade) of taxa previously part ofThalassiosiracan progress forward.
5 DATA AVAILABILITY STATEMENT
All data of this study can be obtained from the corresponding author.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Contrasts of bimodal tropical instability waves (TIWs)-induced wind stress perturbations in the Pacific Ocean among observations, ocean models, and coupled climate models*
- Variability of the Pacific subtropical cells under global warming in CMIP6 models*
- Identification of thermal front dynamics in the northern Malacca Strait using ROMS 3D-model*
- Magmatic-tectonic response of the South China Craton to the Paleo-Pacific subduction during the Triassic: a new viewpoint based on Well NK-1*
- An improved positioning model of deep-seafloor datum point at large incidence angle*
- Microplastics in sediment of the Three Gorges Reservoir:abundance and characteristics under different environmental conditions*