New record of the feather star Palaeocomatella hiwia(Echinodermata: Crinoidea: Comatulida) from Zhenbei Seamount in South China Sea: new insights into the taxonomic position of Palaeocomatella*
2024-02-27ShaoSUNZijieMEIZhongliSHAJingWANGNanshengCHEN
Shao’e SUN, Zijie MEI, Zhongli SHA,**, Jing WANG, Nansheng CHEN
1 Department of Marine Organism Taxonomy and Phylogeny, Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China
2 CAS Key Laboratory of Marine Ecology and Environmental Sciences, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
3 Laoshan Laboratory, Qingdao 266237, China
4 Shandong Province Key Laboratory of Experimental Marine Biology, Institute of Oceanology, Chinese Academy of Sciences,Qingdao 266071, China
5 Center for Ocean Mega-Science, Chinese Academy of Sciences, Qingdao 266071, China
6 University of Chinese Academy of Sciences, Beijing 100049, China
Abstract One specimen belonging to the family Comatellinae was collected from the Zhenbei Seamount (332.5-478.2 m) in the South China Sea in July 2022.Based on the morphological characters,the specimen was identified as Palaeocomatella hiwia McKnight, 1977.It is first recorded from China Sea and redescribed in detail.This specimen differs from the original description from New Zealand for never showing syzygy at br4+5 or br5+6 on interior and br1+2 on exterior arms.However, it is much conform to the redescription to specimens from Indonesia, with only differences in position of the second syzygy and distalmost pinnule comb.Specimen is deposited in the Institute of Oceanology, Chinese Academy of Sciences.Phylogenetic analyses based on the mitochondrial c oxidase subunit I (COI) and 16S rRNA genes indicated that P.hiwia was nested within the tribe Phanogeniini and clustered with Aphanocomaster pulcher.Furthermore, P.hiwia showed same morphological features in terms of mouth placement, comb location, and number of comb teeth rows as other genera of Phanogeniini.Therefore, we suggest that the genus Palaeocomatella should be put in the tribe Phanogeniini.
Keyword: new record; Palaeocomatella hiwia McKnight, 1977; South China Sea; phylogenetic relationship; taxonomy
1 INTRODUCTION
The Comatulidae Fleming, 1828, is the second most diverse family of featherstars (followed by Antedonidae Norman, 1865) of extant comatulids,which is widely distributed in Indo-western Pacific region adjacent temperate seas.They are most common in shallow water, forming an ecologically component of benthic communities (Hoggett and Rowe, 1986; Messing, 2001; Kogo and Fujita, 2014;Summers et al., 2017).Because the members of Comatulidae from deep-sea are known from only a few species, their species diversity, morphological variability, taxonomic relationship and distribution still poorly understood.
The most easily recognized apomorphic features of the family Comatulidae is the comb-likes structure on the terminal several to many ossicles of oral pinnules (Messing, 2001; Summers et al.,2017).Clark (1931) mentioned 90 extant species in 19 genera and three subfamilies, and this taxonomic system was accepted by most researchers until 1986.Hoggett and Rowe (1986), Rowe et al.(1986), and Messing (2001) placed 95 species in 15 genera of four subfamilies and 6incertaesedisgenera.Summer et al.(2014) revised the 95 species and retained 4 genera in the subfamily Comatellinae,13 genera in the subfamily Comatulinae, and 4 assignment uncertain genera.Currently, two subfamilies, six tribes, 22 genera, 97 nominal species and 6 assignment uncertain species are known in the family Comatulidae (Summers et al., 2017; Messing et al., 2023).Previous molecular phylogenetic relationships (Hemery et al., 2013; Rouse et al.,2013) strongly supported the monophyly of Comatulidae, however, White et al.(2001) and Rouse et al.(2013) recovered some subfamilies and genera as paraphyletic.Summer et al.(2014)conducted a molecular phylogenetic analysis of Comatulidae, containing 43 nominal species spanning 17 genera and all subfamilies, and revealed two genera as paraphyletic.
The genusPalaeocomatellaAH Clark, 1912 is an extant genus, containing three species, which only distributes in the Western Pacific, with depth range 212-808 m (Messing et al., 2000; Messing,2001, 2007; Ocean Biodiversity Information System,https://obis.org/).P.difficilis(Carpenter, 1888) is recorded from southern Indonesia (Kepulauan Kai).P.hiwiaMcKnight, 1977 is distributed from New Zealand, the Kermadec Islands, and Indonesia(Messing et al., 2000), andP.decora(Clark, 1912)is described from the Loyalty Ridge off New Caledonia and East China Sea (Messing, 2001).Although previous molecular studies analyzed the phylogenetic relationships of the genera of Comatulidae, noPalaeocomatellaspecies was represented (Hemery et al., 2013; Rouse et al., 2013;Summer et al., 2014) The phylogenetic position ofPalaeocomatellais unclear and it is still considered as an incertae sedis genus.
In July 2022, surveys conducted by the Institute of Oceanology, Chinese Academy of Sciences, at the Zhenbei Seamount in the South China Sea.Collected benthic organisms included several crinoids.After careful examination, one specimen was identified asP.hiwia, which had only been reported from Southwest Pacific previously.Herein,we formed the first report ofP.hiwiafrom the Zhenbei Seamount in the South China Sea.Thus,the distribution range of this species extends from Southwest Pacific to Northwest Pacific.In addition,we present the molecular phylogenetic analyses,evaluating the systematic status ofPalaeocomatellaand several major deep-sea groups of Comatulidae.Furthermore, we discussed the evolutionary transition of the position of mouth and the comb-like structure.Our study will provide new insights into the evolution of Comatulidae and other crinoids.
2 MATERIAL AND METHOD
2.1 Sampling and preservation
A single specimen ofPalaeocomatellasp.was collected at 332.5-478.2 m from the Zhenbei Seamount in the South China Sea (station FX-Dive 310, 15°03′N, 116°33′E) using the Remotely Operated Vehicle submersible (ROV), carrying by the R/VKexue(Science in Chinese) in 2022.The specimen was preserved in 75% ethanol, assigned the voucher number WJ56-NH119, and deposited at the Marine Biological Museum of Chinese Academy of Sciences (MBMCAS), Institute of Oceanology,Chinese Academy of Sciences (IOCAS).
2.2 Morphology observation
According to the original descriptions,illustrations (McKnight, 1977; Hoggett and Rowe,1986; Messing et al., 2000), and taxonomic keys(Messing, 2001, 2007; Summers et al., 2017), the specimen was identified asP.hiwia.The preserved specimen was examined and dissected with a ZEISS SteREO Discovery V20 dissecting microscope.The photographs were taken by Nikon D800 and Canon EOS 6D camera.For finer structures that were difficult to measure directly, ZEISS Axiocam 506 microscope camera was used to take photographs,and Leica LAS Image Analysis software was used to conduct the measurements.
2.3 DNA extraction, sequencing, and phylogenetic analysis
Total genomic DNA of a specimen ofP.hiwiawas extracted from the distal arms and pinnules using E.Z.N.A.®Tissue DNA Kit (OMEGA)according to the manufacturer’s instructions.The DNA was eluted in sterile distilled H2O (RNase free) and stored at -20 ℃.Two mitochondrial gene fragments (COI, ~600 bp; 16S, ~500 bp) were extracted from its mitochondrial genome (determined through Illumina sequencing in our unpublished work) with GenBank accession Nos.OP856953 and OP857544.
Before phylogenetic analysis, we estimated the intergeneric genetic distance based on the available COI barcoding sequences of 18 genera in the family Comatellinae (Supplementary Table S1).Kimura-2 parameter (K2P) genetic distances were calculated using MEGA v.6 (Tamura et al., 2013).For the phylogenetic analysis, the concatenated sequences of COI and 16S fragments of 40 species (18 genera)from Comatellinae and two outgroups from Bathycrinidae were used (Supplementary Table S1).The nucleotide sequences for COI and 16S genes were all aligned with MEGA v.6 (Tamura et al.,2013).However, for COI gene, the alignment was performed based on codon positions.
Phylogenetic trees were constructed by maximum likelihood (ML) and Bayesian inference (BI)analysis.PartitionFinder 2.1.1 (Lanfear et al., 2017)was used to select the most suitable partitioning scheme and substitution model (Supplementary Table S2).ML analysis was conducted by IQ-TREE web server (Trifinopoulos et al., 2016) using the best-fit partitioning scheme and model with 5 000 ultra-fast bootstrap replications (Minh et al., 2013).BI analyses were performed using MrBayes 3.1 software (Ronquist and Huelsenbeck, 2003) with partitioning model.The Markov chain Monte Carlo(MCMC) was run for 5 000 000 generations, with a sampling frequency of 100 generations to allow sufficient time for convergence (the standard deviation of split frequencies less than 0.01).The effective sample size (ESS) values for all sampling parameters were checked with Tracer v1.7(Rambaut et al., 2018).The first 12 500 trees were discarded as burn-in and the remaining trees were used to compute the 50% majority-rule consensus tree and the posterior probabilities (PP).
3 RESULT AND DISCUSSION
3.1 Systematics
Family Comatulidae Fleming, 1828
Subfamily Comatulinae Fleming, 1828
GenusPalaeocomatellaAH Clark, 1912
Actinomelra(part) Carpenter, 1888.Challenger Reports, Zoology, 26∶93.
PalaeocomalellaClark, 1912.Proc.Biol.Soc.Washington, 25∶18; 1915.American Naturalist, 49∶525, 539; 1918.Unstalked Crinoids of the Siboga Exped., 3, 7;Gislen, 1924.Zool.Bidrag fran Uppsala,9∶38.Clark, 1931.Bull.U.S.Nat.Mus.,82:153.Messing, 2001.Bull.Biol.Soc.Washington, 291.Summers et al., 2017.Zootaxa, 184.
Type species:ActinometradifficilisCarpenter, 1888.
Diagnosis: Mouth central; more than 10 arms (up to 28); centrodorsal circular and large with the cirri present; all of the division series are 2; IBr2series united by synarthry; 1stsyzygy at 3+4 on arms arising from IBr; 1stsyzygy at 1+2 or 3+4, or both on the outer arms arising from IIBr and following brachitaxes; inner arms with 1+2 alone or 1+2, 3+4(occasionally 1+2, 4+5); distal intersyzygial interval 3-5; pinnule combs occur on P3-P6; comb teeth triangular, curved, confluent with exterior side of pinnule; no transverse proximal tooth; terminal comb tooth fan-shaped (Clark, 1931; Messing,2001; Summers et al., 2017).
Palaeocomatella hiwiaMcKnight, 1977 (Figs.1-2)
ComissiahiwiaMcKnight, 1977.NZOI Records,94, Figs.1-2.
PalaeocomatellahiwiaHoggett and Rowe, 1986.Zool.J.Linn.Soc.124; Messing et al., 2000.Mémoires de le Museum National d’Histoire Naturelle, Paris 184:642-644; Messing, 2001.Bull.Biol.Soc.Washington, 292; Messing, 2007.Pacific Science, 61:99.Summers et al., 2017.Zootaxa, 184.

Fig.1 Palaeocomatella hiwia McKnight, 1977, collected from the Zhenbei Seamount in the South China Sea(general view)

Fig.2 Palaeocomatella hiwia McKnight, 1977
Material examined
MBM287586, Zhenbei Seamount in the South China Sea, station FX-Dive 310 (15°03′N, 116°33′E),depth 332.5-478.2 m, Aug.2022.
Description
Centrodorsal disc-shaped, diameter 4.91 mm;aboral pole largely rounded, relatively wide, and flat in plane (Fig.2a).
Cirri crowded and irregularly arranged in two rows, with one or two slender immature cirri(Fig.2b-c); XXⅥ, 17-18,11-13 mm; c1very short,with length-to-width (L/W) 0.4-0.5; c2with L/W 1.3-1.5; c3longest, cylinder-shaped, L/W 2.2-2.4;after c3, following cirrals gradually shorter and wider; distal cirrals slightly wider than long, L/W 0.7-0.8.c4L/W 1.4-1.6; c5approximately equal in length and width.Aboral distal tubercles appear on c4or c5and located at the edge of cirrals.The following tubercles appear in middle part of cirrals,compressed and tall, rounded or triangular.Opposing spines triangular in shape.Terminal claws sharply curved, longer than the penultimate cirral.
Radials partially visible and almost hidden by centrodorsal, small interradial triangular shape.In all branchitaxes, every first brachial and every second brachial united by synarthry.IBr1mostly visible, relatively short, rectangular, W/L 3.5-4.1, in contact with proximal corners so that arm bases are well separated.IBr2axil pentagonal, W/L 2.5-2.7,with short lateral margins.ⅡBr1short, rectangular,W/L 2.1-2.3; ⅡBr2axil pentagonal, W/L 2.2-2.5,asymmetrical (Lateral margin supporting ⅢBr longer than the other margin).ⅢBr1oblong, W/L 2.2; ⅢBr2similar to preceding axil but symmetrical,W/L 2.0.Ⅱ-ⅢBr series more rounded aborally(Fig.2d).
Arms 22, approximately 60 mm long.On interior arms, br1+2square, wider than long, W/L 1.1-1.3.br3distinctly longer, W/L 1.5-1.7.brr3-5rectangular,W/L 1.5-1.9.Following brr becomes triangular with distal margins, W/L 1.0; distal brr cuneate, W/L 1.0.On exterior arms, br1rectangular, W/L 1.8-2.0; br2approximately same as br1, W/L 2.0.br3+4from ⅡBr series 1.5 mm in diameter, W/L 1.4.brr6-8oblong,W/L 1.6-1.7.Syzygy at br3+4on exterior arms and at br1+2on interior arms arising from Ⅱ-ⅢBr series;second syzygy br11+12or br13+14; distal interval 3-4,meanly 4 (Fig.3).

Fig.3 Aboral ray base and distal portion of arms showing distal intersyzygial interval of 3-4
Oral pinnules slender, confluent exterior teeth, no transverse proximal tooth.Distal comb teeth triangular and sometimes fan-shaped (Fig.4a).P1 36-37 segments, 9 comb teeth, 12-13 mm; proximal pinnulars wider than long, rhombic; middle pinnulars slightly longer than wide; comb teeth tall and triangular.P2 shorter, 33 segments, 8 comb teeth, 10 mm; middle pinnulars with L/W up to 1.4.P3 slightly shorter than P2, 27 or 29 segments, 5-6 comb teeth, 8 mm.P4 24-25 segments, 5 comb teeth, 6 mm.P5 without comb teeth, 18 segments,6 mm.P6 first genital pinnule, 17 segments, 6.5 mm.Middle pinnules 18 segments, 7.4 mm.Distal pinnules slimmer, up to 16 segments, 8.5 mm.
Mouth central, large and anal canal rectangular(Fig.4b).Color pale pink in fresh samples, and light yellow and white after alcohol immersion.
Distribution
Palaeocomatellahiwiais a species previously known from the Southwestern Pacific Ocean,including Indonesia, the Kermadec Islands, and New Zealand, at depths from 205 to 924 m(Messing et al., 2000).The present study is the first report of the species from the Zhenbei Seamount in the South China Sea, which extends its distribution to the northwestern Pacific Ocean.
Remarks
The fan-shaped terminal comb tooth is the special characteristic of the genusPalaeocomatella,nothing similar being known to occur in any other confamilials (Messing, 2001).P.decorais distinguished fromP.difficilisandP.hiwiaby its transversely oriented first comb tooth, and the basis of distribution of proximal arm syzygies distinguishedP.difficilisandP.hiwia(Messing, 2007;Summers et al., 2017).

Fig.4 Triangular and fan-shaped comb teeth confluent with edge of pinnule (a); central mouth and marginal anal papilla (b)
This is the first time forP.hiwiarecorded in South China Sea, as the species was previously known only from Indonesia, the Kermadec Islands,and New Zealand.The specimen differs from the original description by McKnight (1977) (New Zealand) for never showing syzygy at br4+5or br5+6on interior and br1+2on exterior arms.McKnight(1977) identifiedP.hiwiaas showing br1+2,4+5or br1+2,5+6on interior arms and br3+4or br1+2,3+4on exterior arms.It differs from redescription description form Kai Islands in Indonesia by Messing et al.(2000) by the presence of the second syzygy br11+12or br13+14, with distal interval 3-4.The specimens described from Indonesia showed the second syzygy br13+14or br14+15, showing distal intersyzygial interval of 4-5.Additionally, the combs of the examined specimen extended to P4,while the distalmost pinnule comb of Indonesia specimens was on P3.The examined specimen ofP.hiwiawas collected in 332.5-478.2 m, overlapping but not completely within the bathymetric range of the Indonesia specimen (212-459 m), but shallower than the records from Kermadec Islands (542-545 m)and New Zealand (602-808 m).
3.2 Barcoding, phylogenetic relationships, and taxonomic implication
The intergeneric genetic distance (K2P) of family Comatellinae was established based on COI gene.The intergeneric distances within Comatellinae ranged from 5.8% to 17.8%.Palaeocomatella(P.hiwia) showed the nearest distance withComissia(5.8%), and the greatest distance withComatella(15.2%).It is not surprising thatC.hiwiahas been regarded as a synonym ofP.hiwia(Hoggett and Rowe, 1986).
The phylogenetic tree derived from BI and ML analyses showed congruent results (Fig.5), which returned two previously accepted subfamilies,Comatellinae and Comatulinae, as well supported monophyletic clades.The validity of the five tribes of subfamily Comatulinae, namely, Comasterini,Capillasterini, Neocomatellini, Comatulini and Phanogeniini, and was supported by our analyses(see Fig.5).P.hiwia(previouslyincertaesedis) was nested within Phanogeniini.Although the bootstrap value for the node ofP.hiwiaandAphanocomaster pulcheris not high, the node of the generaPalaeocomatella,Phanogenia, andAphanocomasteris well supported.Therefore,P.hiwiashould be put in Phanogeniini, suggesting a new taxonomic position ofP.hiwia.This relationship did not contradict the classification based on morphological evidences.Among Comatellinae, the genera,Aphanocomaster,Comissia,Palaeocomatella, andPhanogeniahave common features in terms of mouth placement, comb location, and number of comb teeth rows (Summer et al., 2014).In this genus, the mouth lies in the center of the oral surface of the visceral mass with the anal papilla offcenter.Their tooth arranged only in one row, which confluent with exterior edge of the pinnular.Aiming for a classification system that is based on both molecular phylogeny and morphological character,we support that Palaeocomatella should be put in the tribe Phanogeniini (see Fig.6 for detail).
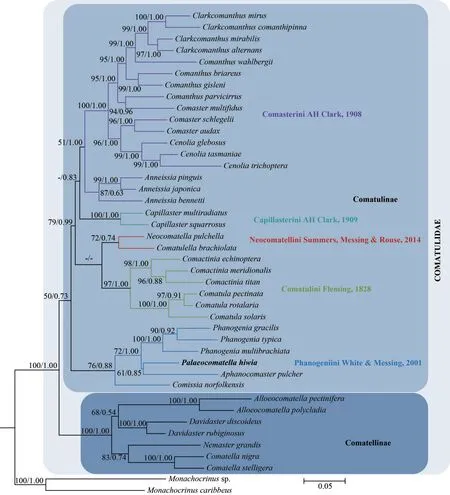
Fig.5 Maximum likelihood (ML) and Bayesian Inference (BI) trees of family Comatulidae using combined sequences of COI and 16S
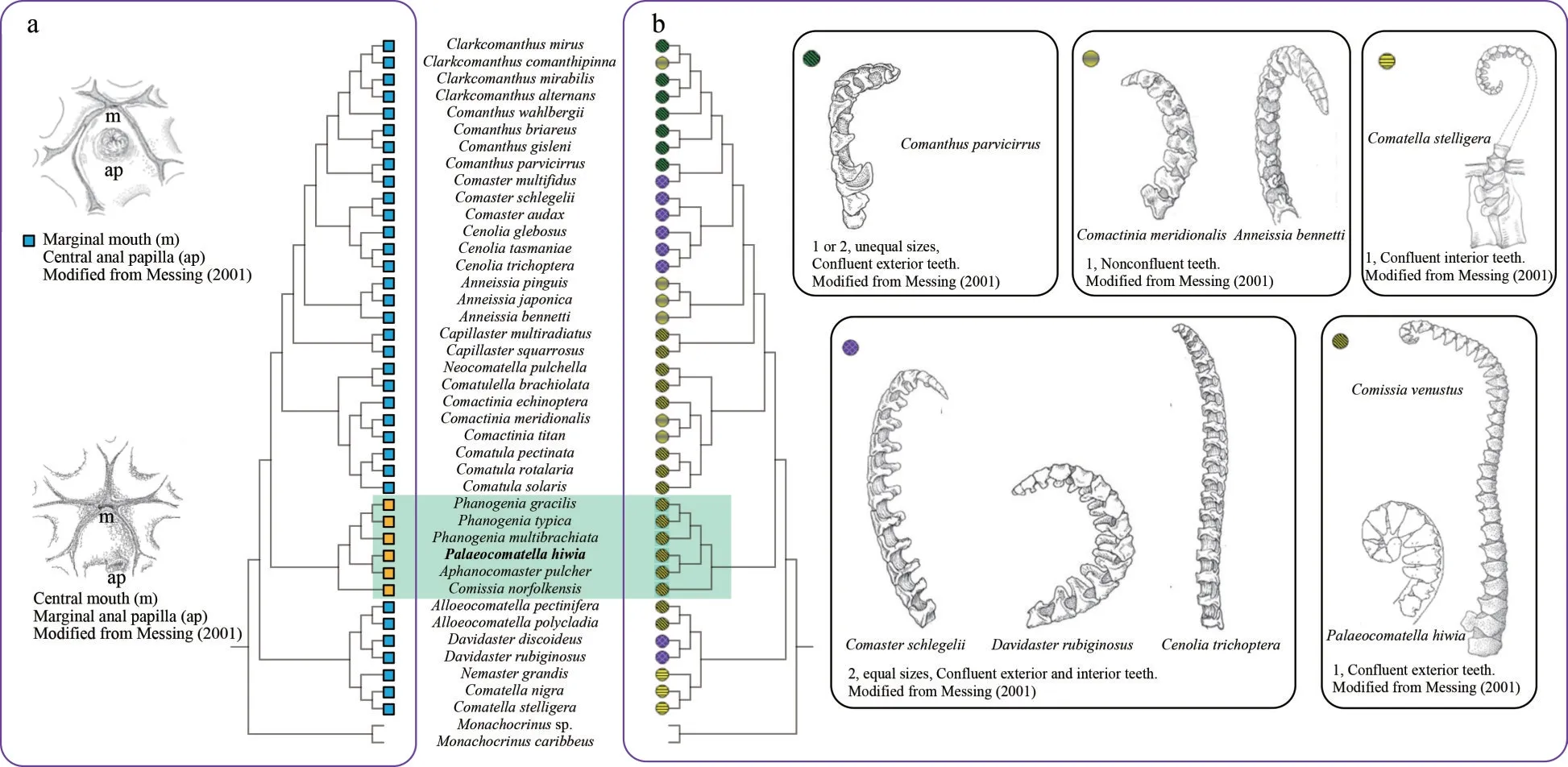
Fig.6 Phylogenetic topologies with mapped morphological characteristics
4 CONCLUSION
The morphological characters of the specimen was similar to those described by McKnight (1977)and Messing et al.(2000).PalaeocomatellahiwiaMcKnight is first recorded form China Sea.However, the position of the syzygy on arms and pinnule with distalmost comb are slightly different.The result of DNA barcoding analysis showed thatPalaeocomatellahave the nearest distance withComissia(5.8%).Phylogenetic analyses indicated thatP.hiwiawas nested within the tribe Phanogeniini and clustered withAphanocomaster pulcher.Combining the morphology and phylogenetic analyses, the genusPalaeocomatellashould be put in the tribe Phanogeniini.
5 DATA AVAILABILITY STATEMENT
The authors declare that the data supporting the findings of this study are available within the article and the appendix sections.The molecular sequences will be available from the NCBI.
6 ACKNOWLEDGMENT
We thank the assistance of the crew of R/VKexueand ROVFaxian(Discover in Chinese) for sample collection.
杂志排行

Journal of Oceanology and Limnology的其它文章
- Contrasts of bimodal tropical instability waves (TIWs)-induced wind stress perturbations in the Pacific Ocean among observations, ocean models, and coupled climate models*
- Variability of the Pacific subtropical cells under global warming in CMIP6 models*
- Identification of thermal front dynamics in the northern Malacca Strait using ROMS 3D-model*
- Magmatic-tectonic response of the South China Craton to the Paleo-Pacific subduction during the Triassic: a new viewpoint based on Well NK-1*
- An improved positioning model of deep-seafloor datum point at large incidence angle*
- Microplastics in sediment of the Three Gorges Reservoir:abundance and characteristics under different environmental conditions*