Neomicrocoleus gen.nov.(Microcoleaceae, Oscillatoriales), a novel cyanobacterial genus from benthic mats in a water channel*
2024-02-27RuozhenGENGZhongshiHEKaihuiGAOPengXIAOHeZHANGSiCHENHuaLIGongliangYURenhuiLI
Ruozhen GENG, Zhongshi HE, Kaihui GAO, Peng XIAO, He ZHANG, Si CHEN,Hua LI, Gongliang YU, Renhui LI,**
1 College of Life and Environmental Sciences, Wenzhou University, Wenzhou 325035, China
2 Key Laboratory of Algal Biology, Institute of Hydrobiology, Chinese Academy of Sciences, Wuhan 430072, China 3 University of Chinese Academy of Sciences, Beijing 100049, China
4 Hubei Institution of Land Surveying and Mapping, Wuhan 430072, China
Abstract A new filamentous cyanobacterial strain (CHAB 4127) was successfully isolated from the algal mat during the field investigation of the open channel from Luanhe River to Tianjin, China.The polyphasic approach combining morphological, ultrastructural, ecological, and molecular features was used to characterize this studied strain.The strain is morphologically similar to the Microcoleus-like cyanobacterial taxa under light microscopy, and the radial arrangement of thylakoids is also consistent with that of Microcoleus-like groups.The phylogenetic position of CHAB 4127 based on 16S rRNA gene sequences, is shown to be clearly clustered into an independent clade with the newly established genus Microcoleusiopsis.The maximum similarity of 16S rRNA gene of the studied strain with other existing related cyanobacterial taxa is 93.97%, and the ITS secondary structures is also obviously different from other members of Microcoleaceae.Based on all the above evidence, we proposed the establishment of this novel cyanobacterial genus as Neomicrocoleus, with its type species, Neomicrocoleus tianjinensis.
Keyword: Neomicrocoleus tianjinensis; morphology; 16S rRNA; taxonomy; polyphasic approach
1 INTRODUCTION
Cyanobacteria are the oldest photoautotrophic organisms on earth (Schopf, 2011).As the main contributor to the early aerobic environment on the Earth, cyanobacteria strongly influenced the evolution of the earth’s environment and life systems(Rasmussen et al., 2008).The cyanobacterial taxonomic system has been officially established for less than a century since its inception (Geitler,1925b, 1932).However, the lengthy and complex evolutionary history of cyanobacteria makes it difficult to distinguish them based on morphological characteristics alone.The polyphasic approach combining morphological, ultrastructural, ecological and molecular (16S rRNA gene, 16S-23S ITS region and genome) information has gradually become the most suitable method to characterize cyanobacterial taxa and conduct their taxonomic studies (Comte et al., 2007; Sciuto et al., 2011;Komárek, 2018).In the past decades, with the description of a large number of novel cyanobacterial genera and species using the polyphasic approach,the taxonomic system of cyanobacteria has undergone considerable revisions (Komárek et al., 2014, 2020;Mareš, 2018).
The genusMicrocoleusDesmazières ex Gomont 1892, now belonging to Microcoleaceae in Oscillatoriales, was established with type speciesMicrocoleusvaginatusGomont 1892 by Gomont(Gomont, 1892).Based on AlgaeBase Database(www.algaebase.org, accessed on 9 November 2022), 137 species ofMicrocoleushave been recorded, of which 55 species have been officially taxonomically accepted.The typical morphological features of this genus are that filaments are long,solitary, forming thin strata or mats, covered by gelatinous, hyaline, colorless and homogeneous sheaths, and each sheath usually packs with numbers of trichomes (ranging from 2-3 to 100)(Boyer et al., 2002).The species ofMicrocoleusare mainly benthic and soil species, and have a wide range of ecological niches in various ecosystems.Some species have strong adaptability to extreme environments such as deserts and droughts, and several species can also survive in estuaries, mineral springs, sea coasts and inland saline habitats (Manen and Falquet, 2002; Strunecký et al., 2013, 2014;Engene et al., 2018; Heidari et al., 2018).Additionally,some species ofMicrocoleushave the ability to synthesize and release secondary metabolites that are harmful to humans, animals and plants.They can not only produce pungent taste and odor (T & O)substances such as geosmin or/ and 2-Methylisoborneol(2-MIB), but also produce cyanotoxins like microcystins (MCs), neurotoxic anatoxin-a (ATX)and homoanatoxin-a (HATX), thus threatening human health and water ecosystem security (Bouma-Gregson et al., 2019; Niiyama and Tuji, 2019;Conklin et al., 2020).
As the most diverse and complicated genus in Microcoleaceae,Microcoleushas been difficult to be classified for a long time (Boyer et al., 2002;Strunecký et al., 2013).Numerous molecular biological evidences have indicated that the type genusMicrocoleusis a polyphyletic group with various phylogenetic positions (Strunecký et al.,2013; Geng et al., 2021a).On the phylogenetic lineage,Microcoleusis often mixed with other cyanobacterial genera (such asTychonemaAnagnostidis et Komárek 1988, etc.), and cannot form a narrow monophyletic clade (Geng et al., 2021a).According to the current definition, only a few species and the type speciesM.vaginatusare clustered into a clade (M.amplusN.L.Gardner 1927,M.acremanniiCasamatta et Johansen 2005,M.antarcticusCasamatta et Johansen 2005,M.baicalensisIzhboldina 1984,M.rushforthiiJohansen et Casamatta 2005,M.subandinusSchwabe 1944, etc.) (Siegesmund et al., 2008; Strunecký et al., 2013).Some species form completely separate clades fromM.vaginatus, so they must be removed from the genusMicrocoleusand classified into other existing cyanobacterial genera or established as new genera (Siegesmund et al., 2008; Strunecký et al.,2011).In the classical cyanobacterial taxonomic system based on morphological criteria,Microcoleuswas previously belonging to the family Oscillatoriaceae(Geitler, 1925a), and then placed into subfamily Microcoleoideae, of Phormidiaceae, Oscillatoriales due to the establishment of the family Phormidiaceae by Anagnostidis and Komárek (1988).Strunecký et al.(2013) raised the Microcoleoideae to the level of a family as Microcoleaceae, and this family was adopted in the eight-order cyanobacterial taxonomic system proposed by Komárek et al.(2014).In recent years, the revisions on the genusMicrocoleushave been much performed and more and more new genera and species have been proposed by separating from or transferring intoMicrocoleusorMicrocoleus-like group based on the polyphasic approach, such asColeofasciculuschthonoplastes(Gomont) Siegesmund, Johansen et Friedl 2008(Siegesmund et al., 2008),Wilmottiamurrayi(West et G.S.West) Strunecký, Elster et Komárek 2011(Strunecký et al., 2011),Microcoleusautumnalis(Gomont) Strunecký, Komárek et Johansen 2013(Strunecký et al., 2013),KlisinemaHeidari et Hauer 2018 (Heidari et al., 2018),Microcoleus pseudautumnalisNiiyama et Tuji 2019 (Niiyama and Tuji, 2019) andMicrocoleusanatoxicusStancheva et Conklin 2020 (Conklin et al., 2020).However, the current taxonomic revisions have not yet solved the polyphyletic problem within the genusMicrocoleus.Further taxonomic revisions onMicrocoleusorMicrocoleus-like taxa are still required, and it is expected that more new taxa will be described along the revision process.
In this study, we isolated a filamentous cyanobacterial strain withMicrocoleus-like morphology in an open channel from the project of Luanhe River Diversion into Tianjin (LRDT), the north of China.The polyphasic approach based on morphological features, molecular phylogeny, and ecology was used to characterize this strain.Possessing the unique phylogenetic position close to the recently established genusMicrocoleusiopsisGeng et Yu 2021 within the family Microcoleaceae, the presently studied cyanobacterial strain had 93.48%-93.64% similarities of 16S rRNA gene sequence withMicrocoleusiopsis, lower than the threshold of bacterial genus.Hence, the novel cyanoabcterial genusNeomicrocoleusgen.nov.and its type speciesNeomicrocoleustianjinensissp.nov.were described here.
2 MATERIAL AND METHOD
2.1 Isolation and cultivation
Fresh algae mat samples were gathered in July 2019 from Tianjin channel, Tianjin, China(39°17′22.55′′N, 117°14′42.23′′E).Live materials scraped from the mats were washed thoroughly in sterile liquid CT medium (Ichimura, 1979), and then coated onto the surface of sterile solid CT plate to obtain isolated cyanobacterial species.These solid plates were kept at 25±1 °C on a 12-h:12-h L:D cycle with a photon flux density of 30±5 μmol/(m2·s).Two or three weeks later, uncontaminated unialgal filaments or single cells were transferred into clean screw-capped tube containing 9-mL sterile CT medium.Now, the successfully isolated filamentous strain (named as CHAB 4127) in this study was stored in Wenzhou University, Wenzhou, China.The dry material of this strain (specimen No.WZUHZLTJ201901) is stored in the Herbarium of Wenzhou University (WZUH).
2.2 Morphological and ultrastructural characterization
Morphological observation of strain CHAB 4127 was performed with a Nikon Eclipse 80i microscope(Nikon, Japan) and measurements of filaments and cells were taken from more than 100 individuals with a DS-Ri1 digital camera (Nikon, Japan).Morphological images taken at 400-1 000 times were analyzed using NIS-Elements D 3.2 software.For ultrastructure examination, fresh samples of studied strain were fixed and dehydrated according to the description by Geng et al.(2021a) and the specimens were examined with an HT7700(Japan) transmission electron microscope under 80 kV on Hitachi TEM system control (Hitachi,Japan).
2.3 DNA extraction and PCR amplification
Fresh filaments of strain CHAB 4127 were collected and washed with sterile CT medium for three or four times to remove other bacterial contamination.Total genomic DNA from studied strain was extracted using the modified cetyltrimethylammonium bromide(CTAB) method (Neilan et al., 1995).The 16S rRNA gene and 16S-23S internal transcribed spacer(ITS) were PCR-amplified using the primers PA(Edwards et al., 1989) and B23S (Gkelis et al.,2005) on a BIO-RAD Thermal Cycler (Bio-Rad,Hercules, California USA), and the total PCR reaction system (20 μL) was set as follows: 10 μL of 2×PCR mix with Taq polymerase (Beijing Tsingke Biotech Co., Ltd., Beijing, China), 1 μL of genomic DNA (100 ng/μL), 0.5 μL of each primer (10 μmol/L)and 8 μL of sterile water.The PCR program was run for one cycle of 3 min at 94 °C; 34 cycles of 30 s at 94 °C, 30 s at 58 °C, and 1 min at 72 °C and then a final elongation step at 72 °C for 5 min.The obtained PCR products were purified and cloned to the pMD18-T vector (TaKaRa, Japan), subsequently transformed intoEscherichiacolicells.The positive clones including target fragment were sequenced by an ABI 3730 Automated Sequencer (PerkinElmer,Waltham, Massachusetts USA).The contigs thus obtained were assembled and edited using Sequencher 4.1.4 (Tippmann, 2004).In addition, each target fragment was sequenced for three positive clones at least.
2.4 Phylogenetic analysis
The 16S rRNA gene sequences of this novel strain CHAB 4127 generated from sequencing were initially screened and compared on the NCBI Website using the BLAST tool, and the reference sequences related to studied strain were downloaded from GenBank database to construct the molecular phylogenetic trees.Through screening, 130 sequences were corrected, aligned and produced a 1 237-bp length matrix by using MAFFT v7.475 (Katoh and Standley, 2013).The standard selection nucleic acid substitution model (GTR+I+G), as the best fitting model was selected for Bayesian inference (BI) and maximum likelihood (ML) analyses on the basis of the Akaike Information Criterion (AIC) in ModelFinder(Kalyaanamoorthy et al., 2017).For ML analysis,particular parameters were estimated by IQ-TREE v1.5.6 (Nguyen et al., 2015), and a total of 10 000 bootstrap replicates were made under standard option.As for BI analysis, particular parameters were estimated by MrBayes v3.2.6 (Ronquist et al.,2012), and two runs of eight Markov chains were run for 1 750 000 generations, sampling every 100 generations, with the initial 25% of sampled data were discarded as burn-in.For neighbor joining (NJ)analysis, the Kimura-2 model was chosen with 1 000 bootstrap replicates by using MEGA software v7.0 (Kumar et al., 2016).Three phylogenetic trees were viewed and edited in software FigTree v1.4.3(http://tree.bio.ed.ac.uk/software/figtree/), and strainGloeobacterviolaceusPCC 7421 was selected as the out-group because of its distant phylogenetic relationship and ancient evolutionary position.Thep-distance with pairwise deletion of gaps was established via MEGA software v7.0.
2.5 Construction of secondary structure of 16S-23S internal transcribed spacer (ITS)
Sequences obtained from the studied strain were tested for the presence of tRNA gene sequences using the tRNAscan-SE 2.0 web server (Lowe and Chan, 2016).The 16S-23S rRNA ITS secondary structures of D1-D1′ , Box-B and V3 helices of CHAB 4127 and other related species were determined using S-fold website (https://sfold.wadsworth.org/).The 16S-23S rRNA gene sequences obtained in this study were deposited in GenBank database with accession numbers ON854141-ON854144.The nomenclature and description of this new genus were based on the International Code of Nomenclature for algae, fungi, and plants (ICN).
3 RESULT
3.1 Morphological description
NeomicrocoleusR.Geng et R.Li gen.nov.
Diagnosis: Under light microscopy, the studied strain is morphological similar to the species ofMicrocoleus-like, but it lacks the common sheath that could contain more than one trichomes.Based on the phylogenetic analysis, this species was formed a separated clade that is close to genera of the family Microcoleaceae, likeMicrocoleusiopsis,Kamptonema,Microcoleus, andTychonema.However, the 16S rRNA sequence of this strain showed low similarity with those of Microcoleaceae genera.The significant differences in the D1-D1′, Box-B, and V3 helices also supported the establishment of this novel genus.
Description: In nature, colonies macroscopic,thallus green, always forming benthic masses.In liquid medium, filaments long, unbranched, straight or slightly curved, olive-green when young, and yellow-brown when old.Sheaths usually present,colorless.Trichomes long, unbranched, isopolar,cylindrical, not attenuated towards ends.Moreover,vegetative cells short cylindrical, isopolar, always broader than long.Apical cells usually rounded.Heterocytes and akinetes absent.Reproduction through motile hormogonia formed by necridia.Thylakoids radially arranged.
Type species:NeomicrocoleustianjinensisR.Geng et R.Li sp.nov.
Etymology: The name of this novel genus“Neomicrocoleus” was chosen because of its close relationship with the genusMicrocoleus.
NeomicrocoleustianjinensisR.Geng et R.Li sp.nov.(Fig.1)
Description: In nature, colonies are macroscopic,thallus green, always forming benthic masses.In liquid medium, thallus olive-green when young and yellow to yellow-brown when old.Filaments are unbranched, long, straight, or slightly curved,usually surrounded by sheaths.Sheaths colorless,finer, not diffluent, always open at the apex.Trichomes long, unbranched, isopolar, cylindrical,not attenuated towards ends, and constricted on cross-walls.Vegetative cells always short cylindrical,isopolar, 3.63-(5.22)-8.53 μm long, 9.63-(14.51)-17.62 μm wide, width:length ratio 2.8, with granular content, not aerotopes.Apical cells rounded,sometimes conical when old.Heterocytes and akinetes absent.Reproduction through motile hormogonia formed by necridia.Thylakoids radially arranged (Fig.2).
Reference strain: CHAB 4127.
Type locality: In Tianjin open channel, Tianjin,China (July 2019, 39°17′22.55′′N, 117°14′42.23′′E).
Holotype here designated: Dry material of strainNeomicrocoleustianjinensisCHAB 4127 was preserved in the Herbarium of Wenzhou University(WZUH), Wenzhou, Zhejiang Province, China, as specimen No.WZUH-ZLTJ201901.
Etymology: The name of species “tianjinensis”was chosen because this strain was collected from the Tianjin open channel.
Habitat: Attached on wet rock and wood surfaces.
3.2 Molecular and phylogenetic analyses
After sequencing, we obtained four 16S rRNA gene clones (1 483 bp) of studied strain CHAB 4127, sharing 99.68%-99.84% similarities with each other, and 126 representative cyanobacterial sequences from Microcoleaceae and Oscillatoriaceae,along with four CHAB 4127 clones, were selected to constructe the phylogenetic tree (Fig.3).The phylogenetic analyses indicated that all clones of CHAB 4127 clustered into a well-supported independent clade (cluster A, bootstrap values of NJ/ML/BI were 100%/100%/1.00, respectively), futher clustered with an evolutionary branch formed by

Fig.2 Ultrastructure of Neomicrocoleus tianjinensis CHAB 4127
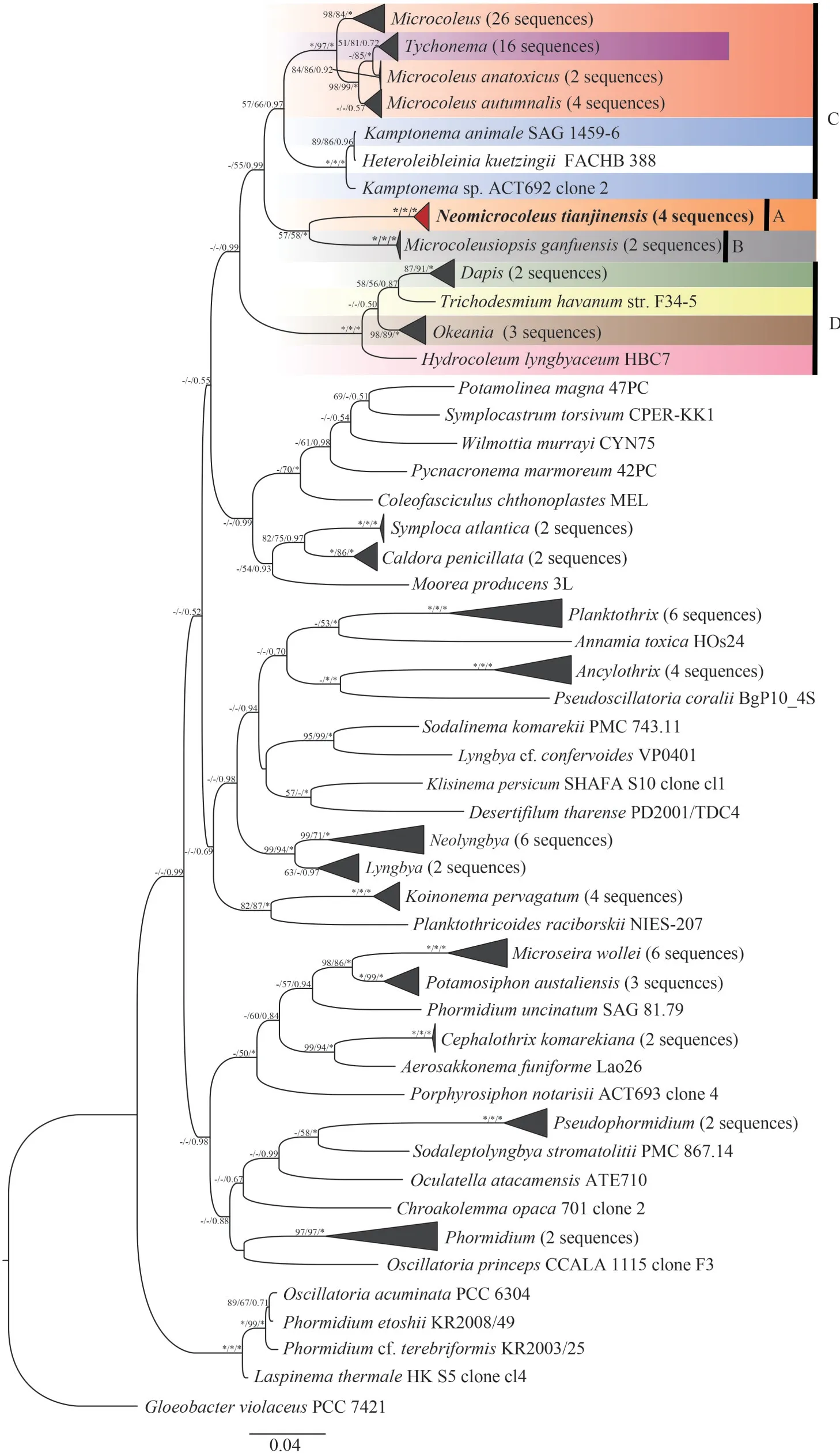
Fig.3 Bayesian inference (BI) phylogenetic tree of Neomicrocoleus tianjinensis CHAB 4127 based on the 16S rRNA gene sequences (1 237 bp)
MicrocoleusiopsisganfuensisCHAB 4138 (cluster B),with bootstrap values of 57%/58%/1.00 supported by NJ/ML/BI approaches.This unique clade was close to the members of family Microcoleaceae—Microcoleus(red part),Tychonema(purple part),KamptonemaStrunecký, Komárek et Smarda 2014(blue part),Microcoleusiopsis(gray part),DapisEngene, Tronholm et Paul 2018 (green part),TrichodesmiumEhrenberg ex Gomont 1892 (yellow part),OkeaniaEngene, Paul, Byrum, Gerwick, Thor et Ellisman 2013 (brown part), andHydrocoleumKützing ex Gomont 1892 (pink part), embedded within the family Microcoleaceae.Based on the similarity matrix of 16S rRNA gene we knew that the novel genusNeomicrocoleusshared the highest similarity of 93.97% with other existing related cyanobacterial genera, probably represented a novel cyanobacterial genus in family Microcoleaceae(Table 1).
3.3 Analyses of ITS secondary structures
The full length of the 16S-23S ITS sequences obtained from studied strainNeomicrocoleus tianjinensisCHAB 4127 was 546 bp (Table 2), and all clones ofN.tianjinensisincluded both tRNAIleand tRNAAla.According to the results of molecular and phylogenetic analyses, seven species which came from four genera of Microcoleaceae, includingMicrocoleusiopsisganfuensisGeng et Yu 2021(CHAB 4138),Microcoleusvaginatus(CSU-U-KK1),Microcoleuspseudautumnalis(Ak1609),Microcoleus anatoxicus(PTRS-2),Microcoleusautumnalis(SAG 78.79),DapispnigousaEngene, Tronholm et Paul 2018 (BCBC12-12) andOkeaniaplumataEngene, Paul, Byrum, Gerwick, Thor et Ellisman 2013 (NAC8-45), along withN.tianjinensisCHAB 4127, were used to compare ITS secondary structures(Figs.4-6).The D1-D1′ helix is generally considered to be the most conserved structure, but these eight species formed seven different shapes (Fig.4).Total length of D1-D1′ helices was ranged from 55 to 63 bases (Table 2).
MicrocoleusvaginatusCSU-U-KK1 andM.anatoxicusPTRS-2 had similar structures, whileN.tianjinensisCHAB 4127 and other 5 species formed their own unique D1-D1′ helix in base sequence and stem-loop structure.Most of them shared a 4-bp(5′-GGGA-UCCC-3′) conservative basal stem, whileN.tianjinensisCHAB 4127 andD.pnigousaBCBC12-12 had a 3-bp (5′-GGG-CCC-3′) and a 2-bp (5′-GG-CC-3′) conservative basal stem,respectively.Similarly, the terminal loops of these eight species showed a clear difference in sequence length and base arrangement, of which the novel speciesN.tianjinensisCHAB 4127 presented a 6 bases (5′-GCAAAA-3′) small loop different from the other species (15 bases inM.ganfuensisCHAB 4138 andO.plumataNAC8-45; 14 bases inM.vaginatusCSU-U-KK1,M.pseudautumnalisAk1609 andM.autumnalisSAG 78.79; 16 bases inM.anatoxicusPTRS-2; 11 bases inD.pnigousaBCBC12-12) (Fig.4).
The Box-B (Fig.5) and V3 (Fig.6) helices also showed more diverse and variable betweenN.tianjinensisstrain and other species in base sequence, sequence length and stem-loop structure,which was also strong evidence to distinguish different taxa of this study.For Box-B helix,N.tianjinensiswas similar toM.vaginatusCSU-UKK1, in whichN.tianjinensiswas comprised of one 5-bp basal stem, one 1:2 base bilateral bulge, and then followed by one 4-bp stem, one 2:1 base bilateral bulge, one 4-bp terminal stem and a 5 bases terminal loop (5′-UUUUC-3′), and was slightly different fromM.vaginatusCSU-U-KK1 in the number of bases and folding types.Furthermore,N.tianjinensishad its unique V3 helix significantly different from the other 7 species, consisting of a 4-bp basal stem, a 6:6 base bilateral bulge followed by a 9-bp terminal stem, and ended with a 3 bases terminal loop (5′-ACU-3′).M.ganfuensisCHAB 4138, the closest genus toN.tianjinensisin phylogenetic relationship (Fig.3), had a completely different secondary structure fromN.tianjinensis, clearly distinguishing and further supportingNeomicrocoleusas a new genus.
4 DISCUSSION
The scarcity of water resources and water quality is one of the major problems faced by the world(Liu and Yang, 2012).Therefore, the construction of the Water Diversion Projects that can regulate the uneven distribution of water resources in time and space, and realize the rational and optimal allocation of water resources has become one of the important solutions in China.Such huge projects often involve long-distance water transportation, and many new open water channels have been produced in the process, forming these special biotopes.The project of Luanhe River Diversion into Tianjin (LRDT) we investigated is one of the notable drinking water supply projects (Cai et al., 2017).In previous studies, the benthic cyanobacterial mats grown on both sides of the open channels were mainly composed of taxa in Chroococcales and Oscillatoriales, such asChroococcus-like (Geng et al., 2021b),Oscillatorialike (Cai et al., 2017),Lyngbya-like (Geng et al.,2021c) andMicrocoleus-like cyanobacteria (Geng et al., 2021a).To better understand the role of theseOscillatorean cyanobacteria in the Water Diversion Process, it is much necessary to accurately describe and correctly characterize them based on the modern cyanobacterial taxonomy.
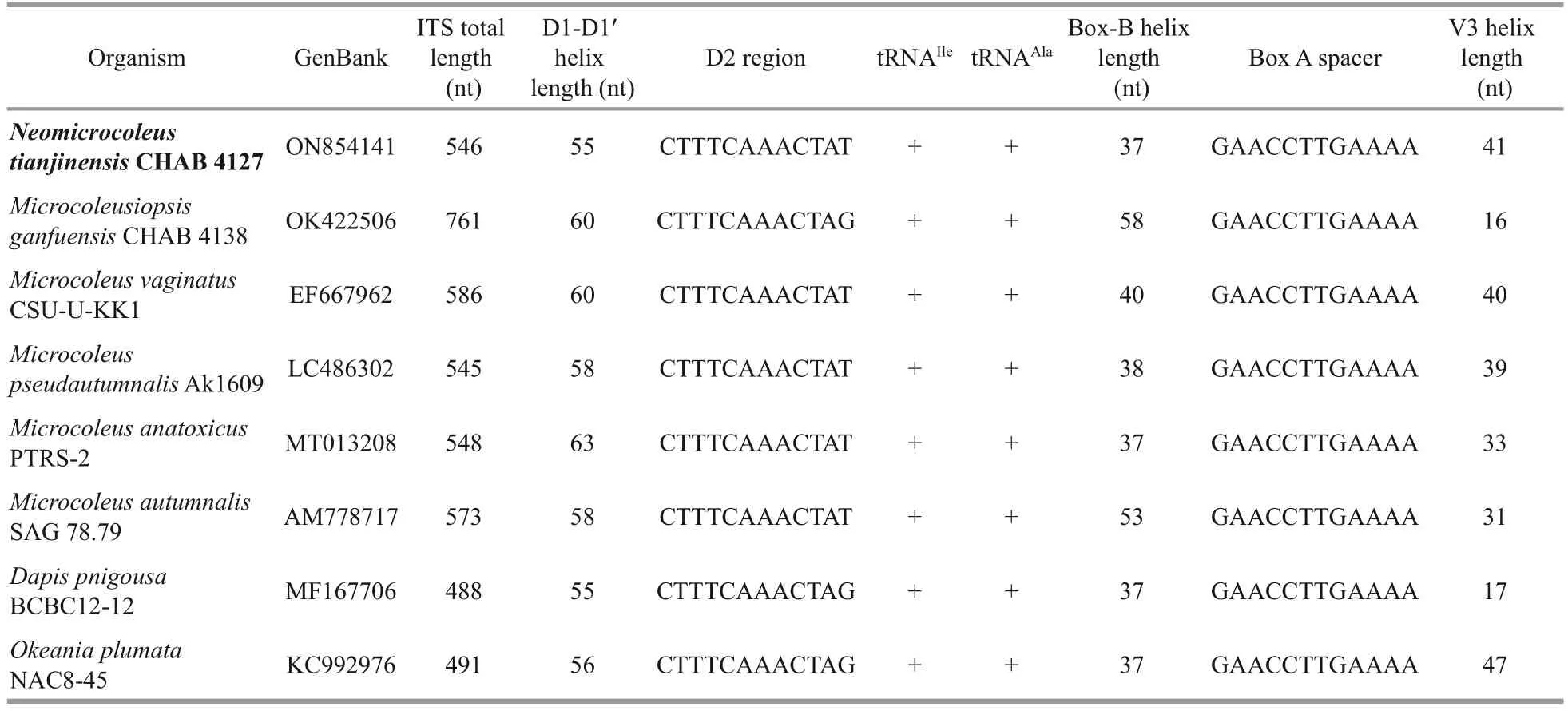
Table 2 Analyses on ITS of 16S-23S region for Neomicrocoleus tianjinensis and its related species
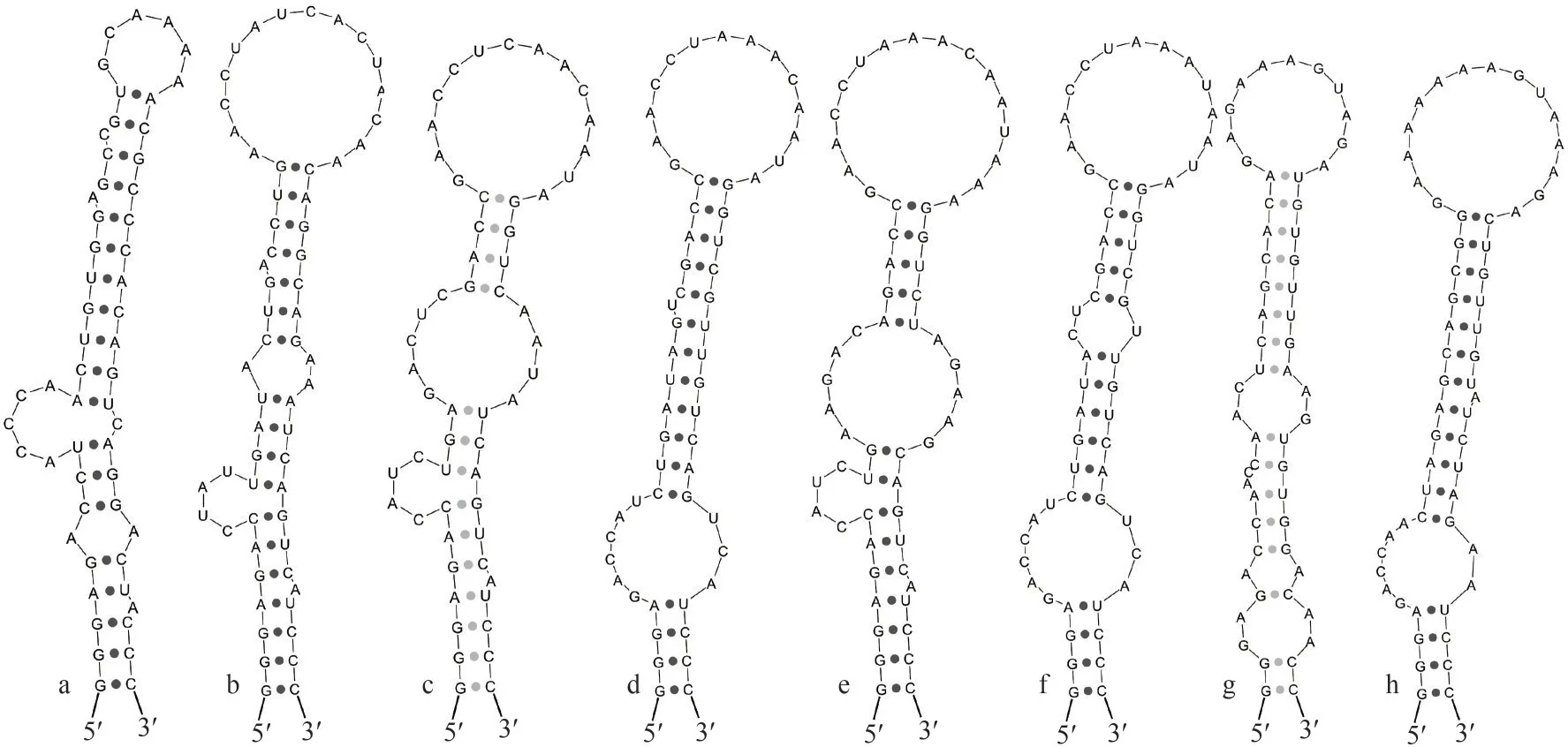
Fig.4 D1-D1′ helix in Neomicrocoleus tianjinensis and other closed species
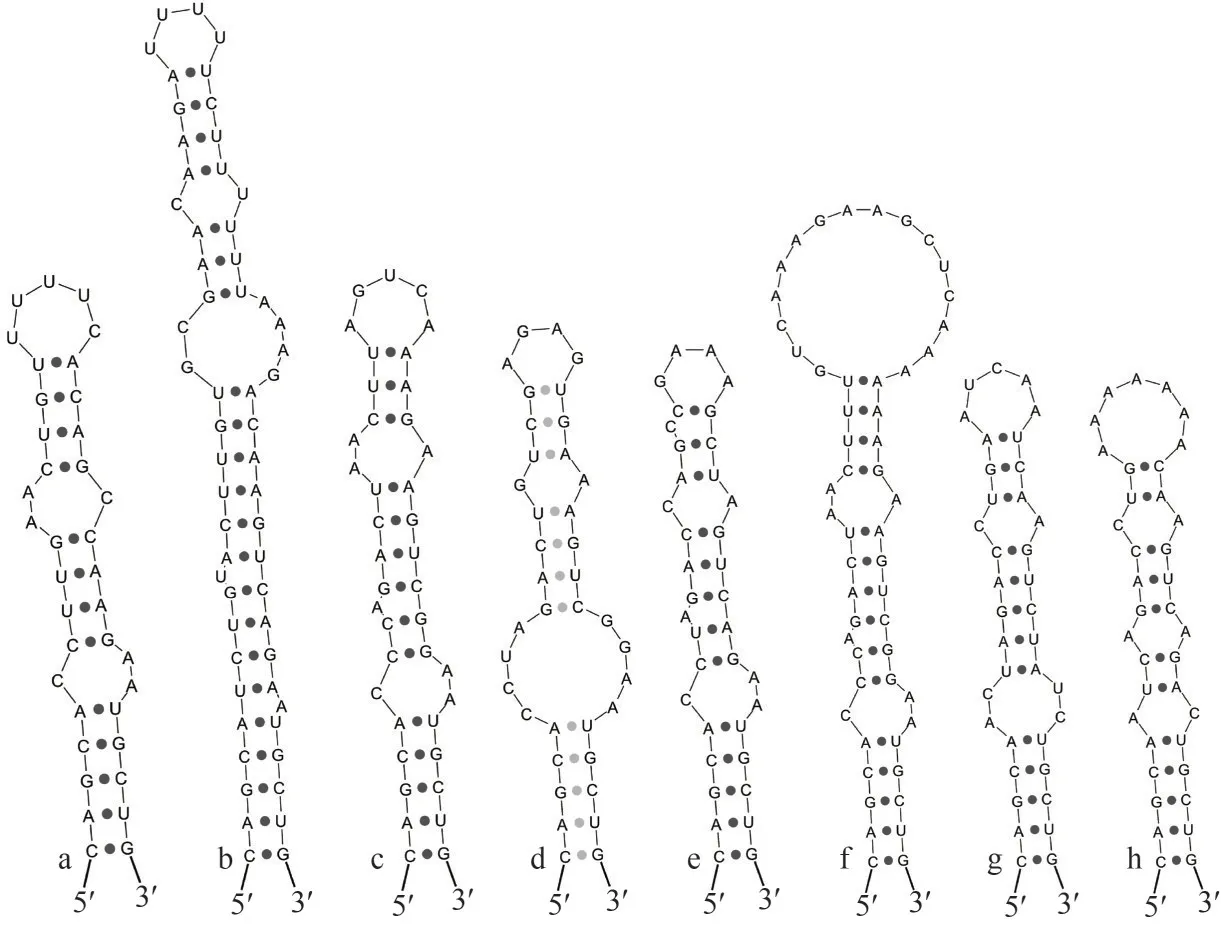
Fig.5 Box-B helix in Neomicrocoleus tianjinensis and other closed species

Fig.6 V3 helix in Neomicrocoleus tianjinensis and other closed species
Modern cyanobacterial taxonomic system aims to accurately identify and revise the complex polyphyletic taxa (families, genera, and species) in cyanobacteria, and promote the classification system to monophyletic, clear, and narrowly defined levels(Anagnostidis and Komárek, 1985; Casamatta et al.,2005; Siegesmund et al., 2008; Komárek et al.,2014).The current taxonomy system does not solve the polyphyletic origination in cyanobacteria at the family and genus level.Some oscillatorialean groups, such asLyngbyaAgardh ex Gomont 1892,MicrocoleusandOscillatoriaVaucher ex Gomont 1892 are still confused at the genus level (Engene et al., 2010; McGregor and Sendall, 2015; Martins et al., 2019; De Lima and Branco, 2020).In the present study, the examined cyanobacterial strain from the open channel was in such an unclear situation.Morphological features of this strain fell into the range ofMicrocoleus-like cyanobacteria characteristics.It was similar in morphology to the genusMicrocoleusiopsis, butNeomicrocoleus tianjinensisCHAB 4127 could be differentiated fromMicrocoleusiopsisganfuensisCHAB 4138 based on morphology.Trichomes ofM.ganfuensisCHAB 4138 were narrower, at 4.52-(5.69)-6.18 µm broad, with a width:length ratio 1.8, but trichomes ofN.tianjinensisCHAB 4127 were 9.63-(14.51)-17.62 μm wide, having a larger cell length-width ratio, with trichomes being more olive-green,rather than blue-green ofM.ganfuensis(Fig.1).In addition,N.tianjinensisCHAB 4127 was also similar to the species ofMicrocoleus,Tychonema,andKamptonema, but its morphological characteristics,such as lacking bundle formation of trichomes covered by a common sheath, without isodiametric cells, and usually surrounded with sheaths, confirmed the distinction ofN.tianjinensisfrom the three genera, respectively (Anagnostidis and Komárek,1988; Strunecký et al., 2013, 2014).
The molecular and phylogenetic results indicated that this strain was clustered with the newly established genusMicrocoleusiopsis(Fig.3 cluster A), separating from other existing groups of Microcoleaceae.In details, its unique position at the phylogenetic tree was located in two evolutionary branches of family Microcoleaceae, one as the clade formed byMicrocoleus,Tychonema, andKamptonemaclade (Fig.3 cluster C), the other as the one byDapis,Okeania, andHydrocoleum(Fig.3 cluster D).The genusMicrocoleusiopsis, established in 2021,was one novel filamentous cyanobacterium without heterocyst differentiation isolated from Ganfu channel, Jiangxi Province, China (Geng et al.,2021a).Comparison of the DNA sequences showed thatN.tianjinensishad 93.48%-93.64% similarities of 16S rRNA gene sequence withMicrocoleusiopsis ganfuensis, less than 95% which was considered as the threshold in bacteriology to differentiate bacterial genera (Wayne et al., 1987; Stackebrandt and Goebel, 1994; Stackebrandt and Ebers, 2006).Therefore, based on the separated phylogenetic positions and the lowed 16S rRNA gene similarity with other related genera (Table 1), the examined strain was strongly verified as an independent new genus of Microcoleaceae.
It was worth mentioning that the 16S-23S rRNA ITS secondary structures of D1-D1′, Box-B, and V3 helices inN.tianjinensisCHAB 4127 also showed obviously differences from other adjacent genera of Microcoleaceae (Figs.4-6).As an important tool for cyanobacterial taxonomy, the ITS secondary structures are usually used to distinguish different genera and species of cyanobacteria (Iteman et al.,2000; Boyer et al., 2002; Johansen and Casamatta,2005; Johansen et al., 2011).The clear differences betweenN.tianjinensisandM.ganfuensisin the secondary structures were also strong evidence to distinguish these two taxa.
Additionally, the habitat feature plays a nonnegligible role in the cyanobacterial classification(Řeháková et al., 2007).The species of Microcoleaceae have a wide range of ecological niches in various ecosystems, including planktic (freshwater/marine),benthic, subaerial, halophytic, and even extreme environments (Anagnostidis and Komárek, 1988;Boyer et al., 2002; Manen and Falquet, 2002;Strunecký et al., 2013, 2014; Engene et al.,2018; Heidari et al., 2018; Geng et al., 2021a).N.tianjinensisCHAB 4127 was a benthic freshwater filamentous cyanobacterium isolated from Tianjin channel, Tianjin, China.Similarly,Microcoleusiopsiswas also separated from the water channel with flowing water habitat likeNeomicrocoleus, located in the subtropical zone (whileNeomicrocoleuswas located in the temperate zone).TheMicrocoleusspp.are mainly benthic and soil species, whileTychonemaspp.usually are planktic or benthic species, members of the two genera are currently mixed in phylogenetic relationships (Strunecký et al., 2013; Geng et al., 2021a).In addition,Kamptonema, close toNeomicrocoleusin phylogenetic tree, is generally of freshwater origin (Strunecký et al., 2014).
Since the establishment of the family Microcoleaceae in 2013, more and more cyanobacterial genera have been reclassified or migrated into this family based on the polyphasic approach (Strunecký et al., 2013).In the system of cyanobacterial taxonomy proposed by Komárek et al.(2014), Microcoleaceae included those genera originally belonging to the subfamily Microcoleoideae, such asMicrocoleus,Hydrocoleum,Symplocastrum(Gomont) Kirchner 1898,SirocoleumKützing ex Gomont 1892, andLyngbyopsisGardner 1927, and several genera transferred from other families such asPlanktothrixAnagnostidis et Komárek 1988 (Suda et al., 2002; Lin et al., 2010),ArthrospiraSitzenberger ex Gomont 1892 (Manen and Falquet, 2002; Dadheech et al., 2010), andTrichodesmium(Orcutt et al., 2002), and the newly established generaOxynema(Chatchawan et al.,2012),AnnamiaNguyen 2013 (Nguyen et al., 2013),JohanseninemaHašler, Dvorák et Poulícková 2014 (Hašler et al., 2014a, b), andKamptonema(Strunecký et al., 2014), reaching 19 genera in total(Komárek et al., 2014).After 2014, five novel genera,CaldoraEngene, Tronholm et Paul 2015(Engene et al., 2015),Dapis(Engene et al., 2018),Klisinema(Heidari et al., 2018),SodalinemaCellamare, Duval, Touibi, Djediat et Bernard 2018(Cellamare et al., 2018), andMicrocoleusiopsis(Geng et al., 2021a) were added into Microcoleaceae.This study describedNeomicrocoleusas the 25thmember in Microcoleaceae, also providing a case for the further taxonomic revision onMicrocoleuslike groups.However, up to now, the problem of polyphyly of Microcoleaceae remains unresolved,and the members of Microcoleaceae and other related families such as Oscillatoriaceae are phylogenetically mixed.In the future, more filamentous cyanobacterial strains and molecular data will be expected to be obtained for further taxonomic revision on the family Microcoleaceae, gradually with the ideal goal as establishing a monophyletic classification system at both genus and family levels.
5 CONCLUSION
In conclusion, we successfully isolated a filamentousMicrocoleus-like strain named as CHAB 4127 from the open channel in Tianjin,China.The strain was taxonomically examined using the polyphasic method.With the combined results on morphology, ultrastructure, ecology,16S rRNA gene sequences, phylogeny and ITS second structures, a novel cyanobacterial genus,Neomicrocoleuswas established, with the type speciesNeomicrocoleustianjinensis.Currently, the artificial open channels are gradually explored and investigated as new habitats for harboring high diversity of cyanobacterial resources, and it is expected that more new taxa will be discovered and described in the future to further increase the species diversity of cyanobacteria.
6 DATA AVAILABILITY STATEMENT
The data that support the findings of the study are available from the corresponding author upon reasonable request.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Contrasts of bimodal tropical instability waves (TIWs)-induced wind stress perturbations in the Pacific Ocean among observations, ocean models, and coupled climate models*
- Variability of the Pacific subtropical cells under global warming in CMIP6 models*
- Identification of thermal front dynamics in the northern Malacca Strait using ROMS 3D-model*
- Magmatic-tectonic response of the South China Craton to the Paleo-Pacific subduction during the Triassic: a new viewpoint based on Well NK-1*
- An improved positioning model of deep-seafloor datum point at large incidence angle*
- Microplastics in sediment of the Three Gorges Reservoir:abundance and characteristics under different environmental conditions*